斑块生境下大别山五针松幼苗的光合生理特征
2017-10-16吴甘霖羊礼敏段仁燕项小燕王志高
许 远, 吴甘霖, 羊礼敏, 段仁燕, 项小燕, 王志高
(安庆师范大学 生命科学学院, 安庆 246133)
斑块生境下大别山五针松幼苗的光合生理特征
许 远, 吴甘霖, 羊礼敏, 段仁燕, 项小燕, 王志高
(安庆师范大学 生命科学学院, 安庆 246133)
根据森林循环理论,森林群落是处于不同发展阶段的镶嵌系统。测定了4个不同斑块类型(林窗斑块、建立斑块、成熟斑块和衰退斑块)下优势树种大别山五针松幼苗的光合生理特征。结果表明:随着森林循环,大别山五针松幼苗的净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、羧化效率(CE)、PSⅡ最大光化学效率(Fv/Fm)均呈现出先下降后上升的趋势。光化学猝灭系数(qP)、非光化学猝灭系数(qN)、PSⅡ量子产额(ΦPSⅡ)和电子传递速率(ETR) 的日变化在林窗斑块中较大,而在成熟斑块较小。说明林窗斑块虽然环境因子的日变化较大,但较好的光照条件有利于大别山五针松幼苗的生长,而在成熟斑块阶段,虽然环境因子的日变化较温和,但较差的光照环境,抑制了大别山五针松幼苗的生长。
光合能力;叶绿素荧光;日变化;斑块
AbstractAccording to the forest cycle theory, forest communities are the embedded systems with different development patch types. The heterogeneity of the photosynthetic characteristics ofPinusdabeshanensisseedling in different patch types (gap patch, building patch, mature patch and degenerate patch) were studied. Our results were as follows: compared to other patch phases, in gap patch phase, the seedlings ofP.dabeshanensisexhibited the highest net photosynthetic rate (Pn), stomatal conductance (Gs), transpiration rate (Tr), carboxylation efficiency (CE), and the maximum quantum efficiency of PSⅡ (Fv/Fm). The diurnal variations of coefficient of photochemical quenching (qP), coefficient of non-photochemical quenching (qN), PSⅡ quantum yields (ΦPSⅡ) and electron transport were (ETR) the highest in the gap patch, and the lowest in the mature patch. Our results suggest that the gap patch is the optimal environment for their growth, and the mature patch inhabits the growth ofP.dabeshanensisseedling.
Keywordsphotosynthetic capability; chlorophyll fluorescence; diurnal variation; patch
自然干扰和斑块动态是理解群落的结构与动态的关键一步[1]。斑块动态过程的研究是近年来生态学研究比较重要的内容[1]。Whitmore[2]在研究热带雨林时曾提出了森林循环理论。该理论指出,森林中存在着有干扰所驱动的循环。根据斑块内个体高度、胸径大小及是否存在枯倒木可人为地分为不同斑块阶段[2-3]。林窗、森林内的开放空间,驱动着森林循环。斑块的动态及发育可能改变着光环境。已有研究证实,在林窗阶段光照相对较好,而在成熟阶段有相对较弱的光照[4]。
植物叶片对不同光照的生理生态学反应是生态学研究的热点[5]。植物可通过内外形态和生理的变化来响应异质光环境[6-8]。随着光强度的增加,植物可能降低单位叶面积重量,增加叶绿素含量,降低叶绿素a/b的值,来提高其对弱光的捕获能力[9]。在异质的光环境下,不同植物的光反应是不同的。比如,一些研究证实,在同样的光环境下,常绿植物要比落叶植物具有更低的光合能力[10-11]。目前多数研究集中在不同光环境下,植物对异质光环境的反应[7,10-11],很少考虑植物对森林循环动态中不同斑块异质光环境的响应。
大别山五针松(Pinusdabeshanensis)是我国特有的极小种群,是国家Ⅱ级濒危保护植物,野外自然株数不到1000株,野外自然分布点在5~9个之间,具有重要的科学研究价值。因此,有必要深入探讨森林循环动态中不同斑块下大别山五针松幼苗光合生理特性的差异。
1 研究区概况
研究样地位于岳西县(115°24′~116°21′E,30°44′~30°51′N),海拔为1000 m左右,年均温为14.1℃~15.6℃,年降水量在1350~1400 mm左右,土壤pH为4.8~5.5。乔木层的主要优势树种为大别山五针松,伴生树种为黄山松(Pinustaiwanensis)、短柄枹栎(Quercusserrata)等,灌木层的主要优势物种为伞八仙(Hydrangeaumbellata)、山橿(Linderareflexa)、菝葜(Smilazchina)等。
2 研究方法
2.1 野外调查
调查大别山五针松林内物种的树高、胸径、冠幅,测定每个斑块的郁闭度。从自然林不同斑块内分别选取大别山五针松幼苗测定光合生理特征。人为地将郁闭度分为3个等级,0%~30%, 40%~60%和60%~100%。
2.2 斑块类型的划分
根据斑块内树木的平均高度、胸径大小及是否存在枯倒木等人为地将斑块划分为林窗斑块、建立斑块、成熟阶段和衰退阶段[2-3]。林窗斑块:内有枯倒木,斑块的树木平均胸径小于样地的平均胸径,郁闭度在0%~30%。建立斑块:斑块的树木平均胸径约等于样地的平均胸径,郁闭度在40%~30%。成熟阶段:斑块内的树木平均胸径高于样地的平均胸径,郁闭度在60%~100%。衰退阶段:斑块内的树木平均胸径高于样地的平均胸径,其中有个别树较大,其高度超过林冠层的平均高度,郁闭度小于60%。
2.3 光合特征的测定
选择植物的叶片,在中上部测定叶片的光合特征,测定时间在晴朗的上午8:00到11:00之间。为降低叶片的影响,仅选择当年生的新生叶来测定。通过便携式光合作用仪(Li-6400) 的针叶叶室测定光响应曲线和CO2响应曲线,设定的光合有效辐射通量密度(PPFD)为10个(0~1000 μmol·m-2·s-1),采用排水体积法测定叶面积[12]。测定叶片的净光合速率(Pn,μmol·m-2·s-1)、胞间 CO2浓度( Ci,mol·mol-1) 、蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,μmol·m-2·s-1)等光合参数,每个重复测定4次。并计算下面几个指标,水分利用率(WUE,mmol/mol)=Pn/Tr、羧化效率(CE,μmol·m-2·s-1)=Pn/Ci。
2.4 叶绿素荧光变量的测定
用叶绿素荧光仪(PAM-2100)测定大别山五针松的叶绿素荧光特征。样品与气体交换的测定为同一叶片。用作用光测定早晨植物未活化时Fo和Fm值,最大光能转化效率Fv/Fm=(Fm-Fo) /Fm。从6:00到18:00测定光化学猝灭系数(qP)、非光化学猝灭系数(qN)、PSⅡ量子产额(ΦPSⅡ)、电子传递速率(ETR)的日变化,每个重复测定4次。
2.5 数据分析
各变量均以加减一个标准误表示,差异性水平(P<0.05)分析均采用SPSS13.0 (SPSS Inc., USA)。
3 结果
光合作用是大别山五针松生长发育的重要基础,它为大别山五针松的生长发育提供必备的能量和光合产物。一般而言,胁迫会抑制植物的光合作用,甚至破坏植物的叶绿体结构和功能。净光合速率表示的是植物单位叶面积的光合同化能力,可直接反应植物受到胁迫的程度。在林窗阶段,大别山五针松幼苗的光合能力最大;随着森林循环(G-B-M-D),其值呈现出先下降后上升的趋势,差异明显。其中,林窗斑块内大别山五针松幼苗的净光合速率约为成熟斑块的2倍(图1)。气孔导度、蒸腾速率和羧化效率也呈现出先下降后上升的趋势,仅水分利用效率呈现先增加后下降的趋势。不同斑块间仅胞间CO2浓度相差较小,其他特征(净光合速率、气孔导度、蒸腾速率、羧化效率和水分利用效率)均存在较大的差异(图1)。
PSⅡ最大光化学效率(Fv/Fm)也是测定植物受到光抑制状况的关键指标。随着胁迫的增加,光系统Ⅱ会受到一定程度的损伤,导致PSⅡ最大光化学效率下降。在不同的斑块阶段,PSⅡ最大光化学效率(Fv/Fm) 存在明显的差异(P<0.05),其中林窗阶段的PSⅡ最大光化学效率最大,成熟阶段最低(图2)。
叶绿素荧光参数qP、qN、ΦPSⅡ、ETR均随着时间的推移而发生变化。一天中,从6:00到18:00,qP和ΦPSⅡ呈现出先下降后上升的趋势,而qN和ETR呈现出相反的趋势(图3)。整体而言,一天中林窗阶段的叶绿素荧光参数变化最大,而成熟阶段变化相对较小。



不同斑块 不同斑块
注:横坐标的G、B、M和D分别表示林窗斑块、建立斑块、成熟斑块和衰退斑块;不同字母表示差异显著(P<0.05)
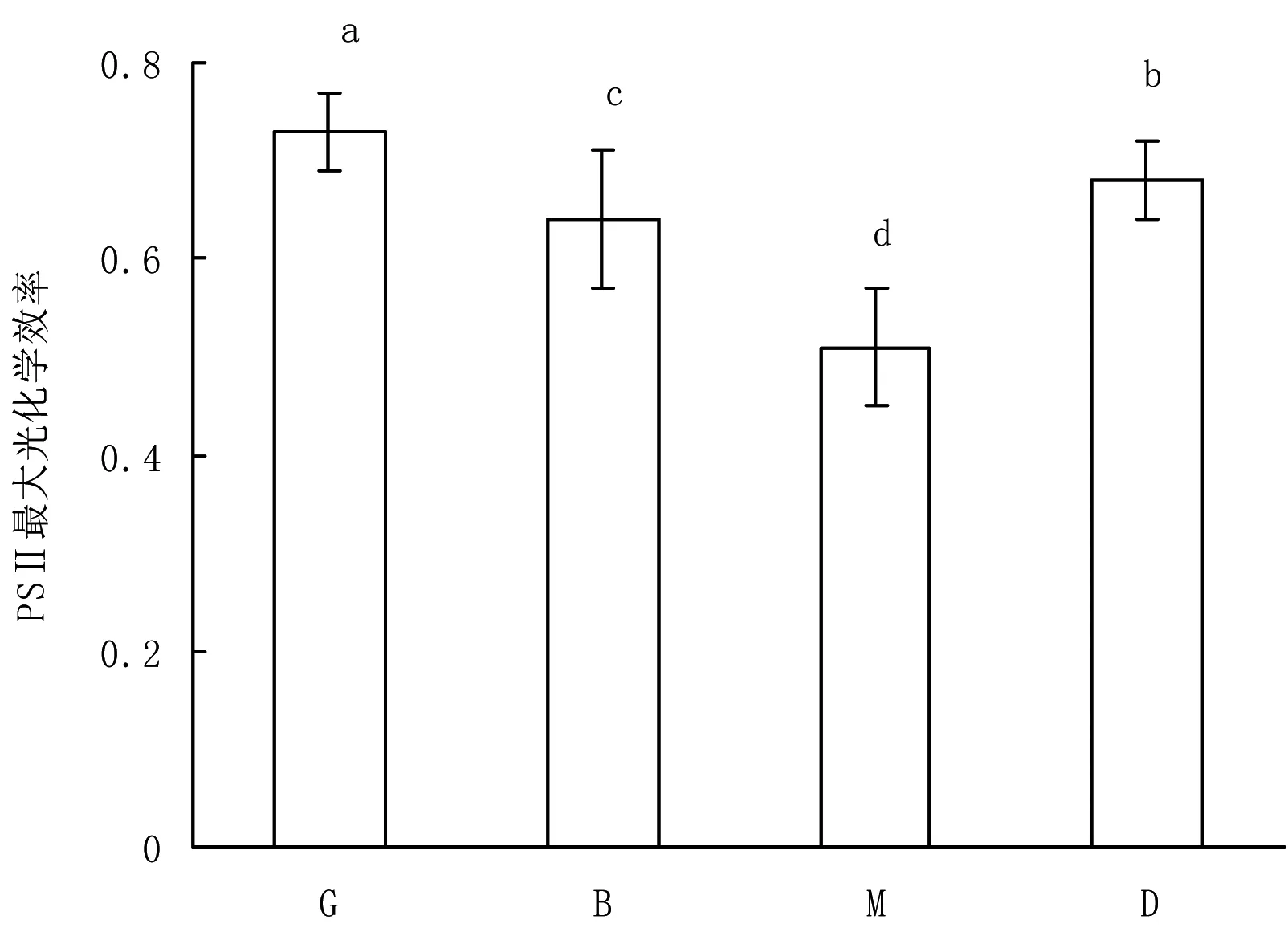
图2 不同斑块内大别山五针松的PSⅡ最大光化学效率
注:横坐标的G、B、M和D分别表示林窗斑块、建立斑块、成熟斑块和衰退斑块;不同字母表示差异显著(P<0.05)

图3 不同斑块大别山五针松叶绿素荧光参数(qP,qN,ΦPSⅡ,ETR)的日变化
4 讨论
随着森林循环,不同斑块有不同的郁闭度,造成不同斑块内光环境的异质性。叶片是植物进行光合作用的主要器官,可以通过一系列的生理生态特征的变化来响应外界光环境的变化[13]。
林窗是森林生态系统重要的驱动力,对维持植物的更新起着重要作用[14-15]。在林窗斑块有较强的光照,为了阻止光的伤害和水分损失,植物会投入较少的N到光合器官[4,14-16]。林窗斑块内大别山五针松气体交换参数(净光合速率、气孔导度、蒸腾速率、羧化效率)要高于其他3个斑块阶段。光系统Ⅱ的最大光化学效率(Fv/Fm)可高效地反映植物是否受到胁迫。在条件最好的情况下,如果没有受到光抑制,其值约为0.8[17]。在林窗斑块,大别山五针松的Fv/Fm约为0.8,qN也相对较低(小于0.6),这意味着林窗斑块有利于其生长与更新。在其他斑块,尤其是成熟斑块,较弱的光照造成净光合速率、气孔导度、蒸腾速率、羧化效率明显的下降,表明大别山五针松幼苗的光合生理特征受到明显的受到抑制。这意味大别山五针松幼苗虽然可以在成熟斑块低光下存活,但叶片将更多的光能用于维持存活。这个结果与我们野外观察到一些成熟斑块内存在着死亡的个体一致。整体看来,4个不同斑块类型(林窗斑块、建立斑块、成熟斑块和衰退斑块)下优势树种大别山五针松幼苗的光合生理特征呈现先下降后上升的趋势,与Duan等[4]所研究的常绿Cyclobalanopsisglauca及落叶树种Bothrocaryumcontroversum对森林循环动态中不同斑块的反应类似。此外,森林循环动态中,不同斑块的温度及竞争状况也会发生变化。有研究证实,物种对不同群落(斑块)内光照、种内种间竞争及温度变化的生理生态学响应也存在着差异[18-20]。
森林循环动态中,林窗斑块虽然环境因子的日变化较大,但较好的光照条件有利于大别山五针松幼苗的生长,而在成熟斑块阶段,虽然环境因子的日变化较温和,但较差的光照环境和较强的竞争,抑制了大别山五针松幼苗的生长。因此,大别山五针松幼苗要完成生长与更新,要克服不同斑块的微生境条件,特别是成熟斑块较低的光照是其更新的重要限制因子。
[1]MANABE T, SHIMATANI K, KAWARASAKI S, et al. The patch mosaic of an old-growth warm-temperate forest: patch-level descriptions of 40-year gap-forming processes and community structures[J]. Ecol Res, 2009, 24(3):575-586.
[2]WHITMORE T C. Canopy gaps and the two major groups of forest trees[J]. Ecology, 1989, 70(3):536-538.
[3]DUAN R Y, HUANG M Y, Wang X A. Forest patch dynamics initiated by disturbance: distribution pattern ofLarixChinensis(bessen) patches along the altitudinal gradient in relation to light and temperature conditions (Qinling Mts. China)[J]. Pol J Ecol, 2013, 61(2):271-281.
[4]DUAN R, HUANG M, KONG X, et al. Ecophysiological responses to different forest patch type of two codominant tree seedlings[J]. Ecol Evol, 2014, 5(2):265-274.
[5]WALTERS M B, REICH P B. Trade-offs in low-light CO2, exchange: a component of variation in shade tolerance among cold temperate tree seedlings[J]. Funct Ecol, 2000, 14(2):155-165.
[6]MILLER R E, GLEADOW R M, WOODROW I E. Cyanogenesis in tropicalPrunusturneriana: characterisation, variation and response to low light[J]. Funct Plant Biol, 2004, 31(5):491-503.
[7]OSONE Y, TATENO M. Nitrogen absorption by roots as a cause of interspecific variations in leaf nitrogen concentration and photosynthetic capacity[J]. Funct Ecol, 2005, 19(3):460-470.
[8]李志辉, 卢兆银, 杨 艳,等. 松栎混交林中马尾松与白栎光合作用的日变化[J]. 中南林业科技大学学报, 2007, 27(1):76-80.
[10]SHIPLEY B, LECHOWICZ M J, WRIGHT I, et al. Fundamental trade-offs generating the worldwide leaf economics spectrum[J]. Ecology, 2006, 87(3):535-541.
[11]PRINGLE E G, ADAMS R I, BROADBENT E, et al. Distinct leaf-trait syndromes of evergreen and deciduous trees in a seasonally dry tropical forest[J]. Biotropica, 2011, 43(3):299-308.
[12]朱教君, 康宏樟, 李智辉. 不同水分胁迫方式对沙地樟子松幼苗光合特性的影响[J]. 北京林业大学学报, 2006, 28(2):57-63.
[13]POORTER L, BONGERS F. Leaf traits are good predictors of plant performance across 53 rain forest species[J]. Ecology, 2006, 87(7):1733-1743.
[14]KITAOKA S, WATANABE M, WATANABE Y, et al. Growth of regenerated tree seedlings associated with microclimatic change in a mature larch plantation after harvesting[J]. Landscape Ecol Eng, 2009, 5(2):137-145.
[15]KUPTZ D, GRAMS T E E, GÜNTER S. Light acclimation of four native tree species in felling gaps within a tropical mountain rainforest[J]. Trees, 2010, 24(1):117-127.
[16]OGUCHI R, HIKOSAKA K, HIURA T, et al. Costs and benefits of photosynthetic light acclimation by tree seedlings in response to gap formation[J]. Oecologia, 2008, 155(4):665-675.
[17]HOUTER N C, PONS T L. Gap size effects on photoinhibition in understorey saplings in tropical rainforest[J]. Plant Ecol, 2005, 179(1):43-51.
[18]DUAN B, DONG T, ZHANG X, et al. Ecophysiological responses of two dominant subalpine tree speciesBetulaalbo-sinensis, andAbiesfaxoniana, to intra-and interspecific competition under elevated temperature[J]. Forest Ecology & Management, 2014, 323:20-27.
[20]ZANOLLA M, ALTAMIRANO M, CARMONA R, et al. Photosynthetic plasticity of the genus Asparagopsis, (Bonnemaisoniales, Rhodophyta) in response to temperature: implications for invasiveness[J]. Biological Invasions, 2015, 17(5):1341-1353.
The photosynthetic physiological characteristics ofPinusdabeshanensisinpatchhabitats
XUYuan,WUGan-lin,YANGLi-min,DUANRen-yan,XIANGXiao-yan,WANGZhi-gao
(College of Life Sciences, Anqing Normal University, Anqing 246133, China)
Q945
A
2095-1736(2017)05-0070-04
2016-05-08;
2016-06-10
国家自然科学基金(31570417);林业公益性行业科研专项(201304314);安徽省自然科学基金(1608085MC63)
许 远,硕士,讲师,研究方向为植物学,E-mail:358828631@qq.com
段仁燕,博士,教授,主要从事生态学研究,E-mail:duanrenyan78@163.com
doi∶10.3969/j.issn.2095-1736.2017.05.070
