不同养殖条件下滩头雅罗鱼性腺发育的组织学研究及生长比较
2017-10-16苏宝锋常玉梅张立民梁利群
闫 浩, 苏宝锋, 常玉梅, 孙 博, 尚 梅, 张立民, 梁利群
(1. 中国水产科学研究院 黑龙江水产研究所, 哈尔滨 150070;2. 上海海洋大学 水产与生命学院, 上海 201306)
不同养殖条件下滩头雅罗鱼性腺发育的组织学研究及生长比较
闫 浩1,2, 苏宝锋1, 常玉梅1, 孙 博1, 尚 梅1, 张立民1, 梁利群1
(1. 中国水产科学研究院 黑龙江水产研究所, 哈尔滨 150070;2. 上海海洋大学 水产与生命学院, 上海 201306)
采用组织切片技术对滩头雅罗鱼(Tribolodonbrandti)性腺结构、弱碱和淡水性腺发育、产卵时间及类型进行了研究。滩头雅罗鱼精巢为典型的小叶型结构,共可分为6个时期。按照卵巢外形特征和生殖细胞类型,卵巢可分为6个发育时期,对应卵细胞6个时相。研究表明,性成熟的滩头雅罗鱼在9月末性腺发育到第Ⅲ期,并以Ⅲ期越冬。而未达到性成熟的幼鱼(2+龄)性腺发育到Ⅱ期,并以Ⅱ期越冬。性成熟的雄鱼在4月末或5月初性腺发育到第Ⅴ期,雌鱼在5月中下旬或6月初才能发育到第Ⅴ期,雄鱼性腺发育快于雌鱼。2~3龄个体在淡水或弱碱水环境中养殖,性腺发育时相并无明显区别。弱碱水2+龄群体体重显著高于淡水同龄群体,然而3+龄群体在弱碱水和淡水中体重没有显著差异。同时,弱碱水和淡水中的同龄雌、雄之间体重变化存在差异。在性腺发育周期中,雌、雄的平均肥满度在1.313~1.697之间波动。随性周期变化,雄性从II期开始性腺指数显著上升;雌性性腺指数在不同发育时期都存在显著差异,在V期最大为14.22。滩头雅罗鱼性腺发育高峰期一年只出现一次,为一次性产卵类型。
滩头雅罗鱼;生殖细胞;性腺发育;性周期;性成熟
AbstractUsing the histological methods, the gonadal development, structures of gonad, spawning time and type of reproduction inTribolodonbrandtiwere studied. The testes ofT.brandtihave typical lobular structures, and can be divided into six developmental stages. According to the characteristics of the ovaries and the types of ovarian germ cells, the oogenesis process can also be divided into six developmental stages. The microscopic observation showed that the gonadal development of sexual matureT.brandtidifferentiated to stage III at the end of September, and stayed at the phase III in the winter period. While the gonadal development of immature juveniles age 2+was at the phase Ⅱ and remained such stage during the winter period. In the late April and early May, gonadal development of sexual mature males differentiated to the phase V, but in the females it was until late May or early June that they reached to the developmental phase V. Taken together, gonadal development of the males was faster than that of the females. No significant difference was observed in the gonad development of 2-3-year-old juvenile cultured in either fresh water or weak alkali water environment. The body weight 2+-year-old group in the weak alkali water were significantly higher than that in the fresh water, but no significant difference was found between 3+-year-old groups reared in weak alkali environment and those in fresh water. Meanwhile, significant differences of body weight existed between females and males in the weak alkali as well as in the fresh water. In the gonad development cycle, the average condition factor of female and male ranged from 1.313 to 1.697. During the sexual cycle, the male gonad index began to rise significantly from the II period, while the female gonad index was significantly different among development periods, with the peak value of 14.22 in the period of V. The conclusion could be drawn that the gonadal development peak ofT.brandtiappears only once a year, the ovary is partial synchronic, and this species is iteroparous.
KeywordsTribolodonbrandti; germ cells; gonadal development; sexual cycle; sexual maturity
滩头雅罗鱼(Tribolodonbrandti)隶属于鲤科,雅罗鱼亚科,三块鱼属,是鲤科鱼类中目前已知的唯一溯河洄游进行生殖产卵的鱼类。此鱼类在我国仅分布在黑龙江绥芬河、图们江流域,长梭型,其肉质细嫩,成熟期色彩艳丽,市场价值较高,是一种稀有的经济型土著鱼类[1]。每年的5月—7月,发育成熟的滩头雅罗鱼溯河而上,选择合适的产卵场所进行产卵。但是由于滩头雅罗鱼渔汛集中,以及不合理的捕捞活动等,导致该鱼溯河种群数量连年减少。加上近年来,由于绥芬河流域疏于管理,截流建坝用于灌溉及生产等行为,引起了生态环境变化,导致滩头雅罗鱼的产卵环境发生了很大变化,致使该鱼产卵种群数量呈逐年锐减趋势。因此,研究行之有效的保护措施保存这一珍贵的鱼类品种已成当务之急。目前,关于滩头雅罗鱼的研究主要集中在野生亲鱼资源和生物学调查[2],人工放流和资源保护[1],苗种繁育[3],生殖群体结构和繁殖策略[4],胚胎发育[5],幼鱼对盐碱的适应性分析[6],种群亲缘关系[7]等方面,而对滩头雅罗鱼性腺发育的研究还尚属空白。性腺是鱼类生殖繁衍的基础,本研究主要运用组织学、统计学、形态学手段对滩头雅罗鱼的性腺发育进行观察、计算、描述,比较精巢和卵巢在不同发育时期的变化过程,探讨在弱碱水和淡水环境中性腺发育的区别,以期为保护这一珍贵鱼类资源,同时合理利用盐碱水体,扩大养殖规模提供参考资料。
1 材料与方法
本实验所用实验材料均来自人工养殖所得的滩头雅罗鱼个体,弱碱水组养殖于黑龙江水产研究所呼兰实验站(年平均碱度:6.88 mmol/L),淡水组养殖于黑龙江水产研究所循环水实验车间。本实验在2015年9月末到2016年9月进行,期间定期收集实验材料,共得样品数130尾。体重66.41~646.04 g,体长16.4~35.5 cm。
鱼体经称重、测量体长后,分别取性腺组织样品固定于Bouin′s液中,经24 h固定后取出转到70%乙醇中长期保存。样品组织经脱水、二甲苯透明后用石蜡包埋,使用切片机连续切片,切片厚度5~7 μm,经苏木精-伊红(H.E)染色,最后用中性树胶封片。用LEICA DMI 6000倒置显微镜进行观察并拍照。
性腺成熟系数和肥满度按以下公式求出:性腺指数(GSI)=性腺重/体重×100,肥满度(CF)=[体重/体长3]×100。利用SPSS 21.0软件,采用Duncan法对不同发育时期的雌、雄性腺指数进行多重比较,分析其显著性差异;采用独立样本t检验,对淡水和弱碱水水体中养殖的2+、3和3+龄滩头雅罗鱼体重和性腺重进行显著性分析。
2 结果与分析
2.1 性腺的形态特征及分期划分
2.1.1 精巢 参照鲥鱼精细胞不同发育阶段的形态特征划分标准[8],滩头雅罗鱼精细胞共分为5种,即精原细胞、初级精母细胞、次级精母细胞、精子细胞和精子。根据精巢形态特征和精巢内5种细胞的形态变化,可将滩头雅罗鱼的精巢分为6个时期。
Ⅰ期:Ⅰ期精巢和卵巢形态相似,呈透明细线状,用肉眼不能辨别出雌雄,有少量的精原细胞,排列松散(图1-1)。
Ⅱ期:精巢表面有少量血管分布,呈细带状,肉眼可分出雌雄,精巢内生殖细胞的类型没有太大改变,Ⅰ期的精原细胞经不断的有丝分裂与增殖形成很多排列紧密的精原细胞(图1-2),此期为精原细胞增殖期。
Ⅲ期:Ⅲ期精巢呈带状,灰白色,表面光滑,黏连着一层黑色腹膜,切片中可见多种类型的生殖细胞,主要有初级精母细胞、次级精母细胞、精子细胞、间质细胞及少量的精原细胞(图1-3、4)。
Ⅳ期:Ⅳ期精巢呈柱状,表面有皱褶,用力按压腹部,有白色精液流出。切片中可见部分精小囊开始破裂,成熟精子被排到小叶腔内(图1-5)。
Ⅴ期:精巢呈宽扁状,乳白色,紧贴于鱼鳔两侧,看不到血管分布,提起鱼头,轻轻挤压腹部,有精液流出。此期精子的嗜碱性最强,被染成深蓝色,精小囊相互融合,精小叶界限不明显,精小叶内充满大量成熟的精子(图1-6)。
Ⅵ期:此期精巢排精后体积萎缩,局部有充血现象,呈粉红色。
2.1.2 卵巢 根据滩头雅罗鱼不同发育时期卵巢形态变化和生殖细胞的形态特征,可将卵巢发育分为6个时期,分别相对应6个时相的生殖细胞。
Ⅰ期,卵巢呈透明的细线状,紧贴于背部两侧,肉眼不能分辨雌雄。此期卵巢中的生殖细胞为第1时相卵原细胞,紧贴在卵巢生殖上皮边缘,有各种不同的形态类型,细胞核明显、较大,占整个卵子直径的一半以上,胞核呈强嗜碱性,经HE染色呈蓝紫色(图2-7)。
Ⅱ期,卵巢呈棒状,表面可见少量血管,颜色微红。卵粒尚未形成,但肉眼可分辨出雌雄。Ⅱ期卵巢中主要以第2时相卵母细胞为主,约占视野中卵子总数的67.3%~76.4%,另外还有小部分的第1时相卵母细胞。由于受到挤压,第2时相卵母细胞形态多变,形状不规则,多呈多边形,也有少数圆形或椭圆形。卵径为56.864~126.044 μm,核径为21.751~60.628 μm,核仁4~16个,在细胞核内缘呈环形分布。胞质呈强的嗜碱性,被染成蓝紫色;胞核呈弱的嗜酸性,呈淡红色。卵膜周围隐约可见一层滤泡膜,随着卵母细胞的不断发育,单层滤泡膜逐渐明显、加厚(图2-8)。
Ⅲ期,卵巢体积明显增大,血管分布明显增多。透过卵巢隔膜可见细小的卵粒,排列紧密呈淡黄色或白色。此时卵巢中可见第2、3时相卵母细胞,但以第3时相为主,Ⅲ期末可占视野中卵子总数的46.5%~67.4%,多呈圆形,卵径141.525~322.426 μm,核径为34.330~76.539 μm,此时相最明显的变化是胞质卵膜边缘开始出现一层环状皮质液泡,4~48个大小不等。随着卵母细胞的发育,皮质液泡逐渐向内延伸,层数不断增多,在3时相后期可达5~7层,几乎充满整个细胞质。在胞核周围开始出现少量的卵黄颗粒,随着卵母细胞的不断发育,逐渐向外延伸开来。卵膜外的滤泡膜由单层逐渐发展为双层,另外在滤泡膜的内缘开始出现一层薄的放射膜(图2-9、10)。
Ⅳ期,卵巢体积进一步增大,呈囊状,里面充满卵粒,几乎充满整个腹腔,呈黄色,个别卵粒出现灰绿色。此期卵巢中可见2、3和4时相卵母细胞,其中以第4时相居多,占切片卵细胞总数的45.7%~78.6%,呈圆型或椭圆形,卵径为414.530~1045.731 μm,核径为82.324~112.431 μm,放射膜逐渐增厚,放射纹清晰可见,可达3.638~5.571 μm。胞质中卵黄颗粒增多,皮质液泡被挤压到卵膜内缘,为1~2层,有时可见受精孔(图2-11)。在卵母细胞第4时相中后期,胞核开始发生极化,逐渐向动物极移动。在生产中第Ⅳ期的卵通常经过人工催产产生成熟的生殖细胞,此期的卵也通常用来生产鱼子酱。
Ⅴ期,卵巢体积增至最大,充满整个腹腔。卵粒呈金黄或灰白色。此期的卵已完全成熟,鱼腹部松软,轻轻按压有卵粒流出,排出的卵可吸水膨胀,卵径达485.828~1134.002 μm(图2-12)。切片中可见各个时相卵母细胞,以第5时相为主,约占总数的44.2%~56.8%,其余多为第1、2时相卵母细胞,3时相卵母细胞偶尔可见。第5时相卵母细胞核已移至动物极,核仁消失,核膜溶解,此期滤泡膜呈游离状,波浪状。卵子可从滤泡膜中解脱出来,排到卵巢腔内,5时相卵母细胞内卵黄颗粒排列松散,相互融合呈板块状。
Ⅵ期,即卵子排空期,经人工催产,大部分成熟卵排出体外,卵巢中剩余一些空的滤泡膜及各个时相卵母细胞,并逐渐恢复到第Ⅱ期。

图1 雄性滩头雅罗鱼性腺不同发育时期组织学观察
1:Ⅰ期精巢,见核仁明显的精原细胞;2:Ⅱ期精巢,精巢中隐约可见精小叶,精原细胞数目增多;3:Ⅲ期精巢,精小叶明显,中间为小叶腔,可见多种类型的生殖细胞;4:精小叶,此时在精小叶内可见多个精小囊;5:Ⅳ期精巢,精小叶内开始出现精子;6:Ⅴ期精巢,精小叶相互融合,内充满精子
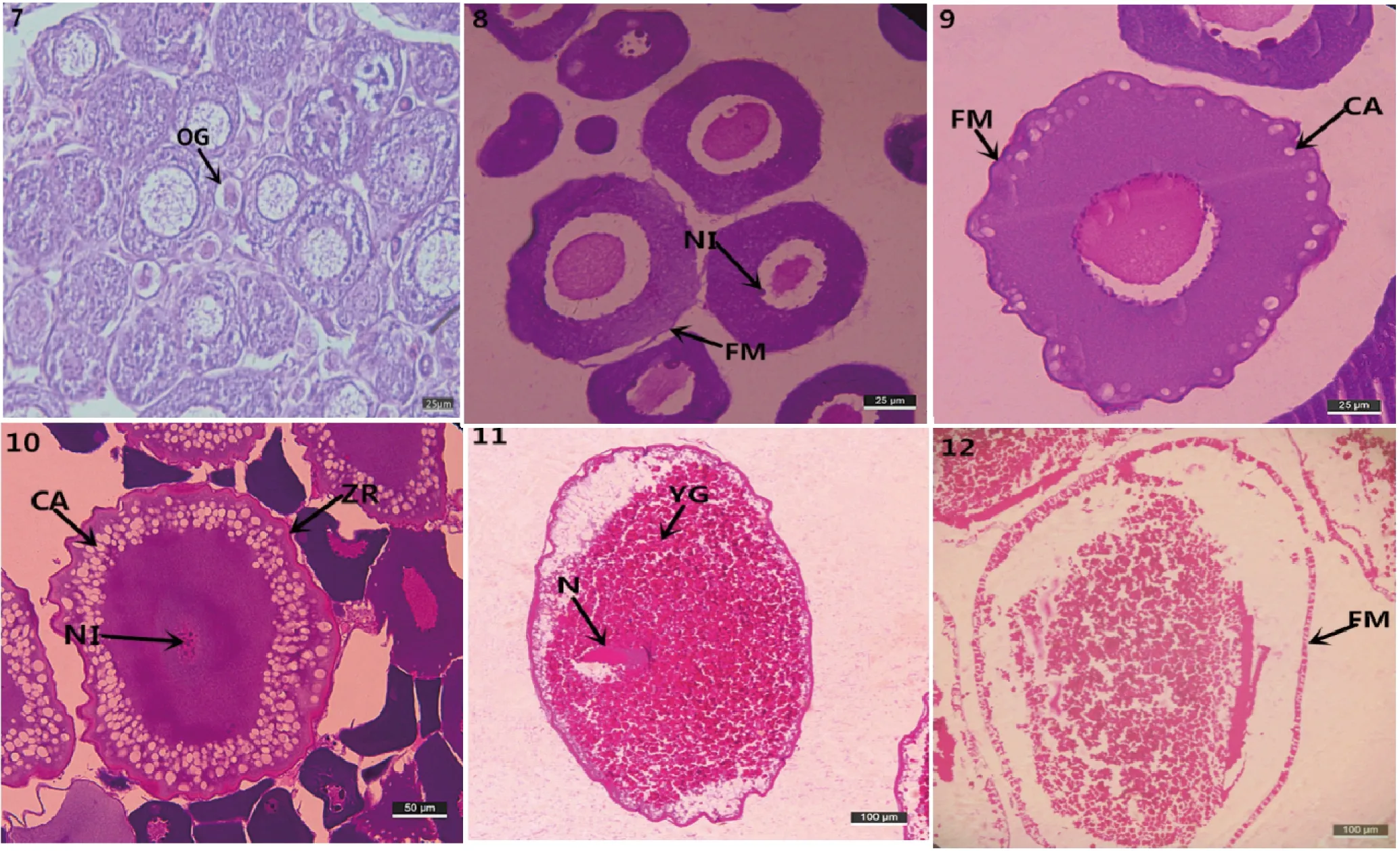
图2 雌性滩头雅罗鱼性腺不同发育时期组织学观察
7:Ⅰ期卵巢,此期胞核较大,占整个卵子直径的一半以上;8:Ⅱ期卵巢,在细胞核内可见多个核仁,呈环状排列;9:Ⅲ期卵巢,开始在滤泡膜的内缘出现几个或几十个大小不等的液泡,形成双层滤泡膜;10:Ⅲ期卵巢后期,皮质液泡层层数增多,向胞核延伸,辐射带增厚;11:Ⅳ期卵巢,胞质中卵黄颗粒增多,胞核发生偏位,逐步向动物极靠近;12:Ⅴ期卵巢,卵子游离于滤泡膜,卵黄颗粒排列松散
2.2 滩头雅罗鱼性腺发育过程中肥满度CF和性腺指数GSI变化
滩头雅罗鱼在整个性腺发育周期中,雌、雄鱼的平均肥满度在1.313~1.697之间上下波动,在不同发育时期间变化不大(P>0.05),见图3。精巢在不同发育时期,性腺指数(GSI)变化如图3所示。随着精巢的不断发育,GSI不断升高,从精巢发育Ⅲ期开始,GSI升高明显加快,在Ⅴ期达到最大值4.40左右,显著高于其他各期(P<0.05)。精巢GSI在Ⅱ、Ⅲ期间无显著差异,增长缓慢,而在第Ⅳ期迅速升高,显著高于Ⅱ、Ⅲ期(P<0.05)。在卵巢中,随着卵巢的不断发育,营养物质和卵黄的不断积累,卵巢增重加快,在第Ⅴ期GSI达到最大值14.22左右,显著高于其他各个时期。卵巢在Ⅱ~Ⅴ期,增重明显,GSI在Ⅱ、Ⅲ、Ⅳ及Ⅴ期之间都有显著差异(P<0.05),见图3。
2.3 在淡水和弱碱水环境中2+、3和3+龄滩头雅罗鱼体重、性腺重和性腺发育时期比较
经过t检验,淡水2+龄群体(79.62±16.30)g和弱碱水2+龄群体(106.80±34.30)g在体重上存在显著差异(P<0.05,表1中未列出)。其他群体组间(3龄弱碱水、淡水间及3+龄弱碱水、淡水间)在体重比较上没有显著差异。对于2+龄群体组,淡水中的滩头雅罗鱼雄性体重和性腺重量显著高于雌性;弱碱水中的雌、雄在体重和性腺重量上没有显著差异。弱碱水中雌性体重显著高于淡水雌性。淡水中雄性性腺重显著高于弱碱水雄性(P<0.05)。对于3龄群体组,弱碱水雌雄体重没有显著差异,但是,淡水雌性体重和性腺重,以及弱碱水雌性性腺重显著高于对应的雄性;弱碱水中雄性体重显著高于淡水中雄性(P<0.05)。
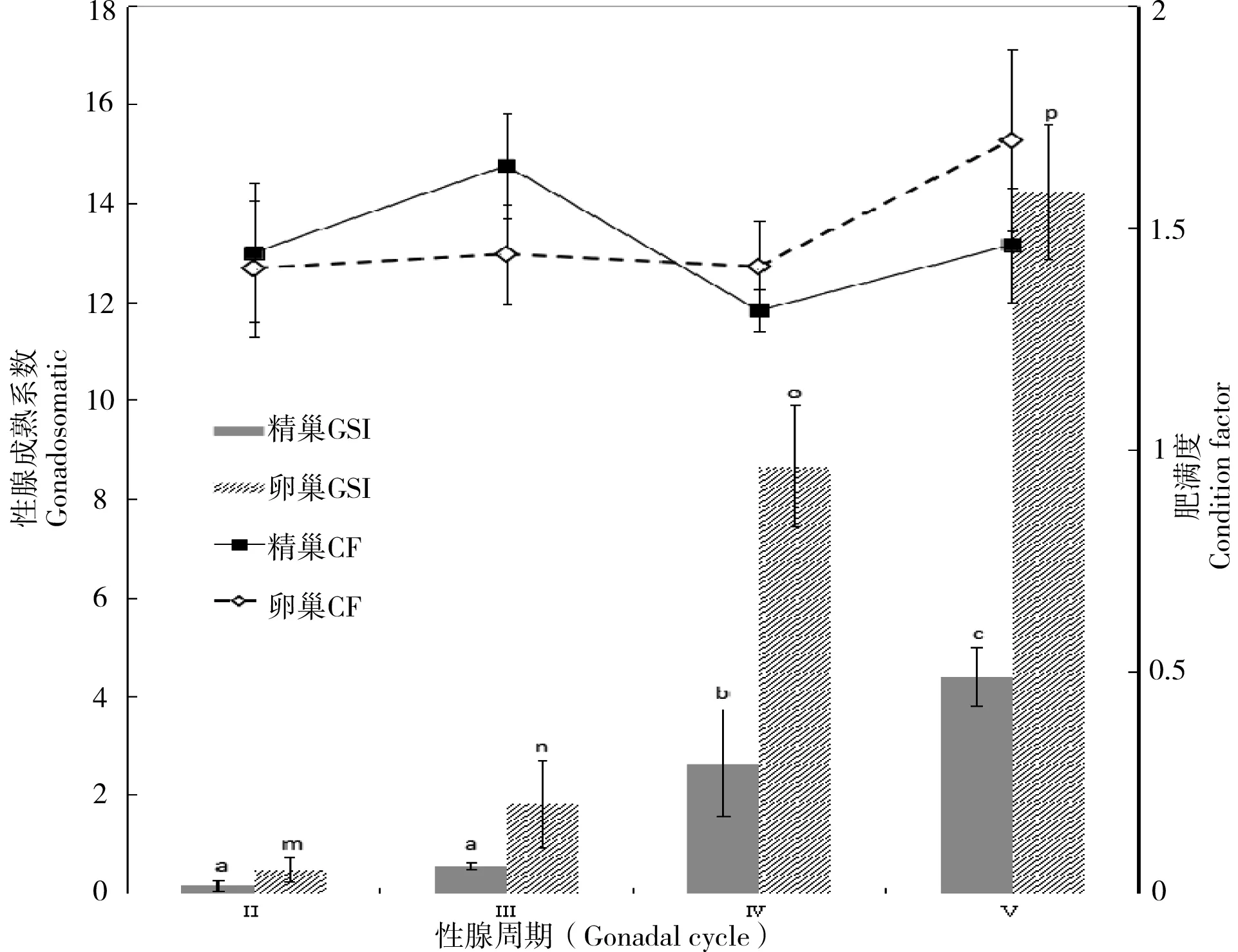
图 3 滩头雅罗鱼性腺发育和肥满度变化
a、b、c:不同发育时期,精巢性腺指数显著性分析;不同字母代表差异显著(P<0.05),相同字母表示差异不显著(P>0.05)。m、n、o、p:不同发育时期,卵巢性腺指数显著性分析;不同字母代表差异显著(P<0.05),相同字母表示差异不显著(P>0.05)
对于3+龄群体组,经过t检验,淡水群体(119.78±20.23)g和弱碱水群体(128.40±12.37)g在体重上没有差异(P>0.05,表1中未列出)。弱碱水雌、雄体重没有显著差异,但是,淡水雌性的体重和性腺重,以及弱碱水雌性性腺重显著高于对应的雄性;弱碱水中雄性体重显著高于淡水中雄性(P<0.05)。
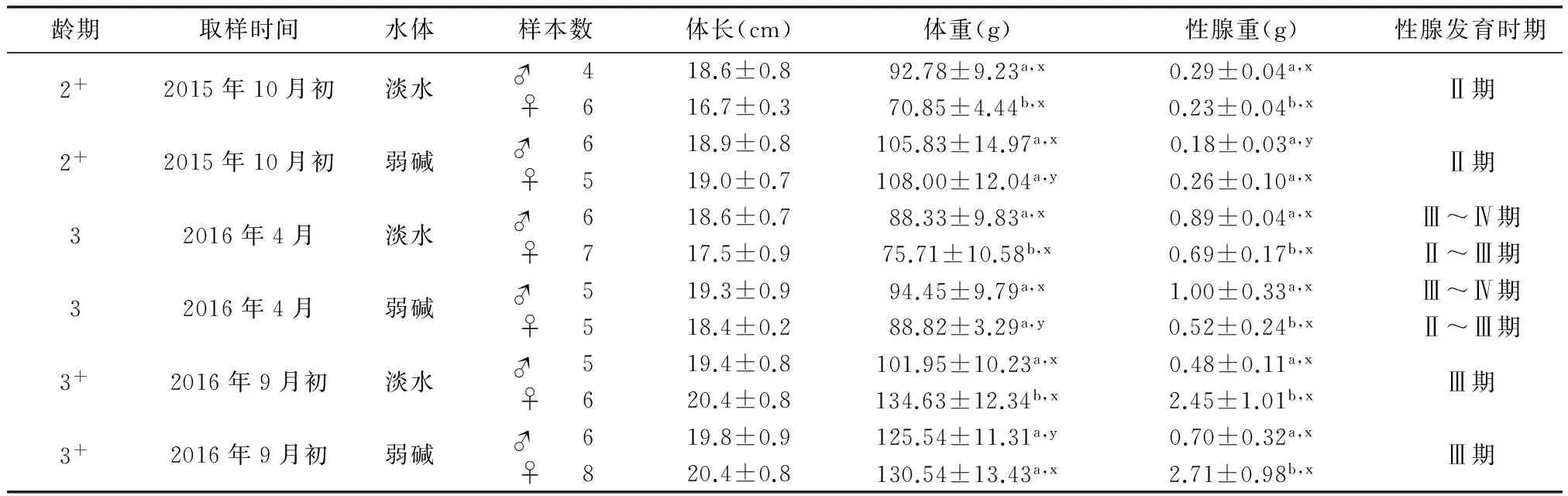
表1 2+、3和3+龄滩头雅罗鱼不同养殖水体生物学数据比较 (样本n=69)
a、b表示相同养殖水体内同龄雅罗鱼雌、雄比较,相同字母表示没有显著差异(P>0.05),不同字母表示显著差异(P<0.05);x、y表示不同养殖水体同龄滩头雅罗鱼雌、雄分别比较。相同字母表示没有显著差异(P>0.05),不同字母表示显著差异(P<0.05)
在淡水或弱碱水环境中,2+龄群体和3+龄群体雌雄性腺发育时期并无差别。在10月初2+龄群体性腺均发育到Ⅱ期;第二年4月,2+龄群体转变为3龄,雄性性腺发育到Ⅲ~Ⅳ期,雌性发育到Ⅱ~Ⅲ期,雄性性腺发育略快于雌性。至9月初,无论雌雄,性腺均又发育到Ⅲ期。2+龄群体经越冬期(10月至翌年3月)体重略有下降,但性腺重表现为增长趋势。
如表2所示,在弱碱水体中,性成熟的滩头雅罗鱼(4+~7+龄)在10月初性腺发育到第Ⅲ期。第二年,雄鱼在4月末或5月初性腺发育到第Ⅴ期,雌鱼在5月中下旬或6月初才能发育到第Ⅴ期,雄鱼性腺发育快于雌鱼。
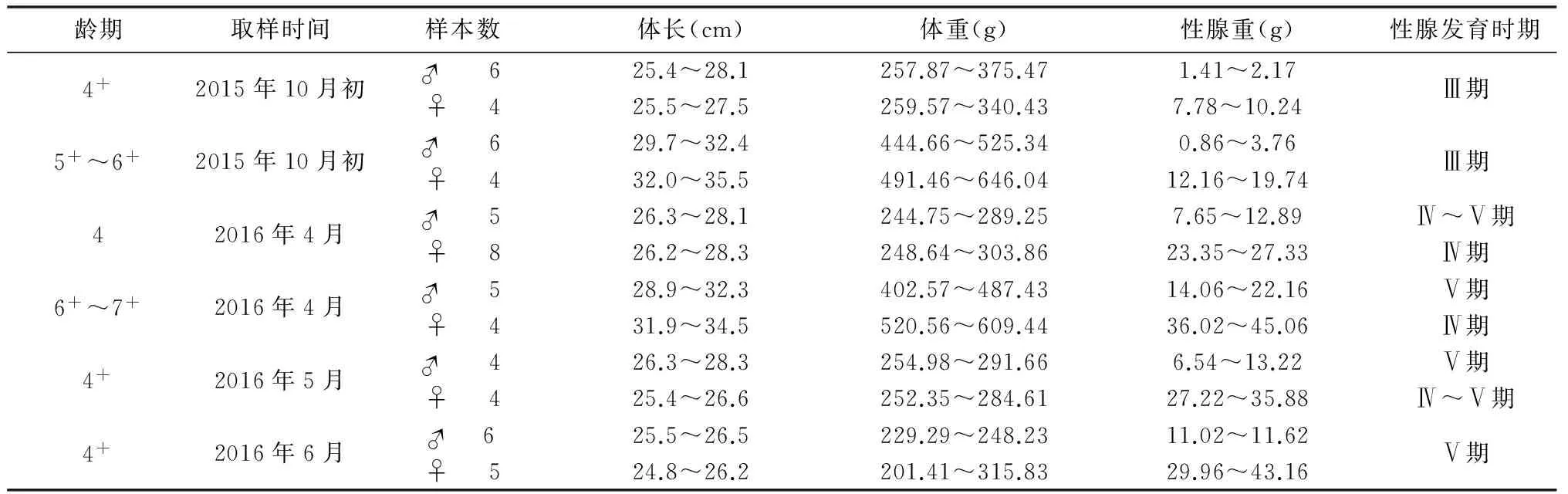
表 2 4+ ~7+ 龄滩头雅罗鱼在弱碱水体中生物学数据(样本n=61)
3 讨论
3.1 东北养殖滩头雅罗鱼精巢结构及发育特点
鱼类精巢按照精巢内生殖细胞的排列方式及分布特点一般可分为两种:小叶型精巢和小管型精巢[9]。小叶型精巢由许多精小叶组成,各个精小叶之间被纤维结缔组织薄膜隔开。在精小叶内,初级精原细胞在有丝分裂过程中形成精小囊,内含精原细胞。一个精小叶中含有多个精小囊,同一个精小囊内的生精细胞处在同一发育时期。经过精子发生和精子形成后,精小囊内充满成熟精子,在生殖季节内精小囊破裂,成熟精子释放到小叶腔内,最后由输精管排出体外。大多数鱼类都属于这种类型,如唐鱼(Tanichthysalbonubes)[10]、刀鲚(Coilianasus)[11]、黄颡鱼(Pelteobagrusfulvidraco)[12]等。小管型精巢有许多定向排列的小管组成,每一个小管都通向中央腔。在小管盲端存在精原细胞,并逐渐形成小囊结构,随着精巢内生殖细胞不断发育,小囊逐渐向中央腔靠拢,当精子成熟后小囊破裂,成熟精子释放到中央腔内,中央腔与输精管相通,释放到中央腔的精子被排出体外。只有少数的鱼属于这种类型,如孔雀鱼(Poeciliareticulata)[13]。本研究切片观察发现,滩头雅罗鱼精巢应归属于小叶型精巢。性成熟的滩头雅罗鱼雄鱼(3龄及以上龄期)在4月下旬到5月初精巢已发育到第Ⅴ期,而卵巢(4+龄及以上龄期)要在5月中下旬到6月份才能发育至第Ⅴ期,说明滩头雅罗鱼雌雄鱼间性腺发育存在不同步现象,雄鱼要先于雌鱼达到性成熟,这在鲥鱼上也有相似的报道[8]。在每年5—7月滩头雅罗鱼排精后精巢逐渐退化至Ⅱ期,在9月末、10月初所取的雄性个体,经切片观察发现此时精巢又发育到第Ⅲ期,未发育成熟的2龄补充群体精巢发育到第Ⅱ期。在高寒地区每年从10月开始到翌年3月为冰封期,所以我们推测具备完整性周期的滩头雅罗鱼雄鱼以Ⅲ期越冬,而未发育成熟的2龄补充群体以Ⅱ期越冬。翌年4月开始随着气温回升,越冬期结束,滩头雅罗鱼重新开始摄食,精巢开始继续发育。
3.2 滩头雅罗鱼卵巢发育及产卵特点
滩头雅罗鱼是鲤科中目前已知的唯一溯河洄游生殖产卵的鱼类,在自然条件下,一般在每年的4—6月开始洄游到我国的绥芬河、图们江流域寻找合适的产卵场所进行产卵。而在黑龙江实验基地池塘养殖的滩头雅罗鱼,可能由于外界条件的因素不能满足其性腺发育的需要,而不能自然进行产卵。卵巢停滞在Ⅳ期而不能向第Ⅴ期过渡,必须在人工利用药物催产的条件下才能完成生殖细胞的最后成熟,这和同属于溯河洄游生殖的鲥鱼相似[8]。5—7月滩头雅罗鱼产完卵后,在9月末、10月初卵巢又发育到Ⅲ期,未性成熟个体(2+龄)卵巢发育到Ⅱ期,因此我们推测具备完整性周期的滩头雅罗鱼雌鱼以Ⅲ期越冬,而未发育成熟的2龄补充群体以Ⅱ期越冬,在光唇鱼(Acrossocheilusfasciatus)[14]和鲥鱼(Tenualosareevasii)[8,15]中也有相似的结果。卵巢按照卵母细胞的发育特点分为3种:完全同步型、部分同步型和不同步型[16-17]。完全同步型,卵巢内的卵母细胞都处于同一发育时期,通常一生只产卵一次就发生死亡,如大麻哈鱼(Oncorhynchusketa)[18]。部分同步型,卵巢内存在两种或两种以上的卵母细胞类型,通常一年只产卵一次,如草鱼(Ctenopharyngodonidellus)[19]、美洲鲥鱼(Alosasapidissima)[20]和虹鳟(Oncorhynchusmykiss)[21]。不同步型,卵巢内存在各种时相的卵母细胞类型,通常一年内产卵多次,如斑马鱼(Daniorerio)[22]、青鳉(Oryziaslatipes)[23]等。在本实验中发现,滩头雅罗鱼卵细胞从第3时相发育到第4时相基本上是同步的,在Ⅳ期卵巢中第4时相卵母细胞占卵总数的45.7%~78.6%左右,第3时相卵母细胞占0.7%~6.0%左右,其余为第1、2时相卵母细胞。在池塘养殖条件下,一年仅产卵一次,因此我们推测滩头雅罗鱼卵巢应属于部分同步型,即一年出现一次产卵高峰。
3.3 盐碱度对性腺发育的影响
对于溯河洄游产卵的鱼类来说,盐度是影响卵巢最后发育成熟及产卵的最重要因子。例如大麻哈鱼幼鱼在海水中摄食肥育,性成熟后会不远万里洄游到淡水中寻找合适的产卵场进行产卵[24]。在对梭鱼(Lizahaematocheila)[25]研究中发现,环境盐度可以影响垂体前叶和间叶的大小。在淡水或少盐水(盐度低于2‰)条件下,垂体前叶体积明显增大,催乳素细胞活动增强。同时,间叶变小,促性腺激素分泌减少。另外卵巢发育停滞在Ⅳ期初,不能继续发育,即使有个别个体能够发育成熟,经人工催产后也不能正常受精,究其原因可能是由于在低盐环境中鱼体内垂体内分泌平衡和激素平衡遭到破坏,使卵巢最终不能发育成熟并产卵;然而这种水体环境对雄性却并无影响,在生殖季节能够正常成熟并排精。另外不同盐度对黑鲷(Sparusmacrocephlus)[26]和鲻鱼(Mugilcephalus)[27]卵巢发育和孵化率都产生影响。有不少学者认为,较高的盐度、较大的水压和较低的水温都有利于卵巢的最后成熟[28]。对鳗鲡(Anguillajaponica)[29]来说正好相反,其性腺在淡水中发育成熟,降海洄游到较深的海洋中进行产卵。鳗鲡在不同盐度环境中养殖,性腺发育会受到不同程度影响。在本研究中发现在呼兰实验基地弱碱水池塘中,4~7龄滩头雅罗鱼卵巢能够发育成熟,在生殖季节经过人工催产能正常产卵并受精,未达到性成熟的2~3+龄群体性腺发育到Ⅱ~Ⅲ期,和淡水养殖条件下的2~3+龄群体并无区别。由于滩头雅罗鱼人工繁殖成功的时间较短,受到实验材料的限制,并未获得4龄及以上淡水养殖条件下的实验鱼,到底在淡水环境中卵巢能否发育到Ⅳ期中后期还尚未可知,因此在淡水养殖条件下滩头雅罗鱼能否用于人工催产还有待进一步研究才能做出准确判断。
4 小结
该研究首次对淡水和弱盐碱水养殖条件下的2~3龄滩头雅罗鱼性腺结构、性腺发育及个体生长进行了跟踪调查;并根据卵巢中细胞发育及其动态变化规律,确定了滩头雅罗鱼卵巢发育类型。9月末,滩头雅罗鱼幼鱼(2+龄)性腺发育到Ⅱ期,并以Ⅱ期越冬;性成熟成鱼在9月末性腺发育到第Ⅲ期,并以Ⅲ期越冬。性成熟的雄鱼性腺发育快于雌鱼。2~3龄个体不论在淡水或弱碱水环境中养殖,性腺发育时相并无明显区别。滩头雅罗鱼为一年一次性产卵类型。弱碱水2+龄群体体重显著高于淡水同龄群体,然而3+龄群体在弱碱水和淡水中体重无显著差异。弱碱水和淡水中的同龄雌、雄之间体重变化存在差异。总之,通过本研究,我们加深了对滩头雅罗鱼性腺发育和繁殖行为的了解,这为深入研究滩头雅罗鱼在盐碱水和淡水中性腺发育规律和个体生长差异提供了依据。
[1]董崇智, 夏重志, 姜作发, 等. 绥芬河滩头雅罗鱼的人工放流[J]. 黑龙江水产, 1997(3): 9-10.
[2]戴定远, 张玉玲, 黄浩明. 图们江滩头鱼的生物学研究[J]. 动物学报, 1982, 28(3): 283-292.
[3]杨荣国, 杨志江, 田 洋, 等. 勃氏雅罗鱼苗种繁育及成鱼养殖技术[J]. 河北渔业, 2012(1): 32-35.
[4]尹家胜, 沈俊宝, 王维坤. 滩头雅罗鱼繁殖策略的遗产进化特征[J]. 云南大学学报(自然科学版), 1999, 21(3): 223-224.
[5]尹家胜, 沈俊宝, 栾晓红. 温度对绥芬河滩头雅罗鱼胚胎发育的影响[J]. 生态学报, 1992, 12(3): 232-238.
[6]池炳杰, 梁利群, 刘春雷,等. 滩头雅罗鱼幼鱼对NaCl浓度和碱度的适应性分析[J]. 中国水产科学, 2011, 18(3): 689-694.
[7]窦新杰, 常玉梅, 唐 然, 等. 几种雅罗鱼亚科鱼类基于mtDNA序列的亲缘关系[J]. 江苏农业学报, 2014, 30(4): 826-832.
[8]王汉平, 魏开金, 姚 红, 等. 养殖鲥鱼性腺发育的研究[J]. 动物学报, 1998, 44(3): 314-321.
[9]BILLARD R. Spermatogenesis and spermatology of some teleost fish species[J]. Reprod Nutr Develop, 1986, 26(4): 877-920.
[10]温茹淑, 陈晓东, 方展强. 唐鱼精巢的组织学观察[J]. 四川动物, 2012, 31(3): 422-425.
[11]徐钢春, 聂志娟, 张呈祥, 等. 刀鲚精巢发育的组织学研究[J]. 华中农业大学学报, 2012, 31(2): 247-252.
[12]杨家云. 瓦氏黄颡鱼精巢发育及精子生物学研究[J]. 西南师范大学学报(自然科学版), 2005, 30(4): 719-724.
[13]熊 正. 孔雀鱼生殖生物学研究[D]. 厦门: 厦门大学, 2008.
[14]姜建湖, 戴海平, 竺俊全, 等. 养殖光唇鱼(Acrossocheilusfasciatus)卵巢发育的组织学观察[J]. 海洋与湖沼, 2013, 44(2): 348-354.
[15]张克俭, 童合一, 陈马康. 鲥鱼在中国东南近海生殖洄游期间性腺发育的研究[J]. 水产学报, 1990, 14(1): 55-59.
[16]施瑔芳. 鱼类性腺发育研究新进展[J]. 水生生物学报, 1988, 12(3): 248-258.
[17]林浩然. 鱼类生理学[M]. 广州: 广东高等教育出版社, 2007: 198.
[18]王继隆, 唐富江, 朱 翥, 等. 乌苏里江秋大麻哈鱼生殖洄游群体结构特征与分析[J]. 湖南农业科学, 2011(21): 120-123.
[19]刘 筠, 陈淑群, 王义铣, 等. 草鱼产卵类型的研究[J]. 水生生物学集刊, 1978, 6(3): 247-260.
[20]洪 磊, 李兆新, 陈 超, 等. 美洲鲥鱼卵巢发育规律和性类固醇激素变化研究[J]. 中国工程科学, 2014, 16(9): 86-92.
[21]蔡海瑶, 张艳萍, 娄忠玉, 等. 虹鳟卵巢发育的组织学研究[J]. 甘肃农业大学学报, 2011, 46(2): 17-22.
[22]王 晶, 王 冰, 李纪同, 等. 斑马鱼性腺发育的组织学观察[J]. 基因组学与应用生物学, 2011, 30(2): 168-174.
[23]张 静. 青鳉性腺发育的组织学观察和DMY、DMRT1基因的表达分析[D]. 上海: 上海海洋大学, 2013.
[24]温海深, 林浩然. 环境因子对硬骨鱼类性腺发育成熟及其排卵和产卵的调控[J]. 应用生态学报, 2001, 12(1): 151-155.
[25]蒋一珪, 周仲利, 俞志田, 等. 环境盐度对梭鱼脑下垂体及性腺发育的影响[J]. 水产学报, 1980, 4(3): 229-240.
[26]施兆鸿. 盐度对黑鲷卵巢发育的影响[J]. 水产学报, 1996, 20(4): 357-360.
[27]时红卫, 吴美琴, 张俊彬. 鲻鱼人工繁殖与胚胎发育观察[J]. 上海海洋大学学报, 2015, 24(6): 855-861.
[28]林浩然, 张梅丽, 张素敏, 等. 鳗鲡繁殖生物学研究Ⅳ. 人工催熟过程中下海鳗鲡的Gth分泌活动、性腺发育状况和脑垂体Gth细胞的超显微结构[J]. 水生生物学报, 1987, 11(4): 320-330.
[29]张 森. 盐度影响人工养殖日本鳗鲡性腺发育的机理研究[D]. 厦门: 集美大学, 2012.
Histological studies on the gonadal development and growth comparison ofTribolodonbrandtiindifferentcultureconditions
YANHao1,2,SUBao-feng1,CHANGYu-mei1,SUNBo1,SHANGMei1,ZHANGLi-min1,LIANGLi-qun1
(1. Heilongjiang Fisheries Research Institute, Chinese Academy of Fishery Sciences, Harbin 150070;2. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China)
S917.4
A
2095-1736(2017)05-0047-06
2016-09-20;
2016-12-01
国家自然科学基金项目(31461163004); 中国水产科学研究院基本科研业务费(2016RC-YJ02); 黑龙江省人力资源和社会保障厅“省留学回国人员择优资助”(黑人社函〔2015〕424号)
闫 浩,硕士,主要研究方向鱼类遗产育种与分子生物学,E-mail: yanhao_02@163.com
梁利群,博士,研究员,主要研究方向水产动物功能基因研究,E-mail: liangliqun@hrfri.ac.cn
doi∶10.3969/j.issn.2095-1736.2017.05.047
