沉默INK4基因座中反义非编码RNA对人宫颈癌细胞株HeLa增殖的影响及其机制
2017-10-10梅奇华杨鸿雁时黔宇李琼王卓赵鸿
梅奇华,杨鸿雁,时黔宇,李琼,王卓,赵鸿
(黔西南州人民医院,贵州黔西南州562400)
沉默INK4基因座中反义非编码RNA对人宫颈癌细胞株HeLa增殖的影响及其机制
梅奇华,杨鸿雁,时黔宇,李琼,王卓,赵鸿
(黔西南州人民医院,贵州黔西南州562400)
目的观察沉默INK4基因座中反义非编码RNA(ANRILA)对人宫颈癌细胞株HeLa增殖的影响,并探讨其机制。方法取对数生长期HeLa细胞分为观察组和对照组,分别ANRIL-siRNA、对照siRNA乱序。转染24、48 h时采用 CCK-8法观察两组细胞增殖情况(结果以OD450表示)。转染48 h时采用荧光定量PCR法检测两组细胞ANRIL、p21、周期蛋白依赖性激酶6(CDK6)、周期蛋白依赖性激酶2(CDK2) mRNA相对表达量,采用CHIP实验观察两组细胞p21基因H3K27位点三甲基化状态。结果转染24 h观察组、对照组OD450分别为0.55±0.08、0.84±0.11,转染72 h观察组、对照组OD450分别为1.21±0.12、1.82±0.14。转染24、72 h观察组OD450均低于对照组(P<0.05)。转染48 h观察组ANRIL及CDK6、CDK2 mRNA相对表达量低于对照组(P均<0.05),p21 mRNA相对表达量高于对照组(P均<0.05)。转染48 h两组细胞p21基因H3K27位点三甲基化水平观察组、对照组分别为1.19±0.08、5.62±0.21,观察组低于对照组(P<0.05)。结论沉默ANRIL可抑制宫颈癌HeLa细胞的增殖。其机制是沉默ANRIL可以降低宫颈癌HeLa细胞p21基因H3K27位点三甲基化水平,促进p21mRNA表达,抑制CDK6、CDK2 mRNA表达,从而抑制宫颈癌HeLa细胞增殖。
宫颈肿瘤;宫颈癌;长链非编码RNA;INK4基因座中反义非编码RNA;细胞增殖;p21基因;周期蛋白依赖性激酶;甲基化
Abstract:ObjectiveTo observe the effect of silencing ANRIL on the proliferation of human cervical cancer cell line HeLa and to explore its mechanism.MethodsThe HeLa cells in the logarithmic phase were divided into the observation group and the control group, which were transfected with ANRIL-siRNA and control siRNA, respectively. After 24 and 48 h transfection, the proliferation of the two groups was observed by CCK-8 method (the result was expressed by OD450). The expression of ANRIL, p21, cell cycle dependent kinase 6 (CDK6) and cyclin-dependent kinase 2 (CDK2) mRNA was detected by fluorescence quantitative PCR at 48 h after transfection, and CHIP assay was used to detect H3K27me3 in p21 gene of the two groups.ResultsAt 24 h after transfection, OD450of the observation group and the control group were 0.55±0.08 and 0.84±0.11, respectively. At 72 h, OD450of the observation group and the control group were 1.21±0.12 and 1.82±0.14, respectively. The OD450of the observation group was lower than that of the control group at 24 and 72 h after transfection (allP<0.05). The mRNA expression of ANRIL, CDK6, and CDK2 in the observation group was lower than that in the control group at 48 h after transfection (allP<0.05), while the expression of p21 in the observation group was higher than that in the control group (P<0.05). H3K27me3 level of p21 gene in the observation group was 1.19±0.08, which was lower than that (5.62±0.21) of the control group at 48 h (P<0.05).ConclusionSilencing ANRIL can inhibit the proliferation of cervical cancer HeLa cells by reducing the H3K27me3 level in p21 gene, and thus promoting the expression of p21 and inhibiting CDK6 and CDK2 mRNA expression.
Keywords: cervical neoplasms; cervical carcinoma; long non-coding RNA; noncoding RNA in antisense of INK4 loci; p21 gene; cyclin-dependent kinase; methylation
宫颈癌的发病机制尚不明确[1~3],目前仍是临床研究的热点。一些长链非编码RNA(LncRNA)参与了多种肿瘤的发生发展过程[4~8]。INK4基因座中反义非编码RNA(ANRIL)是一条由19个外显子组成的LncRNA,终产物长度约3 834个碱基,位于p15-p16-p14基因座且与蛋白编码基因呈反向转录[9]。我们前期研究发现,宫颈癌组织中ANRIL高表达,提示ANRIL可能参与了宫颈癌的发生发展,但其发挥的具体作用及作用机制尚不明确。为此,本研究观察了沉默和过表达ANRIL对宫颈癌HeLa细胞增殖的影响,并探讨其机制。现报告如下。
1 材料与方法
1.1 细胞、试剂及仪器 人宫颈癌HeLa细胞株购自上海中科院生化细胞所细胞库,培养基为DMEM培养基+10%胎牛血清,ANRIL siRNA由广州锐博生物技术有限公司合成。siRNA瞬时转染试剂均采用Invitrogen公司的Lipofectamine 2000包裹。PCR荧光染料为TAKATA SYBR Green,Ki67、H3K27me3抗体购自美国abcam公司,KLF2、p21抗体购自美国Millipore公司。蛋白提取采用碧云天公司RIPA中型裂解液,RNA提取采用TAKARA公司TRIzol试剂,逆转录试剂盒购自TAKARA公司,细胞增殖检测采用日本同仁公司CCK-8试剂盒,引物由上海上海生工生物技术有限公司合成。RIP试剂盒为Millipore公司Magna RIP RNA-Binding Protein Immunoprecipitation Kit(17-700),CHIP试剂盒购自Millipore公司(17-295),G蛋白磁珠购自Santa Cruz公司。细胞凋亡检测试剂盒购自Roche公司。荧光定量PCR仪、检测酶标仪购自Bio-Rad公司。
1.2 HeLa细胞分组及ANRIL-siRNA转染 取对数生长期HeLa细胞,培养至细胞密度约70%时换为无血清培养基饥饿2 h, 设观察组和对照组。观察组转染ANRIL-siRNA,对照组转染Lipofectamine 3000包裹的对照siRNA乱序(NC)。将转染后细胞置于5 % CO2、37 ℃培养箱中培养。
1.3 两组细胞增殖情况观察 分别于转染24、48 h时采用 CCK-8法检测两组细胞增殖情况。操作按试剂盒说明书进行。采用荧光酶标仪检测450 nm处光密度(OD450)值,以此表示各组细胞增殖情况。
1.4 两组细胞ANRIL及p21、CDK6、CDK2mRNA检测 采用荧光定量PCR法,取转染48 h各组细胞,采用TRIzol法提取总RNA,逆转录PCR采用TAKARA公司RT试剂盒、20 μL体系进行反转录。以GAPDH作为内参,引物序列为:ANRIL上游引物5′-CAACATCCACCACTGGATCTTAACA-3′;下游引物5′-AGCTTCGTATCCCCAATGAGATACA-3′。 p21上游引物5′-GGAGGCAAAAGTCCTGTGTTC-3′;下游引物5′- GGAAGGGGAGGATTTGACGA-3′。CDK6上游引物5′-TGGTAACTCCTTCCCCAGAGT-3′;下游引物5′-CGAAGCGAAGTCCTCAACAC-3′。CDK2上游引物5′-CTTTGCTGAGATGGTGACTCG-3′;下游引物5′-TCATCCAGGGGAGGTACAACT-3′。GAPDH上游引物5′-AATGGGCAGCCGTTAGGAAA-3′;下游引物5′-GCGCCCAATACGACCAAATC-3′。实验条件为92 ℃预变性3 min,92 ℃ 30 s、58 ℃、10s,共39个循环,72 ℃ 2 min。用2-ΔΔCt表示ANRIL的相对表达量。
1.5 两组细胞p21基因H3K27位点三甲基化水平观测 采用CHIP实验。取各组细胞,醛固定交联后,细胞裂解液裂解细胞,超声破碎,加入H3K27me3抗体孵育过夜,过夜后加入磁珠捕获蛋白DNA结合的复合物,随后使用洗涤液洗涤,再加入5 mmol/L NaCl解交联,解交联结束后回收DNA,采用荧光定量PCR法检测复合物中ANRIL相对表达量。方法同1.3。以复合物中ANRIL相对表达量表示两组细胞p21基因H3K27位点三甲基化水平。
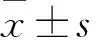
2 结果
2.1 转染24、72 h两组细胞OD450比较 转染24 h观察组、对照组OD450分别为0.55±0.08、0.84±0.11,转染72 h观察组、对照组OD450分别为1.21±0.12、1.82±0.14。转染24、72 h时观察组OD450均低于对照组(P<0.05)。
2.2 转染48 h两组细胞ANRIL及p21、CDK6、CDK2 mRNA相对表达量比较 转染48 h两组细胞ANRIL及p21、CDK6、CDK2 mRNA相对表达量见表1。转染48 h观察组ANRIL及CDK6、CDK2 mRNA相对表达量低于对照组(P均<0.05),p21 mRNA相对表达量高于对照组(P<0.05)。
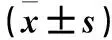
表1 转染48 h两组细胞ANRIL及p21、CDK6、CDK2 mRNA相对表达量比较
注:与对照组相比,*P<0.05。
2.3 转染48 h两组细胞p21基因H3K27位点三甲基化水平比较 转染48 h两组细胞p21基因H3K27位点三甲基化水平观察组、对照组分别为1.19±0.08、5.62±0.21,观察组低于对照组(P<0.05)。
3 讨论
LncRNA是一类长度超过200 bp的转录本,具有复杂的二级结构,相较于microRNA,其作用方式更为复杂[11],例如HotAir、Xist等可以与蛋白修饰复合体互作,影响组蛋白H3K27表观遗传学改变[12],而H19则可以作为分子筛,发挥类似海绵作用吸附microRNA分子[13],最终影响靶mRNA。有别于microRNA分子相对单一的抑制作用,LncRNA既可促进基因的转录,也可以抑制转录;既可以结合核酸,也可以结合蛋白;既可以修饰蛋白质,也可以影响mRNA的成熟和稳定。近期越来越多的研究者发现,LncRNA在肿瘤的演进中扮演关键角色,与肿瘤的增殖、凋亡、迁移、侵袭密切相关,例如 HOTTIP通过调控HOXA13基因转录参与肝癌增殖、侵袭等变化,并与患者的预后相关[13];GIHCG可以通过调控miR-200b/a/429促进肝癌进展[14];MALAT1通过沉默miR-140促进葡萄膜黑素瘤细胞增殖和迁移[15];ANRIL作为发现较早的分子,被发现可以作为癌基因促进多个肿瘤的增殖、侵袭[16]。然而,宫颈癌相关的LncRNA研究却非常少。
本研究结果显示,转染ANRILsiRNA 的HeLa细胞增殖能力下降。进一步分析发现,沉默ANRIL后,细胞周期监测点蛋白p21 mRNA上调,而CDK2、CDK6下调。p21基因也被称为细胞周期依赖性激酶抑制剂1或CDK反应蛋白1,位于p53基因下游的细胞周期素依赖性激酶抑制因子,p21和p53一起构成了细胞周期G1检查站[17],假如肿瘤细胞因快速增殖而产生了DNA损伤,而这些损伤不能得到及时的修复,那么该细胞则无法通过G1检查站。p21通过这一作用减少了受损DNA的复制和积累,从而协同p53发挥抑癌作用。CDK2是重要的细胞周期推动因素,属于丝氨酸/苏氨酸激酶家族,也被称为细胞分裂蛋白激酶2,调控细胞周期由G1到S期转换。CDK6也是细胞周期由G1到S期转换的主要推动因素之一,受到细胞周期蛋白cyclin D的调控。由此可知, ANRIL通过调控p21转录,进一步调控细胞周期依赖性激酶CDK2、CDK6的表达,从而操控细胞周期,发挥癌基因的作用。
ANRIL主要定位于细胞核内,提示其可与蛋白交互作用发挥功能,既往研究提示 ANRIL可引起多个基因的启动子区组蛋白表观遗传学印记改变,从而调控基因转录。组蛋白H3的不同赖氨酸残疾的甲基化/乙酰化状态对基因的转录激活或抑制发挥着不同的作用, H3K27位点三甲基化抑制基因转录。本研究结果显示,沉默ANRIL后HeLa细胞p21基因H3K27位点三甲基化水平降低,这应该是HeLa细胞p21 mRNA表达上调的原因。
[1] Stuwe E, Tóth KF, Aravin AA. Small but sturdy: small RNAs in cellular memory and epigenetics[J]. Genes Dev,2014,28(5):423-431.
[2] Zhang C, Gao L, Xu EY. LncRNA, a new component of expanding RNA-protein regulatory network important for animal sperm development[J]. Semin Cell Dev Biol, 2016,59(16):30175-30176.
[3] Sullenger BA, Nair S. From the RNA world to the clinic[J]. Science, 2016,352 (6292):1417-1420.
[4] Duval M, Cossart P, Lebreton A. Mammalian microRNAs and long noncoding RNAs in the host-bacterial pathogen crosstalk[J]. Semin Cell Dev Biol, 2016,9521 (16):30162-301628.
[5] Chen LL. Linking long noncoding RNA localization and function[J]. Trends Biochem Sci, 2016,41 (9):761-772.
[6] Schmitt AM, Chang HY. Long noncoding RNAs in cancer pathways[J]. Cancer Cell, 2016,29(4):452-463.
[7] Thomson DW, Dinger ME. Endogenous microRNA sponges: evidence and controversy[J]. Nat Rev Genet,2016,17(5):272-283.
[8] 张珊,洪颖,马莉,等.宫颈细胞学阴性HPV阳性妇女的HPV分型检测[J].山东医药,2015,55(43):5-7.
[9] Xu S, Wang H. ANRIL lncRNA triggers efficient therapeutic efficacy by reprogramming the aberrant INK4-hub in melanoma[J]. Cancer Lett, 2016,381(1):41-48.
[10] Nie FQ, Sun M. Long noncoding RNA ANRIL promotes non-small cell lung cancer cell proliferation and inhibits apoptosis by silencing KLF2 and P21 expression[J]. Mol Cancer Ther, 2015,14(1):268-277.
[11] Quinn JJ, Chang HY. Unique features of long non-coding RNA biogenesis and function[J]. Nat Rev Genet, 2016,17(1):47-62.
[12] Amort T, Soulière MF. Long non-coding RNAs as targets for cytosine methylation[J]. RNA Biol, 2013,10(6):1003-1008.
[13] Shi X, Sun M, Liu H, et al. Long non-coding RNAs: a new frontier in the study of human diseases[J]. Cancer Lett, 2013,339(2):159-166.
[14] Sang Y, Zhou F. Up-regulation of long non-coding HOTTIP functions as an oncogene by regulating HOXA13 in non-small cell lung cancer[J]. Am J Transl Res, 2016,8(5):2022-2032.
[15] Sui CJ, Zhou YM. Long noncoding RNA GIHCG promotes hepatocellular carcinoma progression through epigenetically regulating miR-200b/a/429[J]. J Mol Med (Berl), 2016,94(11):1281-1296.
Effect of silencing ANRIL on proliferation of human cervical cancer cell line HeLa
MEIQihua,YANGHongyan,SHIQianyu,LIQiong,WANGZhuo,ZHAOHong
(Qianxi'nanPeople'sHospital,Qianxi'nan562400,China)
10.3969/j.issn.1002-266X.2017.35.004
R737.33
A
1002-266X(2017)35-0012-03
2016-10-26)
梅奇华(1972-),女,主治医师,主要研究方向为宫颈癌、癌前病变、围产科学。E-mail: meiqihua111@sina.com
