籼型杂交水稻农艺性状的配合力研究
2017-09-25征张雪丽莫博程代志军胡中立李兰芝郑兴飞湖南省重要有害生物防控协同创新中心湖南农业大学植物保护学院湖南长沙408武汉大学杂交水稻国家重点实验室湖北武汉43007
张 征张雪丽莫博程代志军胡中立李兰芝,*郑兴飞,*湖南省重要有害生物防控协同创新中心 / 湖南农业大学植物保护学院, 湖南长沙 408;武汉大学杂交水稻国家重点实验室, 湖北武汉 43007
籼型杂交水稻农艺性状的配合力研究
张 征1张雪丽1莫博程1代志军1胡中立2李兰芝1,*郑兴飞2,*
1湖南省重要有害生物防控协同创新中心 / 湖南农业大学植物保护学院, 湖南长沙 410128;2武汉大学杂交水稻国家重点实验室, 湖北武汉 430072
如何有效利用杂种优势已成为水稻增产的关键。本研究按照NCII遗传交配设计, 将三系野败型杂交水稻的恢复系和微核心种质构成的115份优异籼稻品种, 分别与4个两系不育系及1个三系不育系测交, 分析各农艺性状配合力、遗传力及相互关系。结果表明, 除单株有效穗数、主穗实粒数外, 其他农艺性状一般配合力差异均达到极显著水平; 除单株有效穗数外, 其他各农艺性状特殊配合力差异也均达到极显著水平。同一组合的不同性状、同一亲本的不同组合所表现出的特殊配合力效应都有所不同, 表明亲本的一般配合力水平与特殊配合力间没有固定的联系。在育种实践中, 选取一般配合力高的亲本, 同时兼具较高特殊配合力是获取高产杂交稻组合的关键。
杂交水稻; 三系野败型; NCII设计; 配合力; 杂种优势
如何有效利用杂种优势是水稻增产的关键。一直以来对杂种优势的遗传基础存在争论, 不同遗传设计、不同作物群体中得到了不同的结论[1-10]。1998年, Monforte等[2]利用5个来自不同野生种的染色体代换系(又称导入系)培育新的重组自交系, 研究产量性状和果实形状与不同遗传背景、环境间的关系。结果表明, 番茄产量的杂种优势与单个位点的超显性无关, 代换系片段之间的互作才是产量杂种优势的原因。2007年, 余传元等[4]以染色体单片段代换系和粳稻测验种, 按North Carolina design II (NCII)设计, 进一步了解杂交稻亲本选择的基本规律。认为应当在双亲一般配合力(general combining ability, GCA)均高的基础上, 保证较高的特殊配合力(specific combining ability, SCA)才是高产杂交水稻亲本选择的基本原则。2015年, Liu等[5]利用广陆矮4号和特青产生的F2回交群体与不育系, 按NCII设计进行测交。发现一般配合力较低的亲本, 子代的株高、抽穗期、颖花数这3个性状反而呈现超表达,且其一般配合力效应显著上升。2016年, Holtrop[6]以硬粒型和马齿型为亲本, 按NCII设计, 在玉米的杂交育种中发现, 产量性状被加性效应控制, 且产量的稳定性和潜力与玉米的种植密度没有明显联系。2009年, Huang等[7]利用油菜籽(Brassica napus L.)隐性雄性不育系和近交系, 按NCII设计, 获得的单交种表明所测性状的GCA和SCA都存在显著差异。建议油菜育种者选择GCA和SCA效应力高的组合, 培育高产杂交后代。以作图群体为基础对水稻、番茄、玉米、油菜等作物的配合力和杂种优势研究表明, 高产量杂交后代的培育理论在不同作物不同品种中存在明显的差异。
在水稻自然品种群体的研究中, NCII设计也被广泛认为是进行配合力和杂种优势研究的经典遗传设计。付新民等[8]利用野败型雄性不育系和恢复系,按照NCII遗传交配设计表明, 在水稻培育中, 应当根据不同性状考虑不育系和恢复系对农艺性状的影响, 进一步提高杂交水稻的杂种优势水平。梁康迳[9]采用包括现今生产上大面积推广的籼型恢复系在内组成的籼粳杂交水稻遗传群体, 按NCII设计, 主要分析其穗部性状的杂种优势的遗传规律, 建议适当扩大双亲的遗传差异, 以保持杂种优势水平, 并与培育目标和组合测配特点相结合, 有效地培育适合不同环境或特定环境的超高产组合。
本研究致力于探寻水稻杂种优势研究中广泛应用的籼型杂交水稻中的配合力效应, 提高杂交水稻的育种效率, 培育超高产杂交水稻。按照NCII遗传交配设计, 构建测交群体, 考察包括产量性状在内的 9个农艺性状。通过对配合力效应的相关参数分析, 系统评价品种间的配合力水平, 分析水稻杂种优势的育种基础, 以便更好地丰富水稻杂种优势育种理论[11-12]。
1 材料与方法
1.1 研究材料
部分材料由华中农业大学余四斌教授、四川农业大学李双成教授和扬州大学汤述翥教授馈赠。其中含29份三系野败型杂交水稻的恢复系和86份微核心种质构成品种群体。4个两系不育系[矮64S (PA64)、广占63S (GZ63)、Y58S和新安S(AS)]及1个三系不育系[珞红 3A(3A)]构成测交不育系。以上材料均为籼稻(详见附表1)。
1.2 试验设计
根据 NCII遗传交配设计, 在海南陵水开展杂交试验, 构建测交群体, 共690个株系(115份亲本+575F1)。2011年和 2012年分别在湖北鄂州、海南陵水两地进行杂交, 获得F1杂交种。田间种植1行10株,种植密度16.7 cm×26.7 cm, 于2013年分别种植在华中农业大学水稻基地(试点 1)和湖北省农业科学院(试点2)两点, 每点两次重复, 随机区组设计种植。
1.3 测定项目及方法
水稻成熟后于田间考察株高, 收种时, 从中间8株中选择长势均一的 3株收种, 室内考察单株有效穗数、主穗长、主穗一次枝梗数、主穗二次枝梗数、株高、主穗实粒数、千粒重、单株实粒重、主穗颖花数 9个性状。相对竞争优势值(RCH relative competition heterosis, %)= (F1-CK)/CK×100。
1.4 数据处理方法
利用SPSS软件完成[13]各亲本及测交F1的农艺性状方差分析。按照NCII设计, 参照莫惠栋老师的方法[14], 分析各个农艺性状一般配合力和特殊配合力。采用Microsoft Excel软件对各个农艺性状进行相关分析[15-16]。
2 结果与分析
2.1 主要农艺性状的表型值及配合力方差分析
表1表明, 在重复间, 所有农艺性状表型F值均未达到显著水平, 说明这些性状表型均比较稳定;不同组合间, 各个性状的差异均达到极显著水平,说明各个处理间存在明显的遗传差异。除了单株有效穗数及主穗实粒数外, 所有性状的一般配合力方差都达极显著水平。单株有效穗数特殊配合力方差未达到显著水平, 提示此性状主要受加性效应的影响。其他性状受加性效应、非加性效应的共同影响。
表2表明, 在重复间, 千粒重、株高及主穗二次枝梗数达到显著水平, 可能是由于肥力条件不均一,加上其他环境因素造成了不同重复间差异较大; 所有性状组合间差异均为极显著; 所有性状的一般配合力方差、特殊配合力方差都达极显著水平。
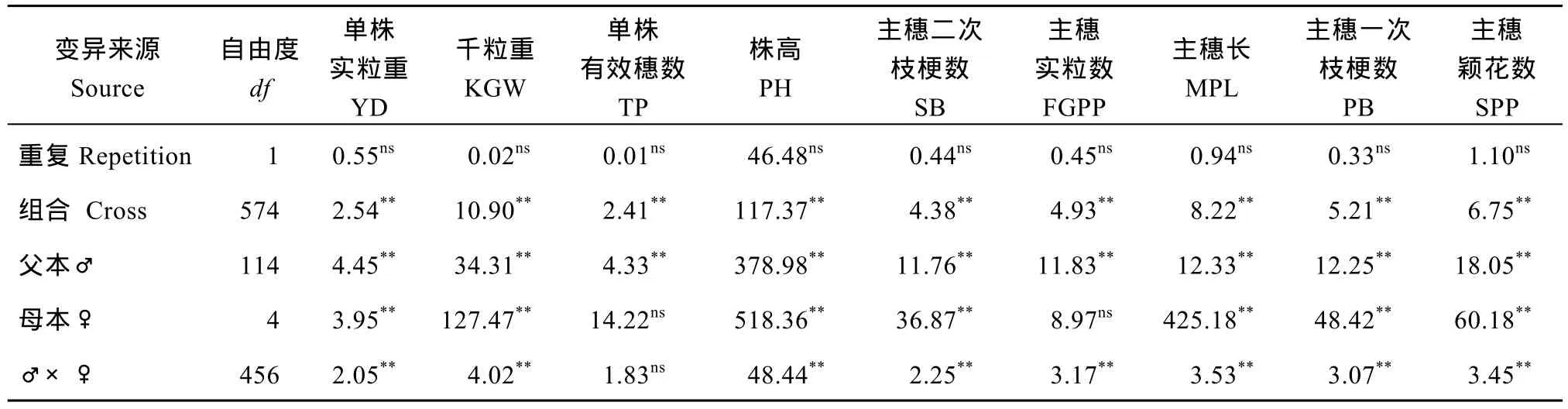
表1 试点1主要农艺性状的表型值及配合力的方差分析Table 1 Variance analysis of the phenotypic values and combining ability of main agronomic traits in Location 1
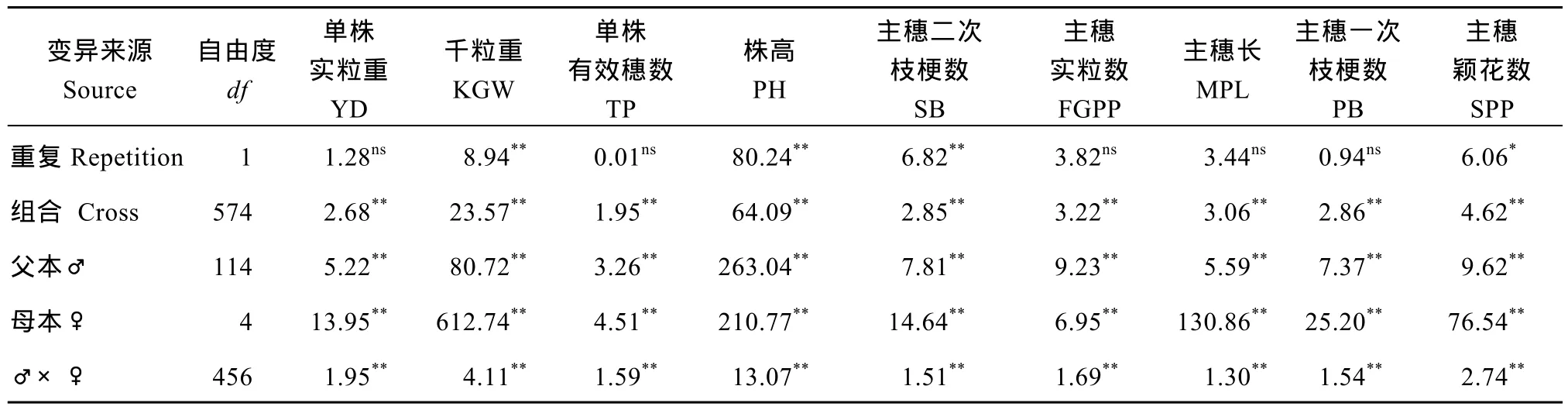
表2 试点2主要农艺性状的表型值及配合力的方差分析Table 2 Variance analysis of the phenotypic values and combining ability of main agronomic traits in Location 2
2.2 亲本主要农艺性状一般配合力效应分析
两试验点农艺性状的一般配合力在同一亲本的不同性状间及同一性状不同亲本间存在显著差异。在试点 1, 单株实粒重为-23.40~21.90, 其中最高的3个亲本为V81、V93、V72; 千粒重为-4.36~3.89, 其中最高的3个亲本为V108、V94、V101, 这些品种均为优良的恢复系材料, 可见在恢复系品种选育时千粒重对产量的重要性; 单株有效穗数为-4.28~5.88, 最高的 3个亲本为 V76、V1、V93; 株高为-23.05~43.21, 最高的3个亲本是V84、V17、V75; 主穗二次枝梗数为-17.21~34.97, 最高的亲本为V69、V89、V72; 主穗实粒数-60.77~71.65, 效应值除V83为 136.10外, 最高的亲本为 V72, V81; 主穗长为–3.81~3.73, 最高的亲本为V99、V60、V17; 主穗一次枝梗数为–2.06~4.61, 最高的亲本为V86、V2、V83;主穗颖花数为–69.84~79.03, 除 V83的值为 175.31外, 较高的亲本为V72、V71。
在试点 2, 单株实粒重–15.28~22.06, 最高的亲本依次为V43、V105、V82。千粒重为–6.27~4.53, 最高的亲本为 V109、V104、V101; 单株有效穗数为–3.26~4.13, 最高的亲本为 V12、V93、V1; 株高为–46.64~50.94, 最高的亲本为V90、V43、V35; 主穗二次枝梗数为–19.77~30.70, 最高的亲本为 V89、V69、V82; 主穗实粒数为–82.68~86.26, 除 V83的值为122.70外, 最高的亲本为V82、V86; 主穗长为–3.02~5.87, 最高的为V56、V60、V84; 主穗一次枝梗数–2.42~2.28, 除了最高的V71值为6.11, 其次是V69、V72; 主穗颖花数–25.00~56.76, 最高的3个依次是V90、V73、V69。
综上, 千粒重一般配合力较高的均是恢复系,单株实粒重、主穗实粒数一般配合力恢复系也较高。可见, 在恢复系选育过程中, 千粒重、单株实粒重、主穗实粒数是核心性状。
2.3 主要农艺性状特殊配合力分析
从表 3和表 4可以看出, 特殊配合力的效应值在不同组合、不同性状间均存在显著差异。且同一组合的不同性状、同一亲本的不同组合所表现出的特殊配合力效应都有所不同。分述如下: (1) 从亲本GCA组合上分析, 双亲 GCA较低也可以产生高的特殊配合力, 如试点1的V82/Y58S主穗二次枝梗数,试点 1的 V87/GZ63主穗一次枝梗数, 试点 2的V46/PA64主穗实粒数等; 双亲高 GCA组合也会产生较低的特殊配合力, 如试点1的V46/Y58S单株有效穗数, 试点 2的V107/GZ63单株实粒重, 试点2的V98/GZ63单株有效穗数等; (2) 3个最高、最低的SCA组合, 其双亲的GCA高低水平组合可有多种类型; (3)同一亲本组合, 在不同性状间SCA可能相差较大, 如试点1的V43/GZ63组合单株有效穗数SCA属于最低的3个之一, 但其株高SCA属于最高的3个之一, 还包括试点2的V46/PA64和V90/3A组合; (4)在3个最高的SCA组合中, 多个性状间均有高的SCA组合, 如试点1的V25/PA64在单株实粒重、千粒重、单株有效穗数、株高、主穗二次枝梗数、主穗实粒数、主穗长、主穗一次枝梗数; 在 3个最低的SCA组合中, 同一个组合的不同性状间也有较低的SCA, 如试点2的V82/Y58S组合的主穗二次枝梗数、主穗实粒数、主穗长。上述结果说明, 亲本的一般配合力类型与亲本组合的特殊配合力效应大小之间并无内在联系, 且在同一亲本组合中不同性状的特殊配合力有所不同, 但相关性状的特殊配合力可能会相似。
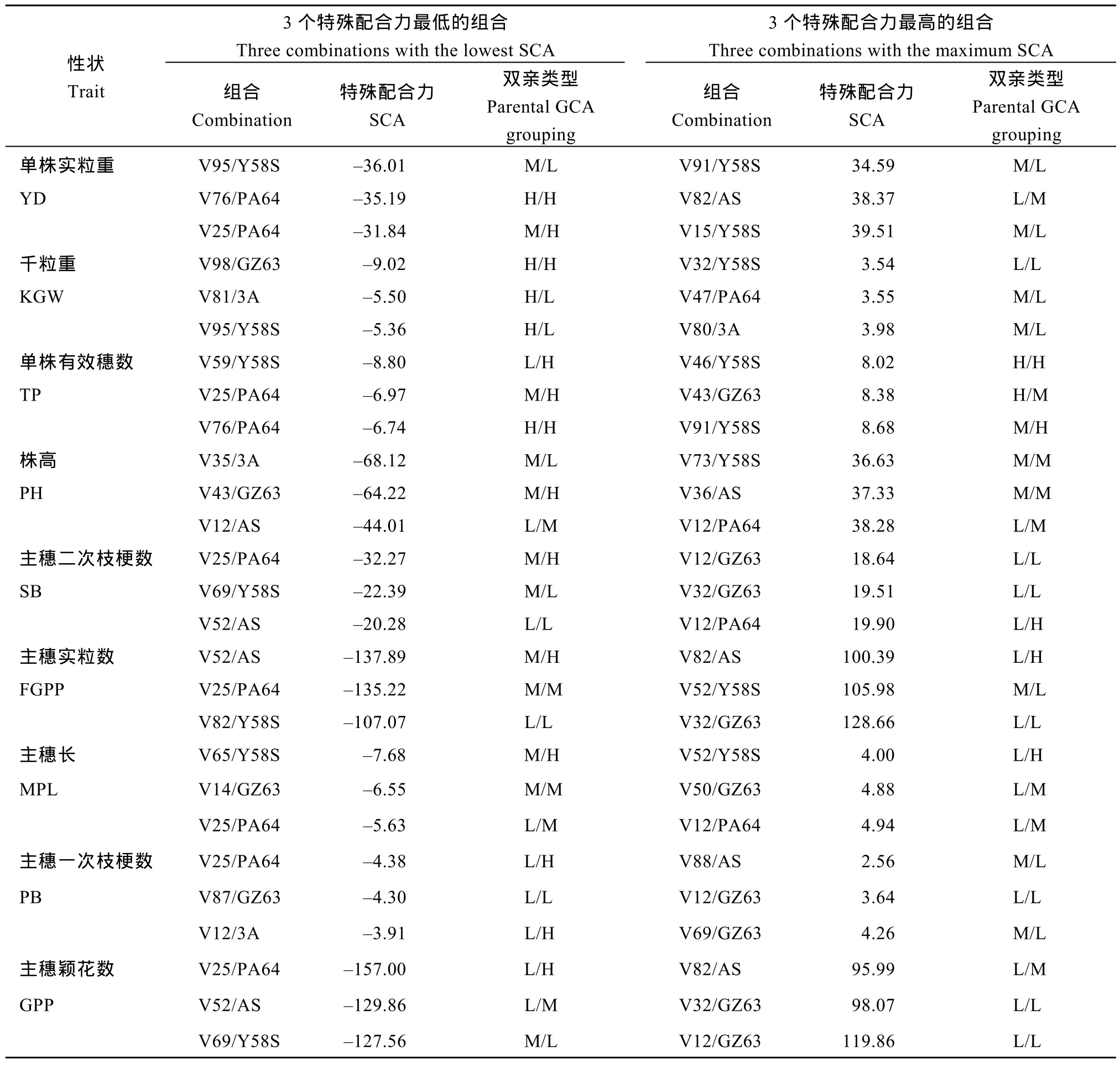
表3 试点1各性状最大、最小特殊配合力组合及双亲一般配合力的类别组合之间的关系Table 3 Top three and bottom three combinations of SCA and the relationship between SCA and parental GCA grouping in Location 1
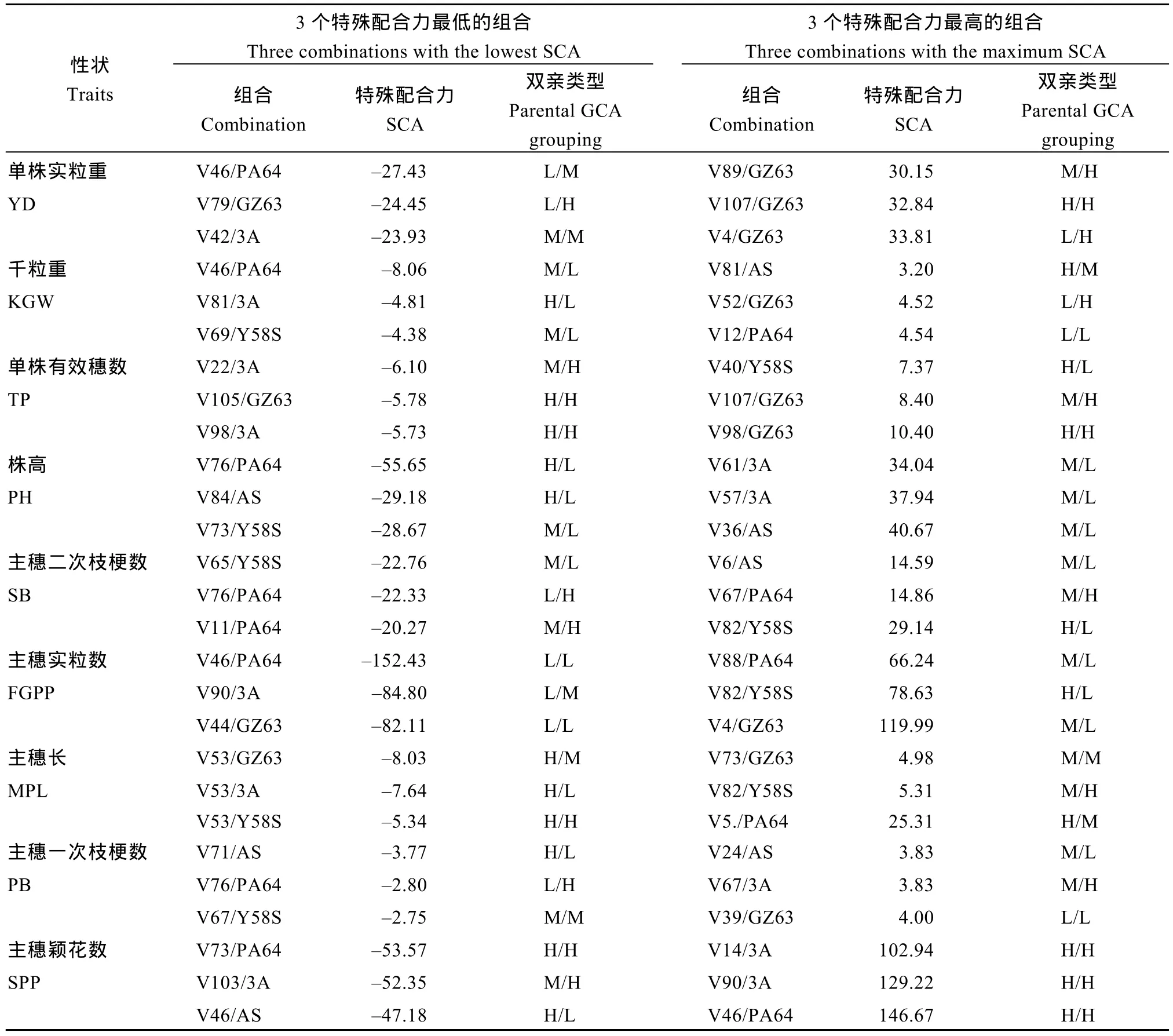
表4 试点2各性状最大、最小特殊配合力组合及双亲一般配合力的类别组合之间的关系Table 4 Top three and bottom three combinations of SCA and the relationship between SCA and parental GCA grouping in Location 2
2.4 主要农艺性状基因型方差、贡献率和遗传力
从表 5可以看出, 除单株实粒重、单株有效穗数及主穗实粒数外, 其余农艺性状的一般配合力方差(Vgca)占比均大于 50%, 其中千粒重的 Vgca最高,达70.2%。这些性状中, 亲本基因间加性效应对杂种F1的性状起主要作用; 从亲本的一般配合力方差来看, 父本在千粒重、单株有效穗数、株高的一般配合力较重要, 而母本的一般配合力(V♀)中主穗长占比高达46.1%, 其他性状占比均低于11%, 这一结果可能与选用的不育系穗型较长特性有关。特殊配合力方差(Vsca)在单株实粒重、单株有效穗数、主穗实粒数的比重高于 50%, 其余性状均低于 40%, 千粒重的特殊配合力方差最小, 说明单株实粒重、单株有效穗数、主穗实粒数主要来自基因间的非加性效应。考察的9个农艺性状均存在高的广义遗传力(HB),单株实粒重、单株有效穗数的狭义遗传力(HN)均小于 20%。可见, 单株实粒重、单株有效穗数受外部环境的影响较大。
从表 6可见, 除单株实粒重、单株有效穗数外,其余性状 Vgca占比均大于 50%, 其中千粒重最高,其Vgca高达86.9%; 因此, 这些性状中, 亲本基因间的加性效应对杂种F1的性状起主要作用。从亲本的一般配合力方差来看, 父本在千粒重、株高、主穗实粒数的一般配合力较重要, 而 V♀中主穗长占比较高, 达49.3%, 其他性状占比均低于22%。Vsca在单株实粒重、单株有效穗数的比重高于 50%, 其余性状均低于 50%, 千粒重、主穗长的特殊配合力方差最小, 说明单株实粒重、单株有效穗数主要来自于基因间的非加性效应。考察的 9个农艺性状, 均存在较高的广义遗传力(HB), 单株实粒重、单株有效穗数的狭义遗传力(HN)均小于 20%。可见, 单株实粒重、单株有效穗数受外部环境的影响较大。
综上可见, 两试验点的基因型方差、贡献率和遗传力趋势均一致。
2.5 一般配合力与杂种各农艺性状间的相关性分析
所有农艺性状与GCA均呈正相关; 除了单株实粒重外, 其他农艺性状与一般配合力间相关性均达到极显著水平(表7), 说明农艺性状千粒重、单株有效穗数、株高、主穗二次枝梗数、主穗实粒数、主穗长、主穗一次枝梗数、主穗颖花数, 其基因的加性效应对杂种 F1的性状表现有重要影响。这与高产杂交组合, 其亲本也多具有优异的性状表型结果一致。
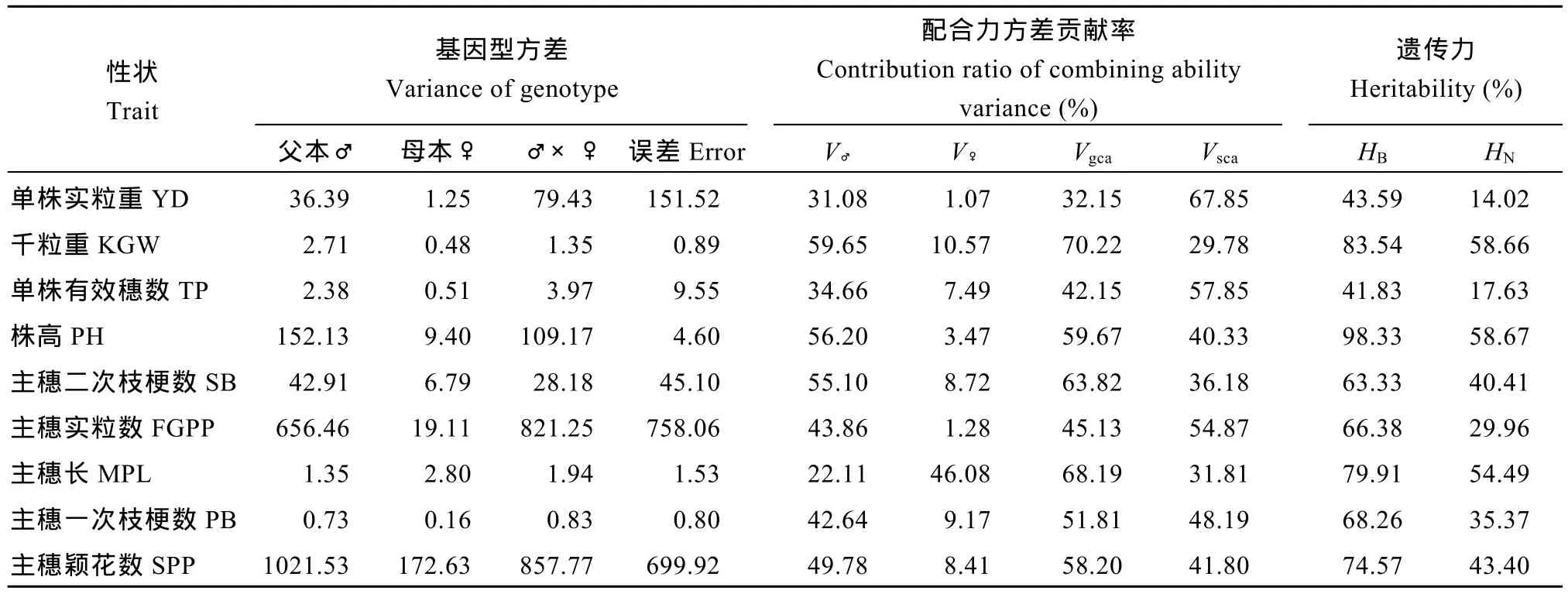
表5 试点1主要农艺性状的基因型方差、父母本及其基因互作对F1各性状方差的贡献率和遗传力Table 5 Genotypic variance for nine traits and the contribution ratio of the male, female parents and their interaction to the total variance of the traits in F1hybrids at Location 1
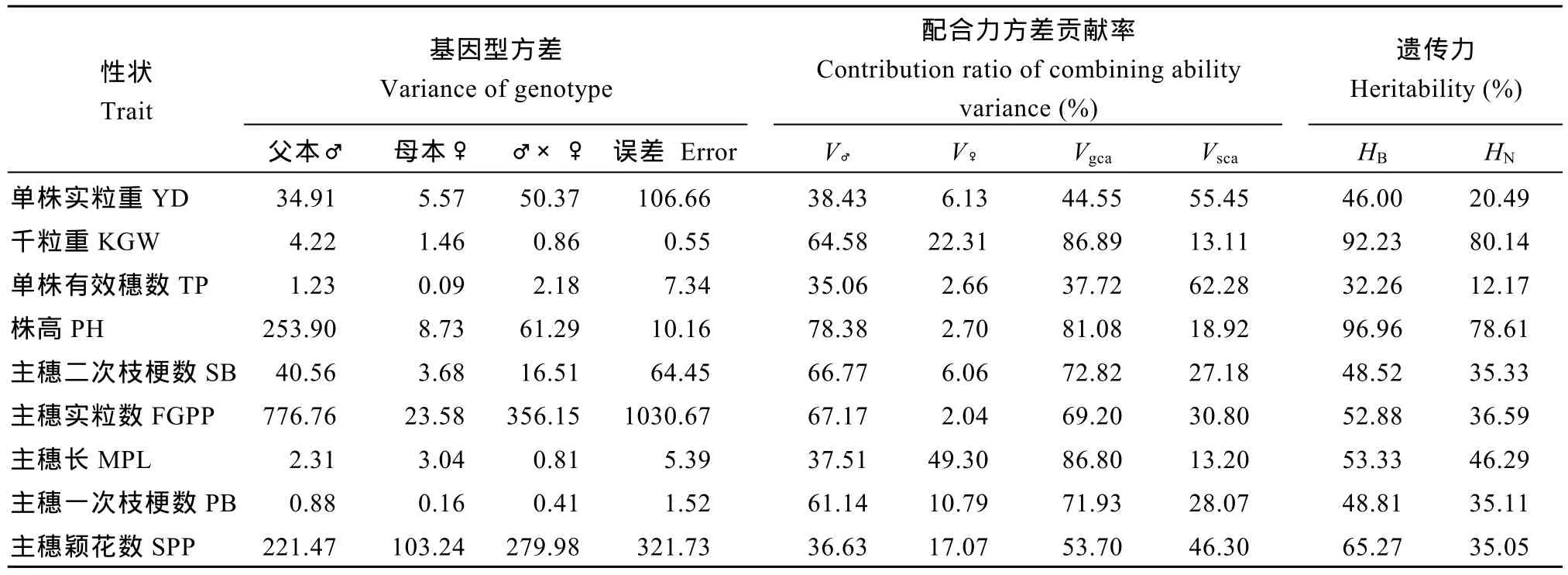
表6 试点2主要农艺性状的基因型方差、父母本及其基因互作对F1各性状方差的贡献率和遗传力Table 6 Genotypic variance for nine traits and the contribution ratio of the male, female parents and their interaction to the total variance of the traits in F1hybrids at Location 2
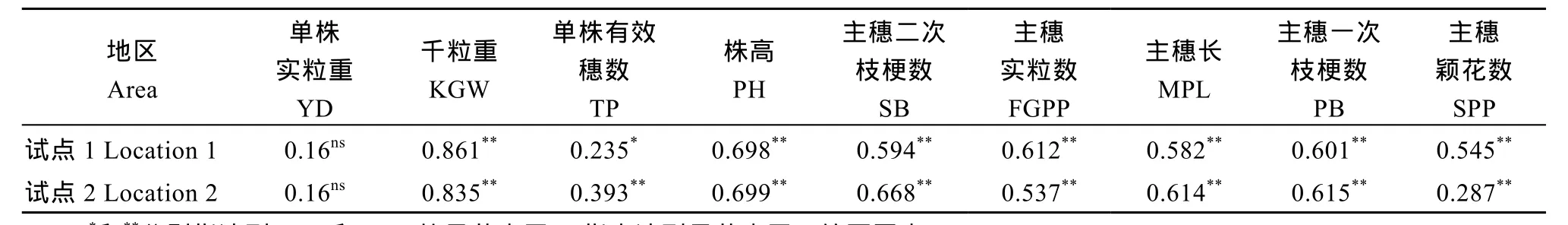
表7 各性状表型值与一般配合力效应的相关系数Table 7 Correlation coefficients (Pcc) between phenotypic values and GCA
2.6 配合力与杂种各农艺性状竞争优势值间相关性分析(RCH)
以品种 93-11为对照, 分析杂交 F1各性状相对竞争优势值与亲本GCA之和(GCA♂+♀)及SCA的相关性。从表8和表9可以看出, 亲本GCA之和与相对竞争优势在千粒重、株高、主穗二次枝梗数、主穗实粒数、主穗一次枝梗数、主穗颖花数性状间均呈极显著正相关, 表明这些农艺性状的双亲均具备较高的一般配合力。对各个性状特殊配合力与相对竞争优势均极显著正相关。比较相对竞争优势值与GCA、SCA间的相关系数, 千粒重、株高GCA与相对竞争优势值相关系数较高, 说明应在双亲千粒重、株高一般配合力均高的前提下, 筛选高的特殊配合力组合; 但单株实粒重、单株有效穗数、主穗二次枝梗数、主穗实粒数、主穗长、主穗一次枝梗数、主穗颖花数SCA与相对竞争优势值相关系数较高, 说明这 7个性状, 亲本一般配合力高低对于筛选高的特殊配合力组合并非必不可少的前提。
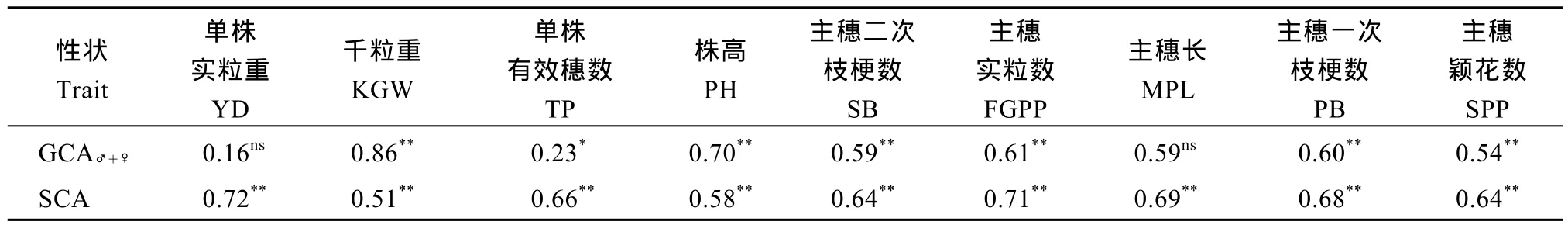
表8 试点1的F1各性状相对竞争优势值(RCH)与GCA及SCA之间的相关系数Table 8 Correlation coefficients of RCH with the sum of parental GCA and SCA for nine traits at Location 1
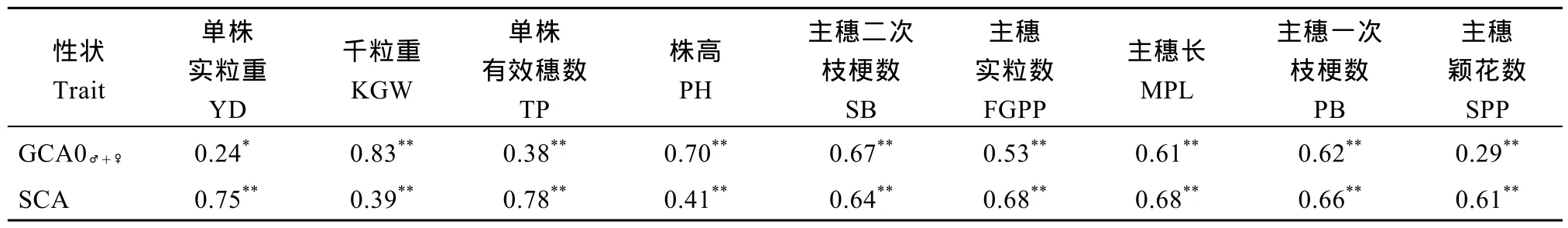
表9 试点2的F1各性状相对竞争优势值(RCH)与GCA和SCA之间的相关系数Table 9 Correlation coefficients of RCH with the sum of parental GCA and SCA for nine traits at Location 2
3 讨论
3.1 杂种优势群体判断
杂种优势, 是指杂交子代在生长活力、育性和种子产量等方面都优于双亲均值的现象[17], 杂交子代的综合表现优于亲本。从植物进化角度考虑, 植物杂交保持了种群的遗传多样性, 增加了自身适应突发环境的能力。自然界中, 植物杂交增加了其适应不同环境的资源库, 这种适应性提高了其杂种后代的综合表现。
当然, 杂种优势形成的基础是亲本间具有一定的遗传差异。胡一波等[18]也认为双亲亲缘关系的远近会影响子代的变异幅度, 亲本间的亲缘关系差异越大, 后代变异幅度越大。遗传背景相似的一类群体可能会形成一个杂种优势群, 不同杂种优势群间的杂交组合容易产生强的杂种优势; 而一个杂种优势群内的杂交组合, 杂种优势较弱。王懿波等[19]对我国玉米主要种质杂种优势群进行了划分, 并提出了我国玉米主要种质的杂交优势利用模式, 广泛用于玉米育种。栽培稻的两个亚种即籼稻、粳稻是水稻两大杂种优势群, 但在其中, 目前并无明显的杂种优势群划分的报道。杂种优势无疑是一个复杂的生物学问题, 针对其遗传基础提出了众多假说, 但各个假说在不同群体的适用性有限。对不同群体,解析的遗传假说均不一致, 反映了杂种优势的随机性。总之, 杂种优势虽然提出已久, 但对它的研究仍有很多的问题亟待解决。
3.2 一般配合力、特殊配合力对筛选高产组合的参照作用
水稻亲本一般配合力、特殊配合力效应的综合评价是杂种优势利用的关键[20-21]。育种实践表明,亲本农艺性状表型优良的杂交组合, 其杂交F1的表型并不一定好; 反之, 亲本表型一般的杂交组合,也可能获得优异的 F1表型。因此, 利用遗传交配设计, 评价亲本一般配合力、特殊配合力效应高低是亲本选育的重要手段。本研究发现, 亲本农艺性状与其一般配合力均为正相关, 其中亲本单株实粒重与一般配合力间相关性不显著。因此, 利用农艺性状表型选育优良组合具有一定的参照价值。
付新民等[8]以明恢63与B5和中国香稻(恢复系)构建的重组自交系与野败型细胞质雄性不育系的杂交组合, 以及余传元等[4]对籼型染色体置换片段在杂交粳稻中的配合力分析结果表明, 在所有测交亲本中, 不同亲本组合及不同性状间的一般配合力和特殊配合力均存在明显差异。余守武等[22]指出选择杂种优势强、农艺性状优良的杂交组合, 恢复系是关键, 而不育系是影响产量性状的主要因素。本研究采用三系野败型杂交水稻的恢复系和微核心种质构成的品种群体, 按照NCII遗传交配设计, 分别与5个不育系测交, 发现亲本的一般配合力水平与特殊配合力之间没有固定的联系。利用不同一般配合力水平的亲本组合, 其后代可能获得高的特殊配合力,也可能出现较低的特殊配合力。与付新民等[8]、余传元等[4]的结论相同。千粒重、株高、主穗实粒数等多个农艺性状的相对竞争优势值与亲本一般配合力之和、特殊配合力均呈极显著正相关。黄殿成等[23]的研究表明, 由于作物遗传理论的复杂性, 农艺性状中优异配合力的标记基因型与品质性状优异配合力的标记基因型存在一定的矛盾, 需要选择适当的标记基因型进行组合, 以同时达到改良亲本农艺性状和品质性状的目的。在后续的研究中, 我们应当结合全基因组重测序、高通量分子生物学检测等方法[24-25], 进一步在分子层面上分析水稻中的配合力与杂种优势间的关系, 更好地解释水稻中的杂种优势现象。
4 结论
亲本一般配合力的高低对特殊配合力并不影响。但包括千粒重在内的多个农艺性状, 其相对竞争优势值与亲本间的一般配合力之和及特殊配合力均极显著正相关。因此, 在育种实践中, 选取一般配合力较高的亲本, 同时兼具较高的特殊配合力是获得高产杂交稻组合的关键。
[1] Sanghera G S, Hussain W. Heterosis and combining ability estimates using Line × Tester analysis to develop rice hybrids for temperate conditions. Notul Sci Biol, 2012, 4: 131
[2] Monforte A J, Tanksley S D. Fine mapping of a quantitative trait locus (QTL) from Lycopersicon hirsutum chromosome 1 affecting fruit characteristics and agronomic traits: breaking linkage among QTLs affecting different traits and dissection of heterosis for yield. Theor Appl Genet, 2000, 100: 471–479
[3] 刘冠明, 李文涛, 曾瑞珍, 张桂权. 水稻亚种间单片段代换系的建立. 中国水稻科学, 2003, 17: 201–204 Liu G M, Li W T, Zeng R Z, Zhang G Q. Development of single segment substitution lines (SSSLs) of subspecies in rice. Chin J Rice Sci, 2003, 17: 201–204 (in Chinese with English abstract)
[4] 余传元, 江玲, 肖应辉, 翟虎渠, 万建民. 籼型染色体置换片段在杂交粳稻中的配合力分析. 作物学报, 2008, 34: 1308–1316 Yu C Y, Jiang L, Xiao Y H, Zhai H Q, Wan J M. Combining ability of yield-component traits for indica chromosome substituted segments in japonica hybrids. Acta Agron Sin, 2008, 34: 1308–1316 (in Chinese with English abstract)
[5] Liu C, Song G Y, Zhou Y H, Qu X F, Guo Z B, Liu Z W, Jiang D M, Yang D C. OsPRR37 and Ghd7 are the major genes for general combining ability of DTH, PH and SPP in rice. Sci Rep, 2015, 5: 12803
[6] Holtrop A. Genetic Architecture for Yield Potential, Density Tolerance, and Yield Stability in Maize (Zea mays L.). PhD Dissertation of University of Guelph, Ontario, Canada, 2016
[7] Huang Z S, Laosuwan P, Machikowa T, Chen Z H. Combining ability for seed yield and other characters in rapeseed. Suranaree J Sci Technol, 2010, 17: 39–47
[8] 付新民, 王岩, 高冠军, 何予卿. 利用水稻重组自交系进行配合力遗传分析. 华中农业大学学报, 2010, 29: 397–402 Fu X M, Wang Y, Gao G J, He Y Q. Combining ability analysis in rice using recombinant inbred lines. J Huazhong Agric Univ, 2010, 29: 397–402 (in Chinese)
[9] 梁康迳. 基因型×环境互作效应对水稻穗部性状杂种优势的影响. 应用生态学报, 1999, 10: 683–688 Liang K J. Interactive effect of genotype and environment on heterosis of panicle traits of rice (Oryza sative L.). Chin J Appl Ecol, 1999, 10: 683–688 (in Chinese with English abstract)
[10] Zhang Y M, Xu S. Mapping quantitative trait loci in F2incorporating phenotypes of F3progeny. Genetics, 2004, 166: 1981–1993
[11] Virmani S S, Aquino R C, Khush G S. Heterosis breeding in rice (Oryza sativa L.). Theor Appl Genet, 1982, 63: 373–380
[12] Gowda M, Kling C, Würschum T, Liu W, Maurer H P, Hahn V, Reif J C. Hybrid breeding in durum wheat: heterosis and combining ability. Crop Sci, 2010, 50: 2224–2230
[13] Green S B, Salkind N J. Using SPSS for Windows and Macintosh: Analyzing and understanding data. Upper Saddle River: Prentice Hall Press, 2010. p 113
[14] 莫惠栋. p×q交配模式的配合力分析(续). 扬州大学学报(农业与生命科学版), 1982, 3(4): 5 Mo H D. The analysis of combining ability in p × q mating pattern (continued). J Yangzhou Univ (Agric & Life Sci Edn), 1982, 3(4): 5 (in Chinese)
[15] 廖伏明, 周坤炉, 盛孝邦, 阳和华, 徐秋生. 籼型三系杂交水稻主要农艺性状配合力研究. 作物学报, 1999, 25: 622–631 Liao F M, Zhou K L, Sheng X B, Yang H H, Xu Q S. Studies on combining ability of major agronomic characters in three-line indica hybrid rice. Acta Agron Sin, 1999, 25: 622–631 (in Chinese with English abstract)
[16] Kannan G R, Ganesh S K. Combining ability analysis for yield, its components and physiological traits in rice under sodicity. Electronic J Plant Breed, 2016, 7: 555–563
[17] Shull G H. What is “heterosis”? Genetics, 1948, 33: 439
[18] 胡一波, 杨修仕, 陆平, 任贵兴. 中国北部藜麦品质性状的多样性和相关性分析. 作物学报, 2017, 43: 464–470 Hu Y B, Yang X S, Lu P, Ren G X. Diversity and correlation of quality traits in quinoa germplasms from North China. Acta Agron Sin, 2017, 43: 464–470 (in Chinese with English abstract)
[19] 王懿波, 王振华. 中国玉米种质基础、杂种优势群划分与杂优模式研究. 玉米科学, 1998, 6(1): 9–13 Wang Y B, Wang Z H. Studies on maize germplasem base, division of heterosis groups and utilizing models of heterosis in China. Maize Sci, 1998, 6(1): 9–13 (in Chinese with English abstract)
[20] Bagheri N, Jelodar N B. Heterosis and combining ability analysis for yield and related-yield traits in hybrid rice. Int J Biol, 2010, 2: 222
[21] Gnanasekaran M, Vivekanandan P, Muthuramu S. Combining ability and heterosis for yield and grain quality in two line rice (Oryza sativa L.) hybrids. Ind J Genet, 2006, 66: 6–9
[22] 余守武, 刘宜柏, 尹建华, 胡标林, 杨平, 揭银泉. 三交水稻的育种研究: III. 三交中晚稻杂种优势的比较研究. 作物学报, 2005, 31: 784–789 Yu S W, Liu Y B, Yi J H, Hu B L, Yang P, Jie Y Q. Study on breeding of three-way hybrid rice (Oryza sativa L.): III. Analysis on the combining ability and heritability of main agronomic traits of three-way hybrid rice in mid-late season cropping. Acta Agron Sin, 2005, 31: 784–789
[23] 黄殿成, 梁奎, 孙程, 江建华, 石明亮, 戴剑, 谢辉, 赵凯铭,阮方松, 马文霞, 洪德林. 杂交粳稻亲本米质性状优异配合力的标记基因型鉴定. 作物学报, 2011, 37: 405–414 Huang D C, Liang K, Sun C, Jiang J H, Shi M L, Dai J, Xie H, Zhao M K, Ruan F S, Ma W X, Hong D L. Identification of marker genotypes associated with elite combining ability for quality traits in parents of hybrid japonica rice (Oryza sativa L.). Acta Agron Sin, 2011, 37: 405–414
[24] Zhang J H, Zhao Y X, Xiao H L, Zheng Y L, Yue B. Genome-wide identification, evolution, and expression analysis of RNA-binding glycine-rich protein family in maize. J Integr Plant Biol, 2014, 56: 1020–1031
[25] Xu Q, Xiong G J, Li P B, He F, Huang Y, Wang K B, Li Z H, Hua J P. Analysis of complete nucleotide sequences of 12 Gossypium chloroplast genomes: origin and evolution of allotetraploids. PLoS One, 2012, 7: e37128
Combining Ability Analysis of Agronomic Trait in Indica × Indica Hybrid Rice
ZHANG Zheng1, ZHANG Xue-Li1, MO Bo-Cheng1, DAI Zhi-Jun1, HU Zhong-Li2, LI Lan-Zhi1,*, and ZHENG Xing-Fei2,*
1Hunan Province Collaborative Innovation Center for the Key Agriculture Pest Control, Hunan Agricultural University, Changsha 410128, China;2State Key Laboratory of Hybrid Rice, Wuhan 430072, China
How to effectively use heterosis is the key of improving yield for rice. In present study, under North Carolina design II, 115 elite indica rice varieties, including three-way wild-abortive hybrid restorer line and mini-core collection, were separately crossed with four two-way restorer lines and one three-way sterile line to generate 575 tested crosses, and combining ability, heritability and correlations of nine agronomic traits were analyzed. Except for tillers per plant and filled grains per panicle, the general combining ability (GCA) variances of most agronomic traits were extremely significant, and except for tillers per plant, the specific combining ability (SCA) variances of other agronomic traits were extremely significant. The SCA of different traits in the same combination or SCA of same trait in different combinations sharing one parent were different, indicating no consistent relationship between SCA and parental GCA. Therefore, selection of high GCA parental line combining with superior SCA combinations is the key to obtain high yield hybrid rice.
Hybrid rice; Three-way wild-abortive type; North Carolina design II; Combining ability; Heterosis
(

附表1 用于测交试验的115份水稻品种及5个不育系材料信息Supplementary table 1 Information of 115 varieties and five sterile lines used in cross experiment
): 2017-01-18; Accepted(接受日期): 2017-04-20; Published online(网络出版日期): 2017-05-08.
10.3724/SP.J.1006.2017.01448
本研究由中国博士后基金项目(2012M511722, 2014T70769), 湖南省自然科学基金项目(2016JJ3070), 湖南省教育厅青年基金项目(2015118)和湖南农业大学作物种质创新与资源利用国家重点实验室培育基地开放课题(16KFXM03)资助。
The study was supported by the China Postdoctoral Science Foundation Funded Project (2012M511722, 2014T70769), the Natural Science Foundation of Hunan Province (2016JJ3070), the Scientific Research Fund of Hunan Provincial Education Department (2015118), and Hunan Provincial Key Laboratory for Germplasm Innovation and Utilization of Crop (16KFXM03).
*通讯作者(Corresponding authors): 李兰芝, E-mail: lancy0829@163.com; 郑兴飞, E-mail: xfzh@whu.edu.cn
联系方式: E-mail: AwithC@gmail.com
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20170508.1006.006.html
