不同水肥运筹对再生季稻根际土壤酶活性及微生物功能多样性的影响
2017-09-25陈鸿飞庞晓敏张仁张志兴徐倩华方长旬李经勇林文雄福建农林大学生命科学学院福建省农业生态过程与安全监控重点实验室福建福州5000福建省南平市建阳区农业局福建建阳5000福建省农业厅种植业技术推广总站福建福州5000重庆市农业科学院重庆060
陈鸿飞庞晓敏张 仁张志兴徐倩华方长旬 李经勇林文雄,*福建农林大学生命科学学院 / 福建省农业生态过程与安全监控重点实验室, 福建福州5000;福建省南平市建阳区农业局, 福建建阳5000;福建省农业厅种植业技术推广总站, 福建福州 5000;重庆市农业科学院, 重庆 060
耕作栽培·生理生化
不同水肥运筹对再生季稻根际土壤酶活性及微生物功能多样性的影响
陈鸿飞1,**庞晓敏1,**张 仁2张志兴1徐倩华3方长旬1李经勇4林文雄1,*
1福建农林大学生命科学学院 / 福建省农业生态过程与安全监控重点实验室, 福建福州350002;2福建省南平市建阳区农业局, 福建建阳353000;3福建省农业厅种植业技术推广总站, 福建福州 350001;4重庆市农业科学院, 重庆 402160
再生季水、肥管理措施是再生稻尤其是机收中低留桩再生稻获得高产稳产的一项关键措施, 明确再生季合理的水肥运筹对提高机收中低留桩再生稻低节位再生芽的萌发成穗具有重要意义。本文以杂交籼稻泸优明占为材料,通过设置再生季分蘖期3个不同水肥处理盆栽试验, 即淹水灌溉不施用促苗肥(S处理)、干湿交替灌溉不施用促苗肥(G处理)和干湿交替灌溉施用促苗肥(GN处理), 研究不同水肥运筹对再生季稻腋芽、产量、根际土壤酶活性及微生物功能多样性的影响。结果表明, 再生季稻不同水肥处理对根际氧化还原电位和根际土壤酶有显著影响, 处理后10 d,与S处理相比, G处理可以显著提高根际土壤的氧化还原电位和根际土壤多酚氧化酶、过氧化物酶、磷酸单酯酶、过氧化氢酶活性, 与G处理相比, GN处理可显著提高根际土壤多酚氧化酶、过氧化物酶、脲酶、蔗糖酶、磷酸单酯酶、过氧化氢酶活性。不同水肥调控下再生季稻根际土壤微生物对碳源的利用程度和碳源代谢的多样性差异显著, 处理后10 d, GN处理和G处理根际微生物对6类碳源的利用程度显著高于S处理, GN处理根际土壤微生物对氨基酸、酚酸、羧酸类碳源的利用程度显著高于G处理。GN处理和G处理的根系伤流量分别比S处理的提高27.27%和14.84%,再生季分蘖数和产量分别比S处理的提高102.50%~111.11%、42.50%~44.44%和91.41%~108.72%、37.93%~40.94%。由此可见, 再生季采用干湿交替灌溉或施用促苗肥均可提高根际土壤酶活性, 促进再生季稻根际土壤微生物对碳源的利用程度和代谢多样性, 从而有利于根际土壤有机质的氧化和腐殖质的形成及增加根际土壤中养分的有效性, 促进再生季新根的形成和腋芽的萌发。采用干湿交替灌溉耦合促苗肥对再生季稻的促控效果是最佳的。
再生稻; 水肥处理; 根际土壤酶活性; 根际微生物
再生稻是一种两收的资源节约型稻作方式, 已成为南方稻区稳粮增收、优化种植结构和提高种粮效益的重要稻作制度。近年来, 随着劳动力成本的不断上升, 机收再生稻逐步成为再生稻的主要种植模式, 采用中低留桩高度成为生产上解决机收再生稻碾压区与非碾压区再生季成熟期不一致和降低机收碾压导致再生季减产的主要措施[1-3], 随着留桩高度的降低, 再生季主体分蘖也由以高节位再生腋芽为主转变为以低节位再生腋芽为主[4], 因此, 促进低节位再生腋芽萌发成穗是提高机收再生稻再生季产量、发挥再生稻稻作制度作用的关键。众多研究结果表明再生腋芽的萌发成穗与再生季稻根系活性有着显著或极显著的相关性[5-6], 再生季稻根系机能对再生腋芽萌发的影响在中低留桩蓄留再生稻时尤为明显; 作物的根系形态和机能除了与品种特性有关外, 还受到环境(水分、营养条件等)的影响[7]。近年来, 许多学者开展了再生季氮肥不同施用量、不同施用比例对机收中低留桩再生稻根系机能和腋芽萌发影响的研究[1,8-10], 但关于再生季水分和氮肥耦合对机收中低留桩再生稻根系机能和腋芽萌发影响的研究却鲜有报道。已有研究表明, 根际是作物–土壤–微生物及其环境条件相互作用的界面, 也是各种养分、水分、有益和有害物质及生物作用于根系或进入根系参与循环的门户[11], 因此, 要阐明再生季水分和氮肥耦合对再生稻根系机能和腋芽萌发的影响, 还必须从根际生态学的角度深入分析。本试验设置再生季不同水肥处理, 探讨对再生稻根系机能及根际土壤酶活性和微生物功能多样性的影响,以期揭示水肥调控再生稻根系机能及腋芽萌发的根际生物学过程, 为机收中低留桩再生稻高产栽培和定向调控提供理论依据。
1 材料与方法
1.1 试验材料与设计
试验于2014—2015年在福建农林大学教学基地(26°10′N, 119°23′E)进行, 供试土壤为沙壤土, 土壤pH 5.65, 含全氮1.98 g kg–1、碱解氮127.90 mg kg–1、有效磷43.10 mg kg–1、速效钾137.51 mg kg–1、有机质23.83 g kg–1。
试验材料为福建省三明市农业科学院选育的杂交稻品种泸优明占, 采用盆栽种植的方式培养, 塑料桶高0.30 m, 顶部直径0.30 m, 底部直径0.23 m,每桶移栽3株水稻, 头季于3月24日播种, 5月4日移栽, 7月18日齐穗, 8月21日收割, 留桩25 cm。头季收割后进行3种水肥处理(S, G, GN), S处理为再生季分蘖期实行淹水灌溉(始终保持水层2~4 cm),不施用促苗肥; G处理为再生季分蘖期干湿交替灌溉(灌 1~2 cm薄水层后自然落干, 直至土壤水势达到-15 kPa时复水至水层 1~2 cm), 不施用促苗肥; GN处理为再生季分蘖期干湿交替灌溉(灌1~2 cm薄水层后自然落干, 直至土壤水势达到-15 kPa时复水至水层 1~2 cm), 施用促苗肥, 在土壤中预埋Watermark Model 200型土壤水分传感器(Irrometer Corporation, 美国)监测水势, 每天下午测水势, 当数值达到限定值时复水, 3个处理再生季氮肥施用见表1, 每个处理40桶, 10月3日再生季齐穗, 11月5日再生季成熟, 3个处理除上述时期水肥管理措施不同外, 头季和再生季其他田间管理措施相同。

表1 再生季稻不同生育时期氮肥施用Table 1 Nitrogen application at different growth stages in ratooning rice
1.2 土壤取样
于头季稻收割后水肥处理前(before irrigation and fertilizer treatments, BIFT)和水肥处理后10 d (10d after irrigation and fertilizer treatments, AIFT)取土壤样品, 每次每个处理随机选取 6株盆栽稻株, 用灭菌过的剪刀剥除离根系较远的土壤, 露出根际土,再尽量取紧靠根面的土壤, 用剪刀轻轻剥掉根际土,混匀后采用五点取样法取土样, 分为 2份分别保存于4℃冰箱和-80℃冰箱备用。
1.3 根系伤流量的测定
参照李合生贴棉吸附法测定伤流量[12], 分别在水肥处理前和处理后10 d测定每个处理6株伤流量,于傍晚7点到次日早上7点收集伤流, 称重, 计算伤流强度。
1.4 再生稻分蘖数观察
头季稻收割后, 选取和稻桩平均数一样或相近的水稻稻桩挂牌, 选取每个处理 9株稻桩挂牌标记,每隔5 d观察再生分蘖数。
1.5 产量和产量构成因子测定
再生季收割时, 选取每处理 6株考察有效穗、穗粒数、结实率和千粒重。
1.6 土壤酶活性测定
参照关松荫等[13]、鲁如坤[14]方法测定土壤酶活性, 采用苯酚钠比色法测定土壤脲酶活性; 采用3,5-二硝基水杨酸比色法测定蔗糖酶活性; 采用高锰酸钾滴定法测定过氧化氢酶活性; 采用对硝基苯磷酸盐法测定磷酸单酯酶活性; 采用邻苯三酚比色法测定多酚氧化酶、过氧化物酶活性。
1.7 土壤微生物群落功能多样性测试
采用BIOLOG ECO微平板法, 称取5 g鲜土于灭菌的三角瓶中, 加入100 mL灭菌生理盐水, 封口, 120 r min–1振荡30 min, 冰上静置2~5 min, 于超净台中轻轻取上清液5 mL移于灭菌的100 mL三角瓶中, 并且加入45 mL无菌水, 摇匀得到1∶100提取液。重复稀释3次, 制得1∶1000的提取液, 立即用于ELISA反应。将BIOLOG ECO平板预热到25℃,用移液枪吸取1∶1000的提取液150 μL于各个孔中,其中A1、A5、A9为150 μL无菌水做空白对照。28℃恒温培养1周, 在0、1、2、3、4、5、6、7 d用ELISA反应平板读数器读取OD590nm的吸光值。1.8 数据分析处理
因 2年间试验处理的趋势基本一致, 本文仅采用2015年数据分析讨论; 微生物群落结构用AWCD作为整体活性的有效指数, AWCD = Σ(C-R)/31, 式中 C为每孔的吸光度, R为对照孔的吸光值; 选择BIOLOG ECO 微平板法培养 96 h的数据计算Simpson优势度指数(J)、Shannon-Wienner多样性指数(H)和Shannon均匀度指数(E), J = 1–Σ Pi2, H= -Σ (Pi·ln Pi), E=H/ln S, 式中Pi为第i孔相对吸光度值与整个平板相对吸光度值总和的比值, S为发生颜色变化的孔数; 采用 Windows 4.5软件作主成分分析(Principal Component Analysis, PCA), R2.14.0软件作聚类分析; 用Microsoft Excel 2013软件进行数据计算和作图, 用DPS 7.05软件进行差异性分析。
2 结果与分析
2.1 不同水肥处理对再生季稻分蘖动态及产量构成的影响
从图 1可知, 再生分蘖增长呈先快后慢趋势,水肥处理后5 d是再生分蘖的快速增长期, 5~15 d再生分蘖进入缓慢增长期, 15 d后基本达到稳定, 但是不同水肥处理再生分蘖的增长速度不同, 水肥处理后5 d, G处理的再生分蘖增长略高于S处理, GN处理的再生分蘖增长明显高于G处理和S处理, 水肥处理后5~15 d, G处理增长速度加快, GN处理和G处理均高于S处理, 至水肥处理后25 d, GN处理的再生分蘖分别比G处理、S处理多46.15%和72.73%。可见, 不同水肥运筹与再生分蘖有着密切联系, 水肥耦合可明显促进休眠芽萌发成再生分蘖。
进一步从表2可知, 与S处理的再生季产量相比, GN处理和G处理有显著的增产作用, GN处理和G处理的再生季产量分别比 S处理的提高 91.41%~108.72%和37.93%~40.94%。在再生季稻产量构成因子中, 3个处理间千粒重和每穗粒数基本相近, 差异未达显著水平, 结实率以S处理最高, 显著高于GN处理和G处理, GN处理和G处理间的结实率差异不显著; 单位面积有效穗数以GN处理最高, G处理次之, GN处理、G处理分别比 S处理高 102.50%~111.11%和 42.50%~44.44%, 3个处理间的差异达显著水平, 表明再生季合理的水分管理和肥料措施确能促进再生腋芽的萌发, 从而通过提高再生季单位面积有效穗实现增产, 但是水肥耦合在促进休眠腋芽萌发的同时, 结实率却有所下降, 这主要是低节位再生分蘖结实率相对较低导致的。
2.2 不同水肥处理对再生季稻根系伤流强度的影响
根系伤流量的变化可以间接地反映水稻的根系活力情况。再生稻不同水肥处理之间的根系伤流量存在显著差异(表3), 与处理前相比, 3个处理的再生季根系伤流量均有所提高, 但提高幅度不同, 再生季水肥处理后10 d, GN处理的根系伤流量为116.72g m–2h–1, G处理的根系伤流量为105.34 g m–2h–1, GN处理比G处理提高10.82%, GN处理和G处理分别比S处理(91.71 g m–2h–1)提高27.27%和14.84%,差异达显著水平, 表明再生季合理的水分管理或肥料措施均对根系特性有显著影响, 且水肥耦合促进再生季稻根系特性的效应较单一措施更为显著。
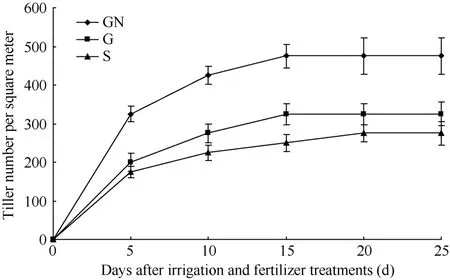
图1 再生稻不同水肥调控下再生分蘖情况Fig. 1 Tiller amounts of ratooning rice under different irrigation and fertilizer treatments
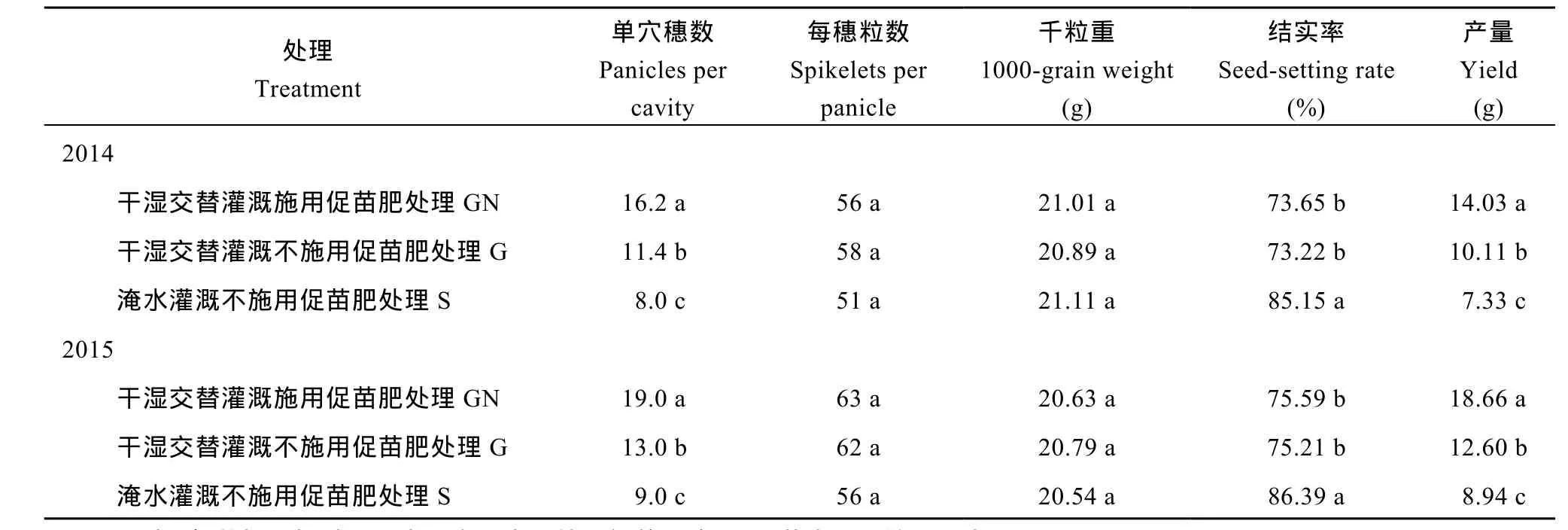
表2 不同水肥处理下再生季水稻盆栽产量及其构成因素Table 2 Yield and its components of ratooning rice in pot experiment under different irrigation and fertilizer treatments
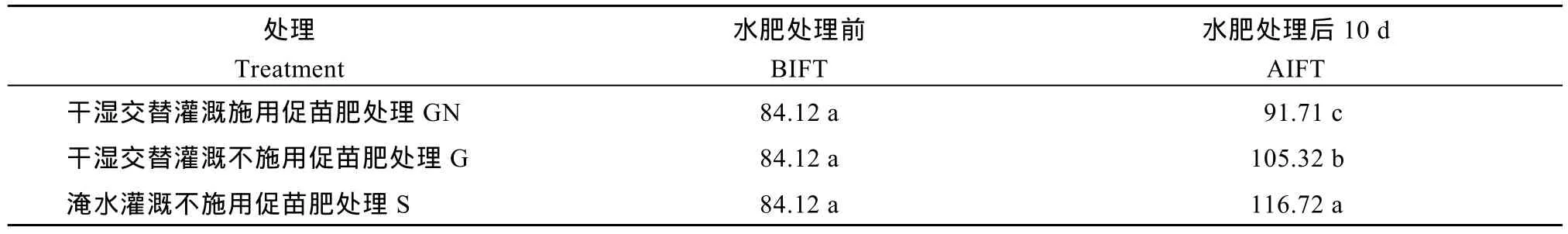
表3 不同水肥处理下再生季稻根系伤流强度的变化Table 3 Bleeding rate of ratooning crop under different irrigation and fertilizer treatments (g m–2h–1)
2.3 不同水肥处理对再生季水稻根际土壤酶活性的影响
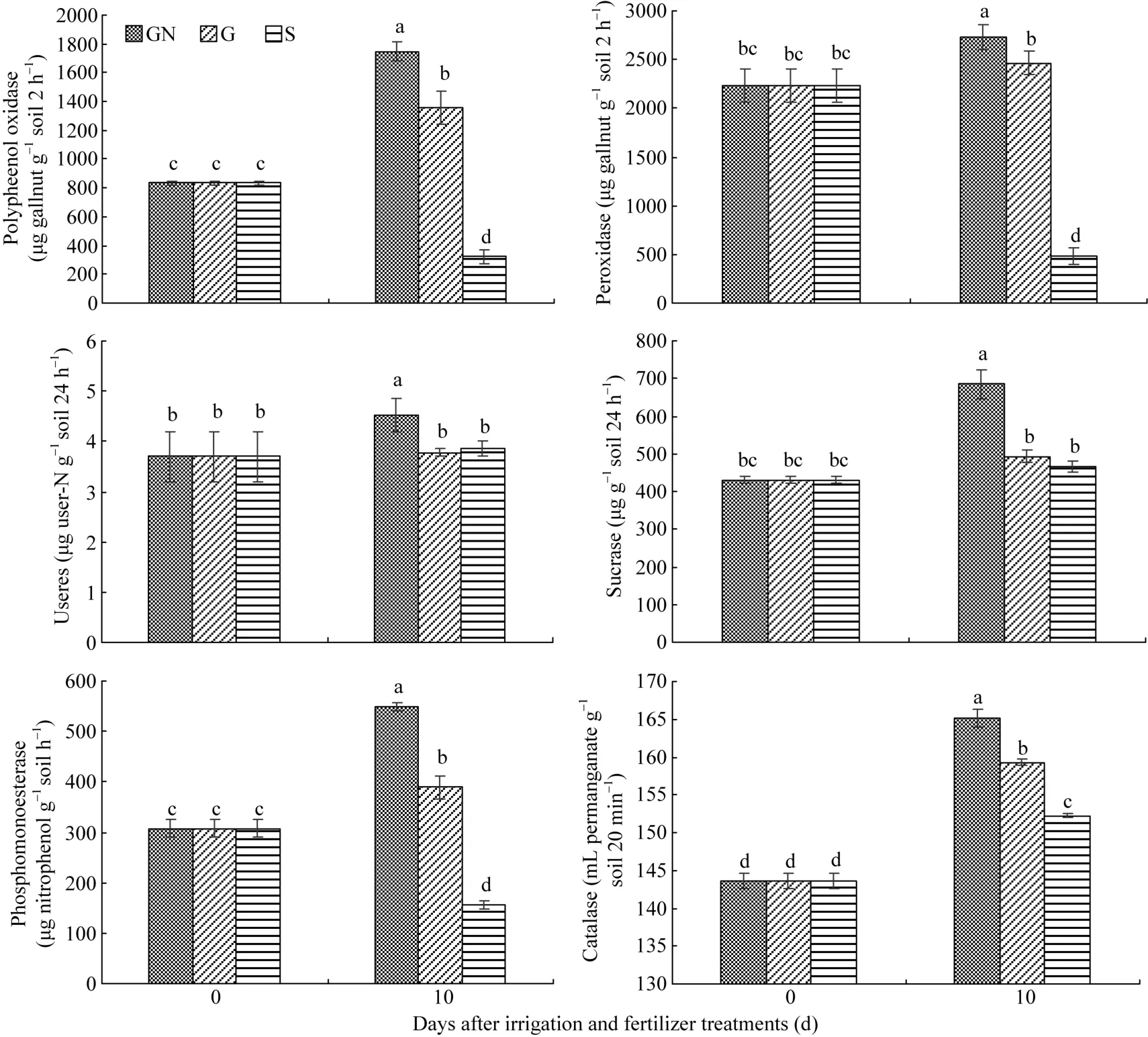
图2 再生季水肥调控对根际土壤酶活性的影响Fig. 2 Effect of different irrigation and fertilizer treatments on rhizospheric soil enzyme activities in ratooning rice
土壤酶参与土壤中各种生物化学过程, 其活性的高低直接反映土壤生物化学过程的强弱[15], 再生季稻不同水肥处理的土壤酶活性差异显著(图2), 与水肥处理前相比, GN处理可以显著提高再生季稻根际土壤的多酚氧化酶、过氧化物酶、脲酶、蔗糖酶、磷酸单酯酶、过氧化氢酶活性, G处理可以显著提高多酚氧化酶、磷酸单酯酶、过氧化氢酶的活性, S处理可以显著提高过氧化氢酶的活性, 而多酚氧化酶、过氧化物酶、磷酸单酯酶活性显著降低; 处理后10 d, 与S处理相比, G处理的土壤多酚氧化酶、过氧化物酶、磷酸单酯酶、过氧化氢酶活性显著提高, 与 G处理相比, GN处理的根际土壤多酚氧化酶、过氧化物酶、脲酶、蔗糖酶、磷酸单酯酶、过氧化氢酶活性显著提高。
2.4 不同水肥处理对土壤Eh和pH值的影响
Eh是反映土壤氧化还原的重要指标, 土壤的Eh适宜范围是200~400 mV。从图3可知, 水肥处理前后, 3个处理的Eh均有所提高, 但提高幅度不同,处理后10 d, S处理的土壤Eh虽有所提高但未达显著水平, GN处理和G处理的土壤Eh显著高于处理前, 与 S处理的差异也达显著水平。在头季稻收割后土壤的 pH值显著提高, 但各个处理之间差异不显著。可见, GN处理和G处理可以提高土壤的通透性和氧含量, 从而显著提高土壤 Eh, 但对土壤 pH值并没有显著影响。
2.5 不同水肥处理再生季水稻根际土壤微生物的Biolog分析
2.5.1 再生季水稻根际土壤微生物对碳源利用程度的动力学特征 从图 4可以发现, 各处理根际微生物对碳源的利用程度随着培养时间的延长而增加,但存在较大差异, 处理后10 d的土壤AWCD值为GN处理>G处理>S处理>处理前, 表明再生季干湿交替灌溉耦合促苗肥处理明显促进了根际土壤微生物群落对碳源的利用程度。
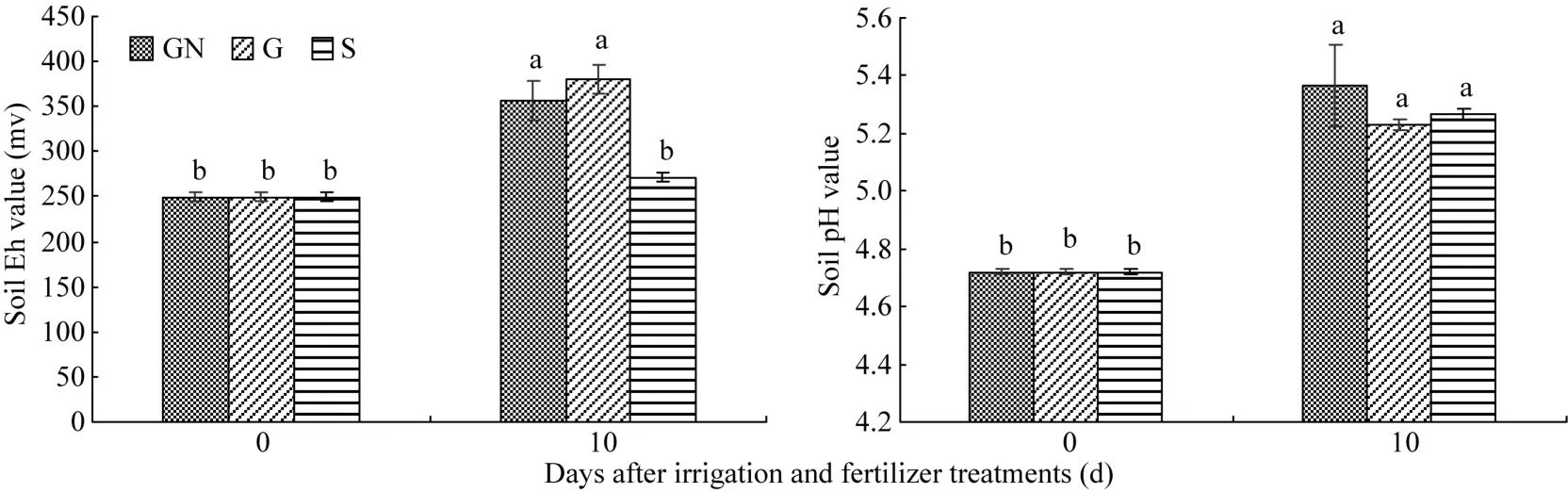
图3 不同水肥调控对再生季水稻土壤Eh、pH值的影响Fig. 3 Effect of different irrigation and fertilizer treatments on rhizospheric soil Eh and pH values in ratooning rice

图4 再生季水肥处理前后根际土壤所有碳源平均颜色(AWCD值)随时间的变化情况Fig. 4 Average well colour development (AWCD) of rhizospheric soil before and after treating
2.5.2 再生季水稻根际土壤微生物群落多样性和均匀度指数分析 土壤微生物群落多样性是群落总体动态变化的反映, 从表4可知, 处理后10 d, GN处理和G处理根际微生物群落多样性和均匀度指数均显著高于S处理和处理前, GN处理和G处理之间的差异也达显著水平, S处理和处理前的差异不显著,可见, 再生季合理的水分管理有助于再生稻根际土壤细菌群落多样性和均匀度的提高, 合理的水分管理耦合施用促苗氮肥则效果更显著。
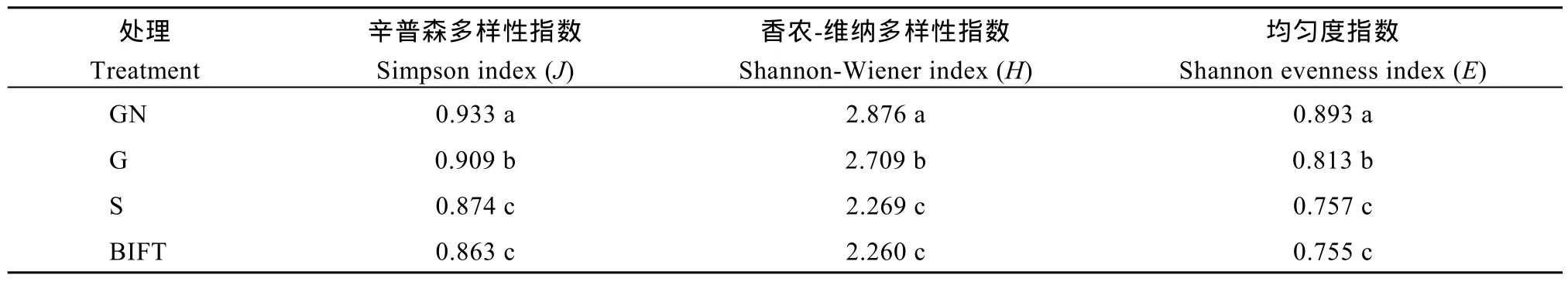
表4 再生季不同水肥运筹方式下根际土壤细菌群落多样性和均匀度指数分析Table 4 Diversity and evenness analysis of rhizospheric soil bacterial communities under different irrigation and fertilizer treatments in ratooning rice
2.5.3 再生季水稻根际土壤微生物对6类碳源的利用程度 对土壤微生物结构的研究, 可以从土壤微生物对碳源利用程度的差异着手。图5表明再生季不同水肥调控下根际土壤微生物对不同类型碳源的利用程度差异显著, 处理后10 d, GN处理和G处理根际微生物对6类碳源的利用程度显著高于S处理, GN处理根际土壤微生物对氨基酸、酚酸、羧酸类碳源的利用程度显著高于 G处理, 而对胺类、聚合物类和碳水化合物类3类碳源的利用程度GN处理和 G处理相近, 差异不显著。由此可见, 再生季合理的水分管理可以显著提高根际微生物对 6类碳源的利用程度, 在此基础上, 适当施用促苗氮肥可以显著提高根际微生物对部分类型碳源的利用程度。
2.5.4 再生季水稻根际土壤微生物对碳源底物利用差异的主成分及聚类分析 从图 6-A可知, 培养96 h的AWCD值能够较好地区分不同水肥处理再生季水稻根际土壤微生物群落对于碳源底物的代谢利用差异。在不同水肥处理后, 主成分1 (PC1)和主成分2 (PC2)分别解释变量方差的49.7%和38.4%, 与主成分1 (PC1)正相关系数最高的单一碳源是Tween 40 (polymers)、D-甘露醇(carbohydrates); 与主成分2(PC2)负相关系数绝对值最高的单一碳源为 α-环式糊精(polymers)和苯乙胺(amines amides)。
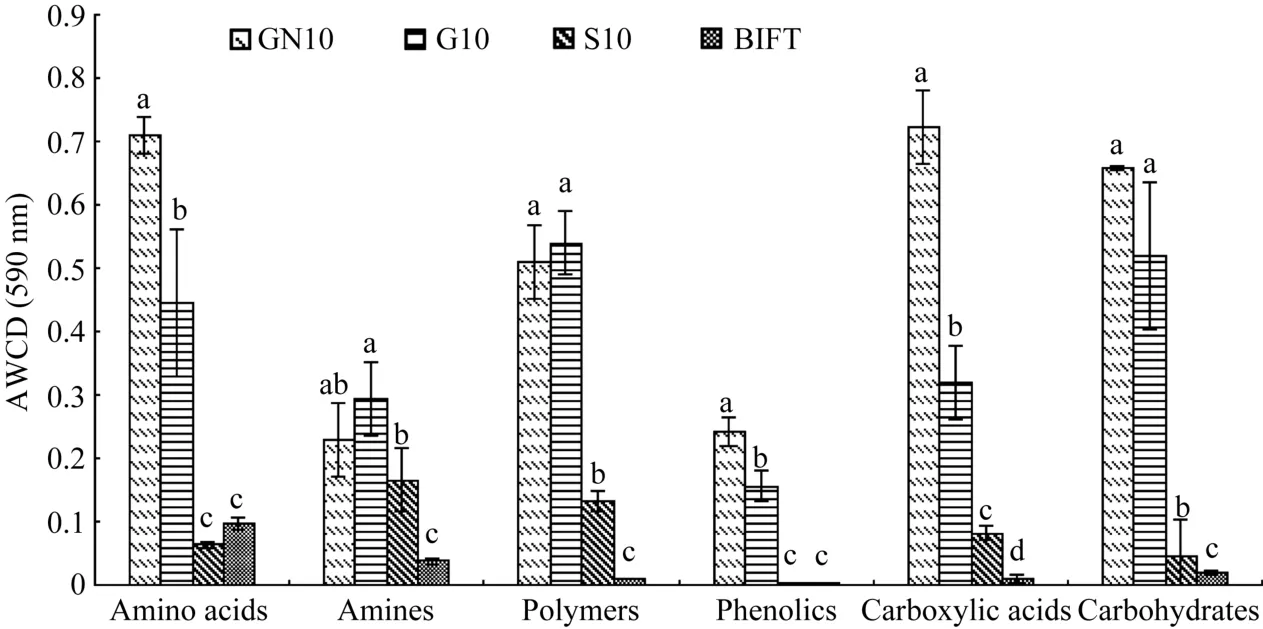
图5 不同水肥处理下根际土壤微生物对不同类型碳源的代谢利用Fig. 5 Effect of different irrigation and fertilizer treatments on utilization of different sources of carbon in rhizospheric soil microbes
图 6-B表明, 不同水肥处理下再生季水稻根际微生物碳源代谢之间存在明显差异。GN和G处理比处理前和 S处理大, 施肥处理比未施肥处理大; G处理和GN处理聚成一类, S处理和处理前聚为一类, 并且各重复样品之间的相关性较好。不同水肥处理再生季水稻的BIOLOG数据分析表明, G处理根际土壤微生物群落的碳源代谢多样性大于S处理和处理前, 在此基础上, 施用促苗肥会进一步提高再生季水稻根际土壤微生物群落碳源代谢的多样性。
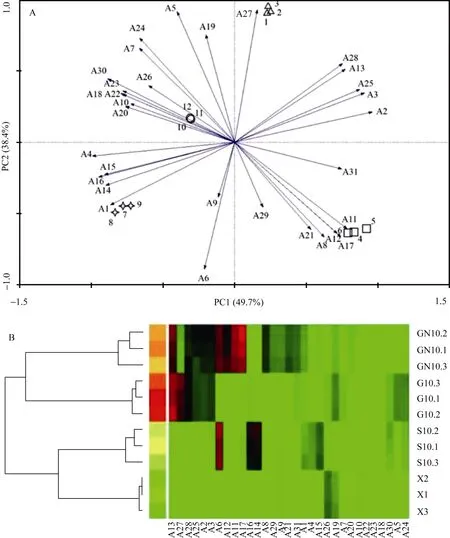
图6 BIOLOG主成分分析图(A)和聚类分析图(B)Fig. 6 Classification of treatments by principal component (A) and cluster (B) analysis with BIOLOG data
3 讨论
再生季稻产量与头季收割后稻桩上休眠腋芽萌发成穗数密切相关[16]。目前, 随着我国经济的发展和农村劳动力的大量转移, 导致季节性缺劳动力,劳动力价格急剧攀升, 机械化轻简再生稻栽培成为再生稻规模化种植发展的必然选择; 机械化轻简再生稻栽培一般采用中低留桩高度来解决机收再生稻碾压区与非碾压区再生季成熟期的不一致和降低机收碾压导致的再生季减产, 因此, 机收中低留桩再生稻低节位腋芽的促发栽培成为机械化轻简再生稻高产稳产栽培的关键; 大量的研究结果表明再生稻休眠芽尤其是低节位腋芽的萌发生长成穗与再生季水稻根系活性有着显著或极显著的相关性[6,17]。再生稻根系由两部分组成, 一是母茎上存活的老根与其在再生季产生的新根, 二是稻桩母茎茎节休眠芽上根原基萌发而成的新根[16], 其中再生新根对机械化收割中低留桩蓄留再生稻再生分蘖的萌发尤为重要,两者间存在着“根芽同伸”关系。作物的根系形态和机能除了与品种特性有关外, 还受到环境(水分、营养条件)的影响[7]。本研究发现再生季单一的干湿交替灌溉或施用促苗肥均能显著提高再生季根系活性,而水肥耦合促进再生季稻根系活性的效应较单一措施则更为显著, 因而, GN处理的再生分蘖分别比G处理、S处理多46.15%和72.73%, 表明再生季合理的水分管理和肥料措施耦合确能更为显著地提高再生季根系活性进而促进再生腋芽的萌发, 提高再生季单位面积有效穗实现增产, 而且增产主要是通过促进低节位腋芽的萌发成穗来实现的。这与俞道标等的研究结果一致[18], 也表明了再生季水、肥运筹通过调控再生季水稻的根系活性进而影响再生腋芽的萌发和再生季稻产量。
环境条件是通过何种途径调控作物的根系构型呢?张福锁等[19]认为栽培调控的实质是调控根际环境, 为作物创造一个良好的根际生态环境从而调控作物-土壤-微生物间的相互作用达到促进作物生长和发育的目的, 根际是作物-土壤-微生物及环境条件相互作用的区域, 也是各种营养物质、水分、有益和有害物质及生物作用于根系或进入根系参与循环的界面。作物-土壤-微生物及其环境之间的相互作用构成了作物的根际生态系统, 它对于土壤的理化性质、根系的形态、生理有着重要的影响[20-21], 根际化学过程包括根际土壤的酸碱反应、氧化还原反应等, 根际土壤酶和根际微生物是根际生态系统中重要组成部分, 在根际生态中起着重要的作用, 它们对于植物根际生态系统的物质循环和能量流动具有重要的影响, 由于根际土壤酶和根际微生物对外界条件尤其是土地管理、耕作制度的影响异常敏感,其变化和群落组成、多样性常常被用作表征土壤质量变化的指标[22]。土壤酶中, 脲酶能分解有机物, 促其水解成氨和二氧化碳; 多酚氧化酶与土壤有机质的形成有关, 是腐殖化的一种媒介; 过氧化物酶是土壤中一种重要的氧化还原酶类, 在有机质氧化和腐殖质形成过程中缺少不了过氧化物酶的作用; 蔗糖酶与土壤肥力的改善和提高关系密切, 它的主要作用是利于土壤中有机质的转化; 磷酸酶对于土壤有机磷化合物的转化有重要的作用, 土壤中有机磷的分解强度取决于其活性高低的变化; 过氧化氢酶活性对于土壤腐殖质化强度大小和有机质积累程度有表征作用。本研究发现再生季稻不同水肥处理的根际氧化还原电位和根际土壤酶活性具有显著差异,处理后10 d, GN处理和G处理可以提高根际土壤的氧化还原电位, 但对根际土壤 pH值的变化没有显著影响, 与S处理相比, G处理对于根际土壤中的多酚氧化酶、过氧化物酶、磷酸单酯酶、过氧化氢酶活性有显著提高作用。Waldrop等[23]的研究结果也指出土壤多酚氧化酶和过氧化物酶的活性对于土壤水分的变化比较敏感, 水分的增加会降低这两种酶的活性, 与G处理相比, GN处理对于六大土壤酶活性都有显著提高作用, 这与周卫等[24]和赵之重[25]的报道相似, 表明再生季干湿交替灌溉有利于提高土壤的通透性和氧含量, 对于根际土壤中有机质的氧化和腐殖质的形成有促进作用, 而水肥耦合措施对于根际土壤有机质的氧化和腐殖质的形成及增加根际土壤中养分有效性的促进作用要显著高于单一措施, 从而为再生季新根的形成和腋芽的萌发提供相应的物质需求保障。
根际土壤微生物是根际土壤酶的重要来源, 同时也直接参与根际的生物过程, 影响根系构型的形成和特性。基于 BIOLOG平板法的根际微生物功能多样性研究表明不同水肥调控下再生季稻根际土壤微生物对碳源的利用程度和碳源代谢的多样性差异显著, 再生季干湿交替水分管理下根际土壤微生物群落对碳源的利用程度和碳源代谢的多样性显著大于淹水处理和处理前。在合理水分管理的基础上, 适当施用促苗肥有利于进一步提高根际微生物对氨基酸、酚酸、羧酸类碳源的利用程度和碳源代谢的多样性。从 31个碳源因子中提取与碳源利用相关的主成分1和主成分2能够解释变量方差总和的88.1%, 在两大成分分离中起主要贡献作用的是聚合物类、碳水化合物类和氨基酸类, 土壤碳水化合物类和氨基酸类是土壤中微生物细胞必需的组成物质和活动的主要能源, 不同水肥调控下根际土壤微生物对根系环境中碳水化合物和氨基酸的利用程度不同, 直接影响根际土壤微生物的数量和种类, 根际土壤微生物数量和种类的不同变化又反过来影响着作物根系的生长发育[26-27]。可见, 再生季合理水分管理和适当促苗肥耦合引起的再生季稻根际土壤微生物对碳源利用程度和碳源代谢多样性的提高, 是导致 GN处理下再生季根际土壤养分有效性和根系活性增强的重要原因。
4 结论
不同水肥调控措施对机收再生稻再生季根际土壤酶活性及微生物功能多样性有显著影响, 与淹水灌溉不施用促苗肥和干湿交替灌溉不施用促苗肥相比, 再生季干湿交替灌溉且施用促苗肥能显著提高根际土壤的氧化还原电位和根际土壤酶活性, 提高根际土壤微生物群落对碳源的利用程度和碳源代谢的多样性, 从而提高机收再生稻再生季的根系机能和产量。因而, 再生季分蘖期采用干湿交替灌溉耦合施用促苗肥是机收中低留桩再生稻高产高效栽培的关键技术之一。
致谢: 本研究得到闽台作物特色种质创制与绿色栽培协同创新中心[闽教科(2015)75号]提供的技术帮助,特此致谢。
[1] 刘正忠. 再生稻头季机收关键技术分析. 农业科技通讯, 2013, (4): 118–120 Liu Z Z. Analysis on the machinery-harvested technique of ratoon rice at the first cropping season. Bull Agric Sci & Technol, 2013, (4): 118–120 (in Chinese with English abstract)
[2] 刘秀斌, 刘正忠, 罗美玉. 再生稻头季机收留桩高度试验初报.福建农业科技, 2014, (5): 13–15 Liu X B, Liu Z Z, Luo M Y. Primary trial report on machineryharvested stubble height of ratoon rice at the first cropping season. Fujian Agric Sci & Technol, 2014, (5): 13–15 (in Chinese with English abstract)
[3] 钱太平, 梅少华, 张键, 喻均吉, 翟中兵, 方锡文, 陈建军, 张丛德, 魏坦雄. 再生稻不同留桩高度和收割方式的产量及其构成因素分析. 湖北农业科学, 2015, 54(1): 14–17 Qian T P, Mei S H, Zhang J, Yu J J, Zhai Z B, Fang X W, Chen J J, Zhang C D, Wei T X. Yield and yield components of ratoon rice with different stubble height and harvest methods. Hubei Agric Sci, 2015, 54(1): 14–17 (in Chinese with English abstract)
[4] 俞道标, 赵雅静, 黄顽春, 李小萍, 姜照伟, 陈双龙. 低桩机割再生稻生育特性和氮肥施用技术研究. 福建农业学报, 2012, 27: 485–490 Yu D B, Zhao Y J, Huang W C, Li X P, Jiang Z W, Chen S L. Growth characteristics and nitrogen application technique of ratoon rice with machine harvest in low cutting. Fujian J Agric Sci, 2012, 27: 485–490 (in Chinese with English abstract)
[5] 张林, 郭晓艺, 刘茂, 熊洪, 朱永川, 周兴兵, 徐富贤. 杂交中稻再生芽生长的影响因素研究. 西南农业学报, 2010, 23(2): 21–26 Zhang L, Guo X Y, Liu M, Xiong H, Zhu Y C, Zhou X B, Xu F X. Factors for promoting ratooning bud growth of mid-season hybrid rice. Southwest China J Agric Sci, 2010, 23(2): 21–26 (in Chinese with English abstract)
[6] 郑景生. 再生稻高产栽培特性与相关性状的基因定位研究.福建农林大学博士学位论文, 福建福州, 2004 Zheng J S. The Characteristics of High-yielding Cultivation and Gene Mapping for Related Traits of Ratoon Rice (Oryza sativa L.). PhD Dissertation of Fujian Agriculture and Forestry University, Fuzhou, China, 2004 (in Chinese with English abstract)
[7] 潘晓华, 王永瑞, 傅家瑞. 水稻根系生长生理的研究进展. 植物学通报, 1996, 13(2): 13–20 Pan X H, Wang Y R, Fu J R. Advance in the study on the growth-physiology in rice of root system (Oryza sativa). Chin Bull Bot, 1996, 13(2): 13–20 (in Chinese with English abstract)
[8] 俞道标, 张燕, 赵雅静, 李小萍, 姜照伟, 陈双龙. 低桩机割再生稻氮肥施用技术研究. 中国农学通报, 2013, 29(36): 210–214 Yu D B, Zhang Y, Zhao Y J, Li X P, Jiang Z W, Chen S L. Nitrogen application technique of ratoon rice with machine harvest in low cutting. Chin Agric Sci Bull, 2013, 29(36): 210–214 (in Chinese with English abstract)
[9] 黄水明. 机收低桩再生稻栽培技术研究. 福建农林大学硕士学位论文, 福建福州, 2010 Huang S M. Study on Low Pile Rationing Rice Cultivation Techniques. MS Thesis of Fujian Agriculture and Forestry University, Fuzhou, China, 2010 (in Chinese with English abstract)
[10] 刘勇涛. 低桩再生稻促芽肥施用方法及低桩机割对再生季产量的影响. 福建稻麦科技, 2013, 31(2): 7–10 Liu Y T. Study on application method of bud fertilizer and influence on yield for II you 5928 as low stubble and machine harvesting. Fujian Sci Technol Rice & Wheat, 2013, 31(2): 7–10 (in Chinese with English abstract)
[11] 李春俭, 马玮, 张福锁. 根际对话及其对植物生长的影响. 植物营养与肥料学报, 2008, 14: 178–183 Li C J, Ma W, Zhang F S. Rhizosphere talk and its impacts on plant growth. Plant Nutr Fert Sci, 2008, 14: 178–183 (in Chinese with English abstract)
[12] 李合生. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 2000. pp 164–261 Li H S. Experiment Principle and Technique on Plant Physiology and Biochemistry. Beijing: Higher Education Press, 2000. pp 164–261 (in Chinese)
[13] 关松荫, 张德生, 张志明. 土壤酶及其研究法. 北京: 农业出版社, 1986. pp 274–325 Guan S Y, Zhang D S, Zhang Z M. Soil Enzymes and Its Research Methods. Beijing: Agriculture Press, 1986. pp 274–325 (in Chinese)
[14] 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社. 2000. pp 252–254 Lu R K. Analysis Method of Soil Agricultural Chemistry. Beijing: China Agricultural Science and Technology Press, 2000. pp 252–254 (in Chinese)
[15] 万忠梅, 吴景贵. 土壤酶活性影响因子研究进展. 西北农林科技大学学报(自然科学版), 2005, 33(6): 87–92 Wan Z M, Wu J G. Study progress on factors affecting soil enzyme activity. J Northwest Sci-Tech Univ Agric & For (Nat Sci Edn), 2005, 33(6): 87–92 (in Chinese with English abstract)
[16] 林文雄, 陈鸿飞, 张志兴, 徐倩华, 屠乃美, 方长旬, 任万军.再生稻产量形成的生理生态特性与关键栽培技术的研究与展望. 中国生态农业学报, 2015, 23(4): 392–401 Lin W X, Chen H F, Zhang Z X, Xu Q H, Tu N M, Fang C X, Ren W J. Research and prospect on physio-ecological properties of ratoon rice yield formation and its key cultivation technology. Chin J Eco-Agric, 2015, 23(4): 392–401 (in Chinese with Englishabstract)
[17] 林文, 李义珍, 姜照伟, 郑景生. 再生稻根系形态和机能的品种间差异及与产量的关联性. 福建农业学报, 2001, (1): 1–4 Lin W, Li Y Z, Jiang Z W, Zheng J S. Morphologic and functional difference of root systems among ratooning rice varieties and its correlation with yield. Fujian J Agric Sci, 2001, (1): 1–4 (in Chinese with English abstract)
[18] 俞道标, 赵雅静, 黄顽春, 李小萍, 姜照伟, 陈双龙. 低桩机割再生稻生育特性和氮肥施用技术研究. 福建农业学报, 2012, 27: 485–490 Yu D B, Zhao Y J, Huang W C, Li X P, Jiang Z W, Chen S L. Growth characteristics and nitrogen application technique of ratoon rice with machine harvest in low cutting. Fujian J Agric Sci, 2012, 27: 485–490 (in Chinese with English abstract)
[19] 张福锁, 王激清, 张卫峰, 崔振岭, 马文奇, 陈新平, 江荣风.中国主要粮食作物肥料利用率现状与提高途径. 土壤学报, 2008, 45: 915–924 Zhang F S, Wang J Q, Zhang W F, Cui Z L, Ma W Q, Chen X P, Jiang R F. Nutrient use efficiencies of major cereal crops in China and measures for improvement. Acta Pedol Sin, 2008, 45: 915–924 (in Chinese with English abstract)
[20] 沈建凯, 黄璜, 傅志强, 张亚, 陈灿, 金小马, 廖晓兰, 龙攀, 高文娟, 谢小婷. 规模化稻鸭生态种养对稻田杂草群落组成及物种多样性的影响. 中国生态农业学报, 2010, 18(1): 123–128 Shen J K, Huang H, Fu Z Q, Zhang Y, Chen C, Jin X M, Liao X L, Long P, Gao W J, Xie X T. Effect of large-scale rice-duck eco-farming on the composition and diversity of weed community in paddy fields. Chin J Eco-Agric, 2010, 18(1): 123–128 (in Chinese with English abstract)
[21] 裴鹏刚, 张均华, 朱练峰, 禹盛苗, 金千瑜. 根际氧浓度调控水稻根系形态和生理特性研究进展. 中国稻米, 2013, 19(2): 6–8 Pei P G, Zhang J H, Zhu L F, Yu S M, Jin Q Y. Research progress on effects of rhizosphere dissolved oxygen on root morphology and physiological characteristics in rice. China Rice, 2013, 19(2): 6–8 (in Chinese)
[22] 胡婵娟, 郭雷, 刘国华. 黄土丘陵沟壑区不同植被恢复格局下土壤微生物群落结构. 生态学报, 2014, 34: 2986–2995 Hu C J, Guo L, Liu G H. Soil microbial community structure under different vegetation restoration patterns in the loess hilly area. Acta Ecol Sin, 2014, 34: 2986–2995 (in Chinese with English abstract)
[23] Waldrop M P, Zak D R, Sinsabaugh R L, Gallo M, Lauber C. Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity. Ecol Appl, 2004, 14: 1172–1177
[24] 周卫, 刘经荣, 张德远. 水稻土有机-无机肥料配合施用的效应(之四). 江西农业大学学报, 1990, (2): 24–31 Zhou W, Liu J R, Zhang D Y. The effect of mixed application of organic and inorganic fertilizers in paddy soil (the fourth report). Acta Agric Univ Jiangxiensis, 1990, (2): 24–31 (in Chinese with English abstract)
[25] 赵之重. 土壤酶与土壤肥力关系的研究. 青海大学学报(自然科学版), 1998, (3): 24–29 Zhao Z Z. The Relationships between soil enzyme and soil fertility. J Qinghai Univ, 1998, (3): 24–29 (in Chinese with English abstract)
[26] 高子勤, 张淑香. 连作障碍与根际微生态研究: I. 根系分泌物及其生态效应. 应用生态学报, 1998, 9: 549–554 Gao Z Q, Zhang S X. Continuous cropping obstacle and rhizospheric microecology: I. Root exudates and their ecological effects. Chin J Appl Ecol, 1998, 9: 549–554 (in Chinese with English abstract)
[27] 张淑香, 高子勤. 连作障碍与根际微生态研究: II. 根系分泌物与酚酸物质. 应用生态学报, 2000, 11: 153–157 Zhang S X, Gao Z Q. Continuous cropping obstacle and rhizospheric microecology: II. Root exudates and phenolic acids. Chin J Appl Ecol, 2000, 11: 153–157 (in Chinese with English abstract)
Effects of Different Irrigation and Fertilizer Application Regimes on Soil Enzyme Activities and Microbial Functional Diversity in Rhizosphere of Ratooning Rice
CHEN Hong-Fei1,**, PANG Xiao-Min1,**, ZHANG Ren2, ZHANG Zhi-Xing1, XU Qian-Hua3, FANG Chang-Xun1, LI Jing-Yong4, and LIN Wen-Xiong1,*
1College of Life Sciences, Fujian Agriculture and Forestry University / Fujian Provincial Key Laboratory of Agroecological Processing and Safety Monitoring, Fujian Agriculture and Forestry University, Fuzhou 350002, China;2Agriculture Bureau of Jianyang District, Nanping City, Fujian Province, Jianyang 353000, China;3Planting Technique Extension Station of Fujian Province Agriculture Department, Fuzhou 350001, China;4Chongqing Academy of Agricultural Sciences, Chongqing 402160, China
Irrigation and fertilizer application regime is one of key measures for high and stable yield of ratooning rice, especiallyfor middle or low stubble ratooning rice by machine-harvest. Ascertaining rational management of water and fertilization is important for improving the axillary bud germination rate of lower nodes in middle or low stubble ratooning rice by machine-harvest. With Luyoumingzhan as a test material, the effects of different irrigation and fertilizer application regimes on ratooning tillers, soil enzyme activity and rhizosphere microbial functional diversity of ratooning rice were studied by setting up three treatments: (1) dry-wet alternate irrigation and nitrogen application for ratooning bud development (GN); (2) dry-wet alternate irrigation and no nitrogen application for ratooning bud development (G); (3) flooding irrigation and no nitrogen application for ratooning bud development (S). The different irrigation and fertilization treatments had a significant impact on the rhizosphere Eh potential and rhizosphere soil enzymes. On the 10th day after treatment, the rhizosphere soil Eh levels and the activities of rhizosphere soil polyphenol oxidase, peroxidase, phosphate monoester enzyme and hydrogen peroxide enzyme were significantly improved in G than in S. The activities of rhizosphere soil polyphenol oxidase, peroxidase, urease, invertase, phosphate monoester enzyme and hydrogen peroxide enzyme were significantly improved in GN than in G. The rhizosphere soil microbe differed significantly in use of single carbon sources and diversity of carbon metabolism under different treatments. On the 10th day after treatment, the uses of single carbon sources and diversity of carbon metabolism in GN and G were significantly greater than those in S. The abilities to utilize amino acids, phenolic acids, carboxylic acid in GN were higher than those in G. Compared with S, bleeding rate of GN and G was increased by 27.27% and 14.84%, ratooning rice tiller and yield were increased by 102.50%–111.11%, 42.50%–44.44% and 91.41%–108.72%, 37.93%–40.94%, respectively. All these results indicated that dry-wet alternate irrigation or nitrogen application for ratooning bud development could improve rhizosphere soil enzyme activities and promote the uses of single carbon sources and diversity of carbon metabolism, which is conducive to increasing the effectiveness of nutrient, the formation of new roots and the axillary buds germination. The effects of dry-wet alternate irrigation coupling with fertilizer are the best for ratooning rice growth.
Ratoon rice; Irrigation and fertilization treatments; Rhizosphere soil enzymes; Rhizosphere microorganisms
(
): 2017-03-06; Accepted(接受日期): 2017-05-10; Published online(网络出版日期): 2017-06-09.
10.3724/SP.J.1006.2017.01507
本研究由国家重点研发计划项目(2016YFD0300508), 福建省高校青年自然基金重点项目(JZ160435)和福建农林大学科技创新专项基金项目(CXZX2016076, CXZX2016077)资助。
This study was supported by the National Key Research and Development Program of China (2016YFD0300508), the Natural Foundation of Fujian Higher Education Institutions for Young Scientists (Key Project) (JZ160435), and the Sci-tech Innovation Fund Project of Fujian Agriculture and Forestry University (CXZX2016076, CXZX2016077).
*通讯作者(Corresponding author): 林文雄, E-mail: lwx@fafu.edu.cn**同等贡献(Contributed equally to this work)
联系方式: 陈鸿飞, E-mail: hongfeichen2006@163.com
URL: http://kns.cnki.net/kcms/detail/11.1809.S.20170609.1745.002.html
