软枣猕猴桃栽培品种DNA 指纹图谱的构建及遗传多样性分析
2017-09-21王丹丹张彦文
王丹丹,张彦文,2
(1.辽东学院农学院,辽宁 丹东118003; 2.东北师范大学草地研究所,吉林 长春 130024)
软枣猕猴桃栽培品种DNA 指纹图谱的构建及遗传多样性分析
王丹丹1,张彦文1,2
(1.辽东学院农学院,辽宁 丹东118003; 2.东北师范大学草地研究所,吉林 长春 130024)
为了对软枣猕猴桃栽培品种进行分子鉴别及分析不同品种间的亲缘关系,采用对叶片直接进行PCR扩增的方法,利用EST-SSR分子标记技术构建了常见的42个软枣猕猴桃品种的数字指纹图谱和模式指纹图谱数据库(cluster project),并进行了遗传多样性分析.结果表明:利用21对软枣猕猴桃核心引物扩增所有品种的基因组,共检测到132个等位位点,包括122个多态性位点,多态性比率为92.4%.每对引物扩增出的等位位点数为3~8个,平均每对引物扩增6.28个基因型,扩增条带大小100~300 bp,多态信息含量(polymorphism information content,CPIC)为0.721,基因流为1.454,平均杂合率为0.691.其中,7对引物,即ACAR2,ACAR13,ACAR53,ACAR75,ACAR92,ACAR112和ACAR120可作为指纹图谱构建的首选引物,利用其中的ACAR2,ACAR13,ACAR53和ACAR75以引物组合方式构建指纹图谱可区分42个品种.聚类分析表明在遗传距离0.64处42个品种被分为一类,在遗传距离0.81处分为4类.聚类结果与品种的特性及来源具有较高的一致性.
软枣猕猴桃;EST-SSR标记;指纹图谱;遗传多样性;聚类分析
品种真实性和纯度鉴定的方法主要有形态学鉴定法、生物化学法及分子标记鉴定法.由于受材料本身以及环境条件的影响,前两种方法已很少作为品种真实性和纯度鉴定的单一的评判标准.[12-15]而分子标记鉴定法 (expressed sequence tag-simple sequence repeats,EST-SSR)以其分子标记为共显性、扩增的谱带少、易识别、统计方便[15-17],以及EST来自DNA编码区其序列保守性相对较高等优点,已成为品种真实性和纯度鉴定的有效方法[18-19].我国已建立了多种植物的指纹图谱以鉴定种质的真实性和纯度,如《水稻品种鉴定DNA指纹法》NY/T1433-2007、《玉米品种鉴定DNA指纹法》NY/T1432-2007等[20-24],而利用EST-SSR分子标记构建软枣猕猴桃DNA指纹谱图的研究尚未见报道[25-27].本研究采用EST-SSR分子标记技术建立了软枣猕猴桃DNA指纹谱图,为其种质资源的真实性和纯度鉴定奠定了基础.同时,在研究过程中,采用对叶片直接进行PCR扩增的方法,简化了实验程序、优化了PCR体系.
1 材料与方法
1.1 实验材料
本实验采用的42个软枣猕猴桃栽培品种分别来自5个国家,具有不同的遗传背景.种子由国内主要软枣猕猴桃研究所或品种引进机构提供.春季或夏季采集生长旺盛的健康、幼嫩叶片于硅胶中干燥保存,以备提取基因组DNA.[28]实验分析工作在辽东学院农学院进行.
1.2 叶片的碱处理
将叶片剪碎,大小不超过2 mm2,否则不宜进行后续的PCR扩增.将叶片放入含有40 μL 0.25 mol/L NaOH的Eppendorf管中,沸水煮30 s;加入40 μL 0.25 mol/L HCl和20 μL 0.5 mol/L Tris-HCl(pH=8.0,含有0.25% 的Nonidet-P-40),再次置于沸水中2 min,随后取出叶片,直接进行PCR扩增.
1.3 EST-SSR PCR扩增
在50 μL PCR反应体系中加入1片经碱处理的叶片.反应液组成为:dNTPs (2.5 mmol/L ) 4 μL,引物(10 μmol/L)各1.8 μL,Taq DNA聚合酶(5 U/μL)0.25 μL,MgCl2(1.5 mmol/L) 3 μL,10×缓冲液5.0 μL,加灭菌的ddH2O至总体积50 μL.PCR反应程序:94℃预变性10 min;94℃变性30 s,58℃~62℃退火30 s,72℃延伸45 s,35个循环;72℃延伸10 min.产物4℃保存[29].
扩增产物采用8%非变性聚丙烯酰胺凝胶电泳分离、银染显色和图谱分析[30].实验所用的21对引物是作者从先前196对EST-SSR引物中筛选出来的[31],具有多态性高、稳定性好等特点(见表1).引物由生工生物工程(上海)股份有限公司合成.
1.4 数据处理
(4) 第四层时间服务:连接信号子网的网关A和网关B作为NTP服务的服务器端来提供时间服务,信号子网内的所有主机(包括ATC车载设备、ATC其它轨旁设备和DCS设备)作为NTP服务的客户端来同步两个网关的时间。
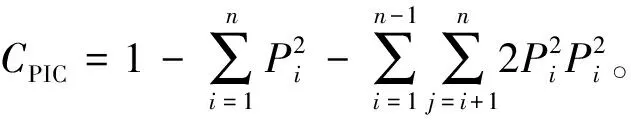
2 结果与分析
2.1 EST-SSR引物扩增
利用21对引物对42份软枣猕猴桃进行扩增,部分引物扩增结果见图1.实验共获得132个等位位点,包括122个多态性位点,多态性比率为92.4%.每对引物扩增出的等位位点数为3~8个,每对引物平均扩增6.28个基因型,扩增的条带大小介于100~300 bp.多态信息含量(CPIC)是表示微卫星DNA变异程度高低的一个指标,反映微卫星DNA多态性高低,其大小取决于检测的等位基因的数目和该基因的基因频率分布,一般来说,CPIC>0.5,多态性较高;0.5≥CPIC≥0.25多态性适中;CPIC<0.25,多态性较低.而本研究21对EST-SSR引物的平均CPIC值为0.721(0.503~0.899,见表1),表现出了较高的多态性;基因流(Nm)平均值为1.454(0.809~3.100,见表1),FST平均值为0.158(0.077~0.236,P<0.05,见表1).
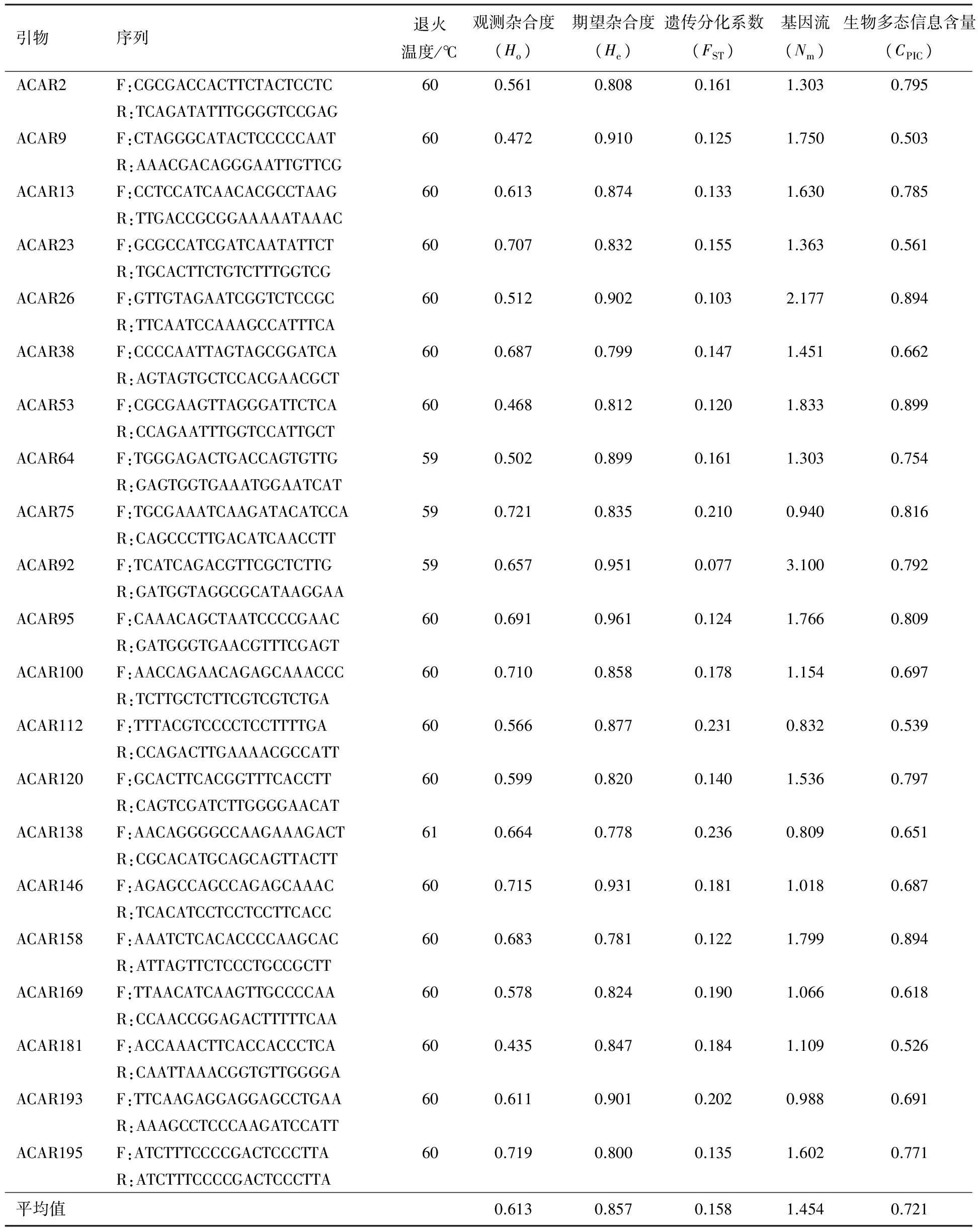
表1 EST-SSR引物在42份供试材料中的扩增情况
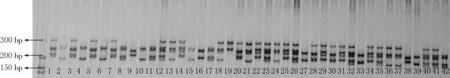
图1 引物ACAR92对软枣猕猴桃扩增的电泳图谱
2.2 EST-SSR指纹图谱的构建
由于同一引物在不同品种中扩增的条带具有差异,同一电泳位置上有带记为1,无带记为0.将数据输入Excel中,按照条带从小到大的顺序排列,同一竖排中进行数据排序,将0和1 分开以便于比对,再将此排中0或者1最少的一组单独提出,随即删除第一排,而后再对第二排进行同样的排序、比对,以此类推,便能逐步比对出与所有品种完全不同的0,1指纹及其所代表的品种,亦能找到指纹完全相同的品种.21对引物中有7对引物具有区别不同品种的能力,即ACAR2,ACAR13,ACAR53,ACAR75,ACAR92,ACAR112和ACAR120(见表2).ACAR2区别品种最多,为3,8,9,14,16,20,23,26,30,33,34,37和40号品种,其中4,15,27,29,32和35号品种带型一致;17,25,31,41和42号品种带型一致;5,11,22,28和39号品种带型一致;10,13,19,21和24号品种带型一致;18和36号品种带型一致.

表2 基于7对EST-SSR引物区分42份品种的能力
根据7对引物的多态性,采用特殊引物法可鉴定32份软枣猕猴桃品种,其余10种不能鉴定,即2,5,10,15,24,28,32,35,38和41号品种.因此,本研究采用引物组合法只需4对EST-SSR引物便可完全区分42份软枣猕猴桃品种,指纹图谱见表3.采用ClusterProject软件对构建的指纹图谱数据进行分析形成指纹模式图谱(见图2),由此,应用两种EST-SSR指纹分析方式共同建立了软枣猕猴桃的指纹图谱数据库.
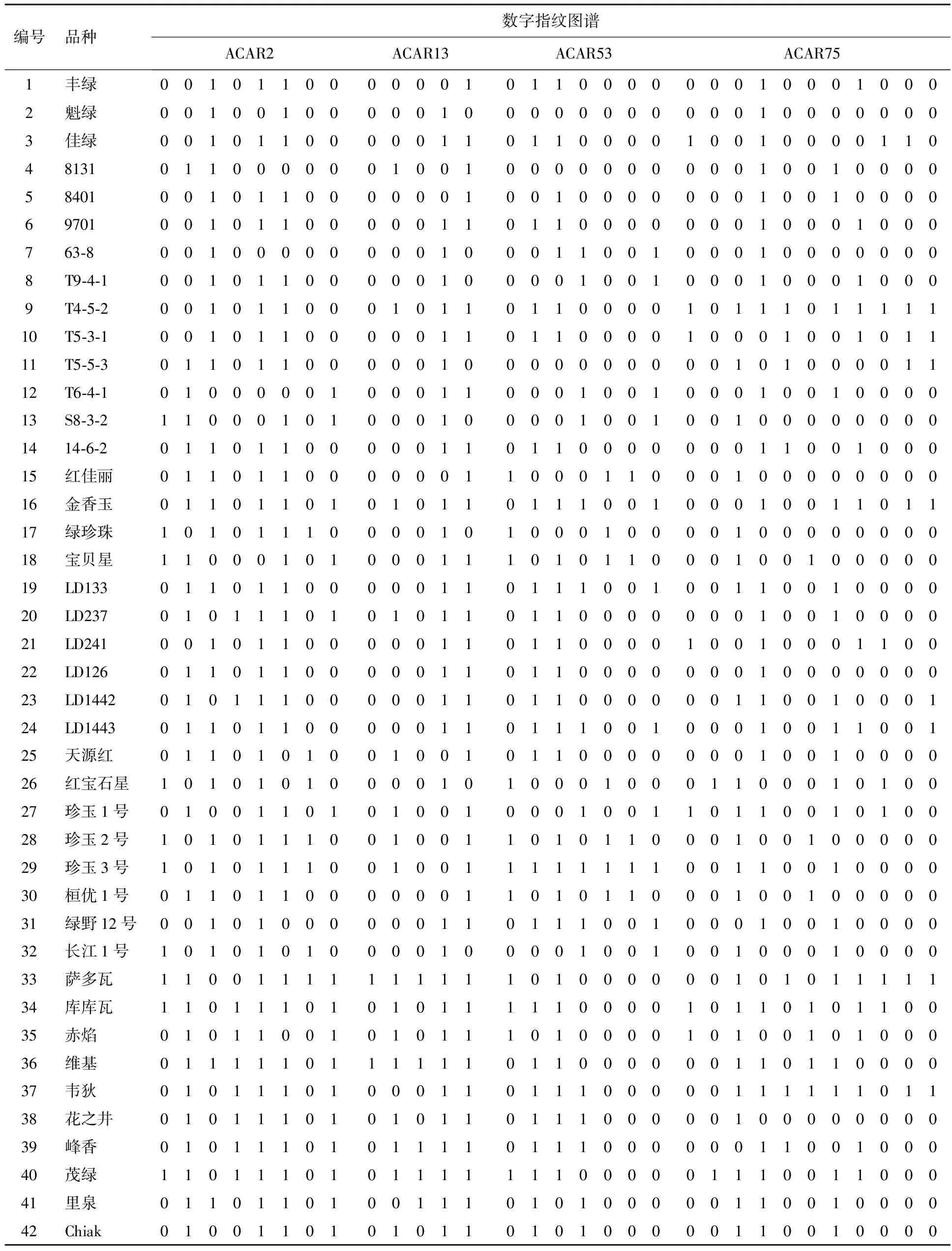
表3 软枣猕猴桃品种的DNA数字指纹图谱

图242份软枣猕猴桃品种的EST-SSR指纹模式图谱
2.3 聚类分析
采用UPGMA进行聚类分析,构建了42份软枣猕猴桃品种的遗传关系树状图(见图3).42份软枣猕猴桃品种被划分为4个类群(见表4),聚类结果与品种的特性、来源具有较高的一致性,即来源于同一地区(或国别)和形态特性相似的品种聚在一个类群.第一类群有28个品种,包括日本品种茂绿,该类群中大部分为绿果皮、绿果肉的品种;第二类群有10个品种,包括新西兰的萨多瓦、乌克兰的赤焰、库库瓦、维基、韦狄4个品种以及国内的LD1442、红佳丽、天源红、红宝石星、LD1443的5个品种,表明它们的遗传关系极为相近,这些品种基本上为红果皮或红肉系列.国外品种和国内品种聚在一起,表明很多国内品种具有国外品种的血统,是授粉系和优良不育系反复使用的结果.第3类群有3个品种,即花之井、峰香和里泉,均来自于日本.第4类群仅有1个品种Chiak,与其他41个品种之间的平均遗传距离为0.451,最大遗传距离为0.871,为所有品种间遗传距离最大的,表明该品种的遗传基础与其他品种差别较大.

图3 42份软枣猕猴桃品种遗传关系树状图

类群品种个数品种名称Ⅰ28金香玉、绿珍珠、LD133、LD237、LD241、LD126、恒优1号、珍玉1号、珍玉2号、珍玉3号、魁绿、8131、63⁃8、T4⁃5⁃2、T5⁃5⁃3、长江1号、茂绿、佳绿、9701、绿野12号、宝贝星、丰绿、8401、T9⁃4⁃2、T5⁃3⁃1、T6⁃4⁃1、14⁃6⁃2、S8⁃3⁃2Ⅱ10LD1442、红佳丽、天源红、红宝石星、LD1443、韦狄、萨多瓦、库库瓦、赤焰、维基Ⅲ3花之井、峰香、里泉Ⅳ1Chiak
3 讨论
本文研究的42个软枣猕猴桃品种来自不同的国家,中国22个品种,新西兰4个品种,乌克兰1个品种,日本4个品种,韩国1个品种.聚类分析在遗传距离0.64处将所有品种分为4类,这4类群基本上是以品种性状和国别进行的类别划分.此外,同一产地的品种聚在一起的较为普遍,比如产于辽宁丹东的珍玉1号、珍玉2号和珍玉3号,说明目前生产上同一地区反复使用同一授粉系或者不育系杂交配种的现象较普遍,导致群体内部基因流较大(Nm为1.454,基因流是基因在群体间的流动,是影响群体内部和群体之间遗传变异程度的重要因素).观测杂合度Ho都略低于期望杂合度He(杂合度的高低反映了居群基因型的一致性,见表1),性状相似现象亦较为普遍,造成同一地区遗传基础较差.此外,有些国产品种和国外品种聚在一起,亲缘关系较近,说明了国际合作育种工作已有成效.为了避免我国软枣猕猴桃遗传基础出现狭窄现象,未来通过加强国内外合作育种,杂交配成更加优良的软枣猕猴桃品种是非常有必要的.
目前,很多国家审定新品种都需要进行植物新品种测试(DUS测试),证明新品种的特性以及与现有品种的不同,而部分品种仅仅是育种者提供品种,经过区域试验后,对于达到要求的品种即可申请为新品种,而对于新品种的描述也只限于形态学的特征,造成品种同物异名和异物同名等现象.而利用EST-SSR为基础的分子标记技术具有明显的优势,如不含内含子,在各组织、各发育时期均可检测到,且不受季节、环境等因素的限制;数量多可覆盖整个基因组;多态性高,成共显性,能够鉴别出纯合、杂合基因型等,即使品种间形态学差异较小,只要在遗传基础上仍有差异,EST-SSR就可发挥作用.
本研究采用对叶片直接进行PCR扩增的方法,简化了PCR程序,筛选出适合鉴定真实性和纯度的4对核心引物,构建了现有软枣猕猴桃种质的DNA指纹数字图谱和EST-SSR指纹模式图谱数据库,对解决软枣猕猴桃出现的品种纠纷问题将有极大的帮助.随着软枣猕猴桃新品种的逐年出现,指纹图谱构建工作仍需不断加密,仍需要大量筛选出新的软枣猕猴桃EST-SSR核心引物,以为新品种做指纹图谱,保证我国软枣猕猴桃产业健康有序的繁荣发展.
[1] CHAT J,JAUREGUI B,PETIT R J,et al.Reticulate evoluion in kiwifruit (Actinidia,Actinidiaceae) identified by comparing their maternal and paternal phylogenies[J].American Journal of Botany,2004,91(6):736-747.
[2] 高建明,罗峰,裴忠有,等.甜高粱重要种质材料的SRAP指纹分析[J].华北农报,2010,25(2):93-98.
[3] 徐东生,华光安,刘殊,等.猕猴桃雌雄识别的多元统计分析[J].武汉植物学研究,1998,16(3):283-284.
[4] 王海飞,关建平,马钰,等.中国蚕豆种质资源ISSR标记遗传多样性分析[J].作物学报,2011(4):595-602.
[5] 王红意,翟红,王玉萍,等.30个中国甘薯主栽品种的RAPD指纹图谱构建及遗传变异分析[J].分子种植育种,2009,7(5):879-884.
[6] 旦巴,何燕,卓嘎,等.SDS法和CTAB法提取西藏黄籽油菜干种子DNA用于SSR分析[J].西藏科技,2011(8):9-11.
[7] 段艳凤,刘杰,卞春松,等.中国88个马铃薯审定品种SSR指纹图谱构建与遗传多样性分析[J].作物学报,2009,35(8):1451-1457.
[8] ELLIS J R,PASHLEY C H,BURKE J M,et al.High genetic diversity in a rare and endangered sunflower as compared to a common congener[J].Molecular Ecology,2006,15(2):2345-2355.
[9] 鲁忠富,徐沛,汪宝根,等.基于SSR分子标记技术的长豇豆种子纯度快速鉴定中的应用[J].浙江农业学报,2010(6):727-730.
[10] MAZZUCATO A,FICCADENTI N,CAIONI M.Genetic diversity and distinctiveness in tomato (SolanumlycopersicumL.) landraces:the Italian case of A pera Abruzzese[J].Hort,2010,125,55-62.
[11] 王省芬,马峙英,张桂寅,等.转基因抗虫棉SSR和AFLP遗传变异研究[J].植物遗传资源学报,2006,7(2):153-158.
[12] 文雁成,王汉中,沈金雄,等.SRAP和SSR标记构建的甘蓝型油菜品种指纹图谱比较[J].中国油料作物学报,2006(3):233-239.
[13] 王华忠,吴则东,王晓武,等.利用SRAP与SSR标记分析不同类型甜菜的遗传多样性[J].作物学报,2008(1):37-46.
[14] PROHENS J,MUNOZ-FLACON J E,RODRIGUEZ-BURRUEZO A,et al.H15 an almzgro-type pickling eggplant with high yield and reduced prickliness [J].Hortscience,2009,44:2017-2019.
[15] 陈杰,杨静,国鸿雁,等.DNA分子标记技术在烟草遗传育种中的应用[J].中国农业通报,2012(7):95-99.
[16] INCE A G,KARACA M,ONUS A N.Polymorphic microsatellite markers transferable across species [J].Plant Molecular Biology Reporter,2010,28:285-291.
[17] LATOCHLA P.The comparison of some biological features ofActinidiaargutacultivars fruit [J].Annals of Warsaw University of Life Sciences—SGGW Horticulture and Landscape Architecture,2007,28(3):105-109.
[18] LI Z Z,HUANG H W,JIANG Z W,et al.Phylogenetic relationships inActinidiaas revealed by RAPDs and PCR-RFLPs of mtDNA [J].Acta Horticulturae,2003,610(8):387-396.
[19] 盖树鹏.玉米品种纯度SSR鉴定与田间鉴定的相关性[J].华北农学报,2010,25(2):28-31.
[20] 肖小余,王玉平,张建勇,等.四川省主要杂交稻亲本的SSR多态性分析和指纹图谱的构建与应用[J].中国水稻科学,2006(01):1-7.
[21] CASTRO I,MARTIN J P,ORTIZ JM,et al.Varietal discrimination and genetic relationships ofVitisviniferaL.cultivars from two major controlled appellation (DOC) regions in Portugal [J].Hort,2011,127(3):507-514.
[22] 葛亚英,张飞,沈晓岚,等.丽穗凤梨ISSR遗传多样性分析与指纹图谱构建[J].中国农业科学,2012(4):726-733.
[23] RAIGON M D,RODRIGUEZ-BURRUEZO A,PROHENS J.effects of organic and conventional cultivation methods on composition of eggplant fruits [J].Journal of Agricultural and Food Chemistry,2010,58(1):6833-6840.
[24] 熊利荣,郑宇.基于形态学的稻谷种子品种识别[J].粮油加工,2010(06):45-48.
[25] 赵久然,王风格.玉米品种DNA指纹鉴定技术与应用[M].北京:中国农业科学技术出版社,2009:331-447.
[26] NAVASCUES M,EMERSON B C.Chloroplast microsatellites:measures of genetic diversity and the effect of homoplasy [J].Molecular Ecology,2005,14(2):1333-1341.
[27] 付小琼,杨付新,刘逢举,等.棉花总DNA提取技术及其在国家棉花区试中的应用[C].中国棉花学会年会论文汇编,2011:124-127.
[28] DOYLE J.A rapid DNA isolation procedure for small quantities of fresh leaf tissue [J].Phytochemical Bulletin,1987,19(5):11-15.
[29] 罗冉,吴委林,张旸,等.SSR分子标记在作物遗传育种中的应用[J].基因组学与应用生物学,2010,29(1):137-143.
[30] PETIT R J,DUMINIL J,FINESCCHI S,et al.Comparative organization of chloroplast,mitochondrial and nuclear diversity in plant populations [J].Molecular Ecology,2005,14(1):689-701.
[31] 王丹丹,牟元珍,杨东霞.软枣猕猴桃EST-PCR体系建立及品种遗传关系[J].辽东学院学报,2016,23(3):177-182.
(责任编辑:方林)
EstablishmentofDNAfingerprintingandanalysisofgeneticdiversityamongActinidiaargutacultivars
WANG Dan-dan1,ZHANG Yan-wen1,2
(1.College of Agriculture,Eastern Liaoning University,Dandong 118003,China; 2.Grassland Research Institute,Northeast Normal University,Changchun 130024,China)
In order to identify the cultivars ofActinidiaargutaand analyze the genetic relationship among different cultivars at a molecular level,we constructed the digital fingerprint and model fingerprint(ClusterProject)for 42 commonA.argutacultivars using EST-SSR (expressed sequence tag-simple sequence repeats) molecular markers.The 21 pairs of primers amplified a total of 132 alleles including 122 polymorphic alleles with a polymorphism rate of 92.4%.Each pairs of primers can amplify 3~8 alleles,with an average of 6.28 alleles per locus.The length of amplified products was 100~300 bp,with the mean value ofCPIC(polymorphism information content) being 0.721,Nm(gene flow) being 1.454,and the mean value ofHo(heterozygosity) being 0.691.Among the 21 primers,7 primers can be used as a preferred choice for fingerprinting,namely ACAR2,ACAR13,ACAR53,ACAR75,ACAR92,ACAR112 and ACAR120,in which 4 primers (ACAR2,ACAR13,ACAR53 and ACAR75) can be employed in combination to identify the 42 cultivars.Cluster analysis showed that the 42 cultivars can be divided into one group at a genetic distance of 0.64,and four groups at a distance of 0.81.The genetic relationship as indicated by the cluster completely reflected the characteristics and the source of the cultivars.
Actinidiaarguta;EST-SSR markers;fingerprinting;genetic diversity;cluster analysis
1000-1832(2017)03-0104-08
10.16163/j.cnki.22-1123/n.2017.03.022
2016-09-23
国家自然科学基金资助项目(31370400);丹东市2016科技攻关项目(2016KJ001);辽东学院青年基金资助项目(2015QN006).
王丹丹(1982—),女,硕士研究生,讲师,主要从事分子生物学研究.
Q 663.4 [学科代码] 180·5155
A
