砂粒粒径对日本囊对虾非特异性免疫和抗氧化酶活性的影响
2017-09-19王庚申谢建军许文军
王庚申,谢建军,何 杰,施 慧,汪 玮,许文军
(1.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江省海水增养殖重点实验室,浙江舟山 316021;2.浙江海洋大学水产学院,浙江舟山 316022)
砂粒粒径对日本囊对虾非特异性免疫和抗氧化酶活性的影响
王庚申1,谢建军1,何 杰1,施 慧1,汪 玮1,许文军2
(1.浙江海洋大学海洋与渔业研究所,浙江省海洋水产研究所,浙江省海水增养殖重点实验室,浙江舟山 316021;2.浙江海洋大学水产学院,浙江舟山 316022)
为了探究砂粒底质对日本囊对虾免疫水平的影响,实验对不同砂粒粒径处理组日本囊对虾的非特异性免疫及抗氧化酶活性差异进行了比较分析。结果表明,3个砂粒底质组日本囊对虾的ACP、AKP和LSZ酶活性均高于无底质组,并且酶活性随着砂粒粒径的增大而降低。几种抗氧化酶活性也有类似结果,砂粒底质组日本囊对虾SOD、CAT及T-AOC酶活性也随着砂粒粒径的增大而降低,而MDA酶活性随着砂粒粒径的增大而升高,说明在粗砂底质中日本囊对虾体内细胞受损程度更高。通过对各砂粒底质组日本囊对虾免疫水平的差异比较发现,粒径<0.5mm的细砂更适合在日本囊对虾工厂化和高位池精养模式中应用。
日本囊对虾;砂粒粒径;非特异性免疫;抗氧化酶
日本囊对虾Marsupenaeus japonicus俗称花虾、竹节虾、车虾等,具有生长快、耐低温、耐干露能力强等优点,养殖潜力巨大,是我国重要的虾类养殖品种之一[1]。其营养丰富,味道鲜美,市场价格高,且具生长快、耐低温、耐干露能力强等特点,深受消费者和养殖业者的喜爱。我国20世纪80年代开展全人工养殖以来,沿海各省市均有养殖,并形成一定的规模[2]。经过多年的养殖实践和摸索,日本囊对虾养殖技术不断完善,池底覆砂的工厂化和高位池精养模式已成为其新兴的养殖模式[3]。
日本囊对虾具有潜沙和底质选择等生物学特性,对底质要求较高,马宜山等[4]研究认为沙底质和泥沙底质更利于日本囊对虾的生长,臧维玲等[5]则通过观察实验发现日本囊对虾幼虾更喜欢细砂底质。目前有关日本囊对虾底质选择的研究多集中在对生长和成活的影响[6],而针对砂粒底质对日本囊对虾免疫水平影响的研究尚未见报道。本研究拟设置不同粒径的砂粒底质,比较不同砂粒粒径条件下日本囊对虾免疫酶活性的变化情况,以期为日本囊对虾工厂化和高位池精养模式提供理论参考。
1 材料与方法
1.1 实验材料
实验于2016年7月22日-8月12日在浙江省海洋水产研究所西轩岛试验场进行,所用日本囊对虾取自试验场精养塘,平均体长 2.76±0.18 cm,平均体质量 0.27±0.03 g,于室内水泥池中暂养 7 d。暂养期间,微充气,日换水量30%,每天06:00和18:00,过量投喂天邦牌对虾配合饲料,3 h后吸出残饵和粪便。实验用砂购于市场,预先经不同规格的筛网筛选,并经高锰酸钾浸泡消毒,清水冲洗,晾干后使用。
1.2 实验设计
实验容器为160 L塑料水槽,规格为76 cm×55 cm×45 cm。本实验设3个覆砂组(LGS组:粒径>2.5 mm;MGS组:粒径 1~2 mm;SGS组:粒径<0.5 mm),砂层厚度为 10 cm,1个对照组(NS组:无砂),每组设 3个重复。暂养结束后,每个水槽内放养规格相近的健康幼虾50尾,适应24h后开始正式实验。实验期间,每天06:00和18:00投喂天邦牌对虾配合饲料,日投饵量根据摄食情况而定,其它条件与暂养期间相同。分别于实验的第0 d、5 d、10 d、15 d、20 d收集日本囊对虾全虾,每个平行取2尾组成混合样,全部样品-80℃保存待测酶活性。
1.3 样品处理
准确称量样品,按重量体积比加入9倍的1.5%生理盐水组织匀浆,2 500 r/min离心10 min,取上清稀释后用于免疫酶活性测定。
1.4 免疫酶活性测定
免疫酶活性采用试剂盒测定,购自南京建成生物工程研究所。酸性磷酸酶(ACP)和碱性磷酸酶(AKP)活性测定采用磷酸苯二钠法[7];溶菌酶(LSZ)以溶壁微球菌冻干粉为底物,采用HULTMARK等[8]的方法改进进行;超氧化物歧化酶(SOD)测定采用黄嘌呤氧化法;谷胱甘肽过氧化物酶(GSH-px)采用5-二硫代硝基苯甲酸法测定;过氧化氢酶(CAT)活性的测定采用钼酸铵法;丙二醛(MDA)的测定采用硫代巴比妥酸法;总抗氧化能力(T-AOC)采用ABTS法测定;蛋白含量的测定采用考马斯亮蓝法。
1.5 统计分析
用SPSS 20.0软件进行单因素方差分析及Duncan多重比较进行数据分析处理,以P<0.05表示差异显著。
2 结果
2.1 砂粒粒径对日本囊对虾非特异性免疫的影响
不同处理组日本囊对虾ACP活性在前5 d变化趋势有所差异,SGS组和MGS组ACP酶活升高,LGS组和NS组则明显下降;5 d之后,各组日本囊对虾ACP酶活均先降后升。4个处理组日本囊对虾平均ACP酶活大小依次为SGS组、MGS组、LGS组和NS组。
各处理组日本囊对虾AKP酶活如图1-b所示。SGS组和MGS组AKP酶活基本保持平稳,下降幅度较小,而LGS组AKP酶活在试验过程中持续下降;NS组在前5 d快速下降,随后保持平稳。试验前期,3个砂粒处理组AKP活性显著高于NS组(P<0.05),但试验结束时LGS组AKP活性低于NS组。
不同处理组日本囊对虾LZM酶活变化趋势有所不同(图1-c)。SGS组和MGS组LZM酶活在前10d出现下降,MGS组下降幅度大于SGS组(P>0.05);10d之后两组LZM酶活开始上升,并保持稳定。LGS组LZM酶活在前15 d持续下降,之后开始上升,NS组则在前10 d快速下降,随后波动幅度较小。4个处理组日本囊对虾平均 LZM 酶活分别为 SGS 组(38.57±3.01)、MGS 组(35.19±5.55)、LGS 组(33.48±10.63)、NS 组(28.01±8.38)。
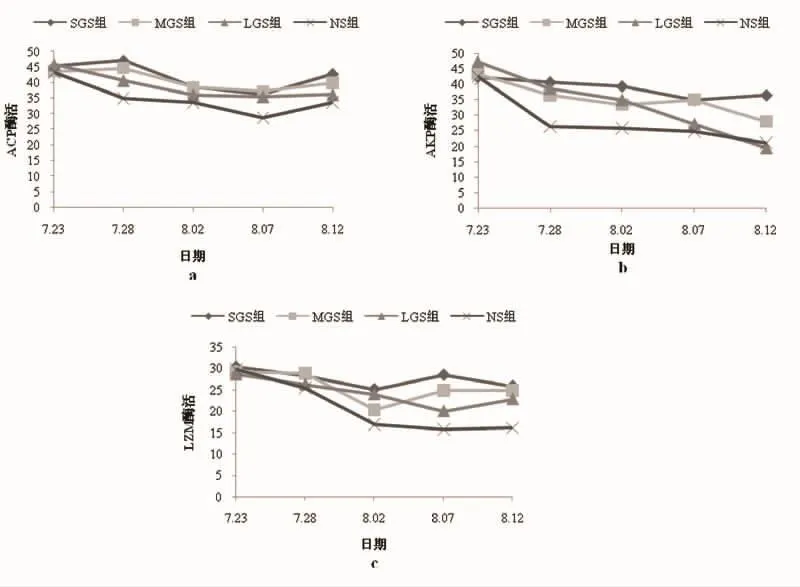
图1 砂粒粒径对日本囊对虾非特异性免疫的影响Fig.1 Effect of grand size on non-specific immune enzymes activities of M.japonicus
2.2 砂粒粒径对日本囊对虾抗氧化酶活性的影响
各处理组日本囊对虾SOD酶活变化情况如图2-a所示。SGS组和MGS组日本囊对虾SOD酶活在试验期间先升后降,但总体波动幅度较小;NS组和LGS组SOD酶活则在前5 d明显上升,随后开始逐渐下降,波动幅度大于 SGS组和 MGS 组(P>0.05)。
4个处理组日本囊对虾GSH-PX酶活在前10 d均先升后降,NS组升高幅度显著高于砂粒底质组(P<0.05);SGS组和MGS组GSH-PX酶活在10 d之后基本保持平稳,LGS组和NS组则再次出现明显的波动。4个处理组日本囊对虾平均GSH-PX酶活为SGS组<MGS组<LGS组<NS组。
不同砂粒处理组日本囊对虾CAT酶活变化趋势基本一致,前期先升后降,至15 d时达到最低,随后开始升高。LGS组和NS组日本囊对虾CAT酶活波动幅度大于SGS组和MGS组(P<0.05)。4个处理组日本囊对虾平均GSH-PX酶活大小依次为SGS组、MGS组、LGS组、NS组。
各处理组日本囊对虾MDA活性变化情况见图2-d。SGS组、MGS组和NS组日本囊对虾MDA活性变化差异不显著(P>0.05),在前5 d出现升高,随后波动下降,而LGS组MDA活性在试验期间则先升后降,波动幅度与其他处理组差异显著(P<0.05)。4个处理组日本囊对虾平均GSH-PX酶活分别为SGS组(0.94±0.47)、MGS 组(0.96±0.42)、LGS 组(1.41±0.35)、NS 组(1.13±0.56)。
不同处理组日本囊对虾T-AOC活性变化情况有所不同,SGS组T-AOC活性在试验期间变化较小,仅在第10 d时出现波动下降。MGS组、LGS组和NS组在前10 d均先升后降,波动幅度差异显著(P<0.05),10 d之后基本保持稳定,变化幅度较小(P>0.05)。不同处理组日本囊对虾平均T-AOC活性分别为SGS组(5.48±0.63)、MGS 组(4.35±0.55)、LGS 组(3.97±1.68)、NS 组(4.33±2.13)。

图2 砂粒粒径对日本囊对虾抗氧化酶活性的影响Fig.2 Effect of grand size on antioxidant enzymes activities of M.japonicus
3 讨论
研究表明,外界因素的胁迫可以影响对虾的免疫功能,比如温度、盐度、外界刺激等[9-10],而目前有关砂粒底质对对虾免疫影响的研究还比较少。底质是对虾主要的生长、栖息和活动场所,在对虾养殖系统中占有重要地位。张沛东等[11]认为底质的结构和特征与对虾的行为活动关系密切,在高密度精养条件下,适宜的底质可以有效地减少因养殖密度增加而引发的机体健康状况和免疫力下降。因此,本文对不同粒径砂粒底质对日本囊对虾的免疫功能的影响进行了研究。
对虾的免疫反应比较原始,体内不能产生免疫球蛋白,为保护自身免受伤害,机体形成了非特异性的免疫保护机制。对虾的非特异性免疫主要包括体液免疫和细胞免疫,其中体液免疫主体是血淋巴中的酶、免疫因子和调节因子[12]。当外来异物侵入机体时,识别因子会先进行识别,然后诱导血细胞合成免疫因子,最后经过一系列的免疫反应清除外来病原体,从而达到免疫的目的[13]。ACP、AKP、LSZ等都是对虾体内重要的非特异性免疫酶。ACP、AKP是两种重要的代谢调控酶,同时也是巨噬细胞溶酶体酶的标志酶,通过形成水解酶体系消除异物,从而起到免疫防御的功能[14-15]。研究发现,3个不同粒径的砂粒组日本囊对虾的ACP、AKP和LSZ酶活均高于无底质组;而在3个砂粒组中,非特异性免疫酶活随着砂粒粒径的增大而降低,究其原因可能是,在粗砂组和无底质组中日本囊对虾的潜沙行为受到抑制,影响了机体对环境变化的调节与整合,从而导致机体非特异性免疫能力下降[11]。另外还发现试验期间3个砂粒底质组日本囊对虾的ACP、AKP和LSZ酶活基本呈现波动震荡的趋势,且波动幅度较小,而无底质组酶活则均出现了明显的下降,说明砂粒底质更有利于维持非特异性免疫酶活性的稳定。
MDA是脂质过氧化物分解后的重要产物,间接反映细胞的受损程度,其含量的测定常与SOD和CAT的测定配合使用[16]。本实验中,LGS组日本囊对虾的平均MDA酶活最高,其次是NS组,说明在LGS组和NS组日本囊对虾细胞受损的程度更高,虾体免疫防御能力下降。SOD是一种重要的抗氧化酶,其活性高低间接反映了机体清除活性氧自由基的能力[17],它能够让活性氧发生歧化,生成过氧化氢和氧气,而GSHPX和CAT可以将过氧化氢还原成水,三者的联合使用可以有效防止生物细胞受自由基的损伤[18]。T-AOC是综合反映了机体非酶抗氧化系统和抗氧化酶系共同完成抗氧化作用。本研究中,实验前期各试验组SOD、CAT及T-AOC酶活均呈现升高趋势,升高幅度依次为NS组、LGS组、MGS组和SGS组,这可能与日本囊对虾在无底质组和粗砂粒组受到的外界刺激更多,导致机体细胞产生了大量的活性氧自由基,从而诱导了抗氧化酶的合成[19]。实验中后期,SOD、CAT及T-AOC酶活则随着砂粒粒径的增加而下降,究其原因可能是当日本囊对虾长期处于外界胁迫环境中,抗氧化酶活性不能无限制上升,达到峰值后会出现下降,即有学者提出的“免疫疲劳”现象[20];另一方面持续的外界底质刺激会使虾体生理机能失调,也会导致抗氧化酶活性的下降[17]。
本研究结果显示,3个砂粒底质组日本囊对虾的非特异性免疫酶活(ACP、AKP和LZM)均高于无底质组,其中以SGS组最高,同时SGS组的SOD、CAT及T-AOC酶活也明显高于其他试验组,MDA则低于他其试验组,说明SGS组日本囊对虾的免疫能力最高,细胞受损程度最小。因此,综合比较认为,粒径<0.5 mm的细砂更适合在日本囊对虾工厂化和高位池精养模式中应用。
[1]王克行.虾蟹类增养殖学[M].北京:中国农业出版社,1997.
[2]李鸿鹏,富 裕,任夙艺,等.舟山近海日本囊对虾野生群体与越冬养成群体形态性状对体重和肉重影响的比较[J].海洋与湖沼,2015,46(5):1 218-1 227.
[3]孙成波,李 婷,李义军,等.高位池精养日本囊对虾的生长规律[J].大连海洋大学学报,2011,26(4):316-321.
[4]马宜山,高锡伦,曹大铮.不同底质对日本对虾生长的影响[J].水产养殖,1990(5):17-18.
[5]臧维玲,戴习林,姚庆祯,等.底质对日本对虾幼虾生长的影响[J].上海海洋大学学报,2003,12(1):72-75.
[6]徐 涵,熊 慧,吴亚林,等.养殖密度、底质类型对日本囊对虾生长、存活的影响[J].湖北农业科学,2015,54(4):923-925.
[7]KRUZEL M,MORAWIECKA B.Acid phosphatase of potato tubers(Solanum tuberosum L).Purification,properties,sugar and amino acid composition[J].Acta Biochimica Polonica,1982,29(3/4):321-330.
[8]HULTMARK D,STEINER H,RASMUSON T.Studies on the method of lysozyme measurement in serum[J].Eur J Biochem,1980,106(6):7-16.
[9]王专伟,黄建华,杨其彬,等.感染白斑综合征病毒的斑节对虾免疫酶变化特征[J].湖北农业科学,2011,50(9):1 851-1 854.
[10]陆承平.动物保护概论[M].北京:高等教育出版社,1999.
[11]张沛东,张 倩,张秀梅,等.底质类型对中国明对虾存活、生长及行为特征的影响[J].中国水产科学,2014,21(5):1 079-1 086.
[12]马寨璞,张繁霜,佟霁坤.甲壳动物免疫系统概述及其图论表示[J].安徽农业科学,2011,39(1):291-294.
[13]丁金强,刘 萍,李 健,等.不同地理群体日本蟳非特异性免疫及抗氧化酶活力的比较[J].水产学报,2013,37(2):275-280.
[14]张龙岗,安 丽,孙 栋,等.盐度对澳洲宝石鲈幼鱼3个免疫因子活力的影响[J].水生态学杂志,2011,32(6):110-114.
[15]刘树青.免疫多糖对中国对虾血清溶菌酶、磷酸酶和过氧化物酶的作用[J].海洋与湖沼,1999,30(3):278-283.
[16]MATHEW S,KUMAR K A,ANANDAN R,et al.Changes in tissue defence system in white spot syndrome virus(WSSV)infected Penaeus monodon[J].Comparative Biochemistry&Physiology Part C:Toxicology&Pharmacology,2007,145(3):315-320.
[17]叶建生,王兴强,马 甡,等.盐度突变对凡纳滨对虾非特异性免疫因子的影响[J].渔业科学进展,2008,29(1):38-43.
[18]祁克宗,王林安.自由基和生物抗氧化系统理论与外科学的关系[J].安徽农业大学学报,1996(2):171-174.
[19]叶继丹,韩友文,赵吉伟,等.喹乙醇对鲤肝胰脏抗氧化酶系统的影响[J].水产学报,2004,28(3):231-235.
[20]CHANG C F,CHEN H Y,SU M S,et al.Immunomodulation by dietary β-1,3-glucan in the brooders of the black tiger shrimp Penaeus monodon[J].Fish&Shellfish Immunology,2000,10(6):505-514.
Effects of Non-Specific Immunity and Activities of Antioxidant Enzymes of Marsupenaeus japonicus in Response to Grain Size
WANG Geng-shen,XIE Jian-jun,HE Jie,et al
(Marine and Fishery Research Institute of Zhejiang Ocean University,Marine Fisheries Research Institute of Zhejiang Province,Key Laboratory of Marinculture and Enhancement of Zhejiang Province,Zhoushan 316021,China)
In order to investigate the effect of grainsize into immune status of Marsupenaeus japonicus,the activities of non-specific immune enzymes and antioxidant enzymes in different grain size groups(LGS:>2.5mm,MGS:1-2 mm,SGS:<0.5 mm,NS:control)were studied.Three non-spectific immune enzymes(ACP,AKP and LSZ)and five antioxidase enzymes(SOD,CAT,GSH-px,MDA and T-AOC)of M.japonicuswere determinined.The results showed that the activitiesof non-spectific immune enzymesin grand groups were higher than thecontrol group;and the activities were gradually decreased with the increasing grand size.The SOD,CAT and T-AOC activities were also decreased gradually with the increasing grand size.The MDA activity was increased gradually with the increasing grand size.That indicated the cell damage of M.japonicus in coarsegrain was higher than that in the fine grain.Therefore,fine sandwith grain size less than 0.5 mm is suggested for cultivating M.japonicus in the factory and high pool intensive mode.
Marsupenaeus japonicus;grandsize;non-spectific immunity;antioxidase
S968.22
A
1008-830X(2017)02-0109-06
2017-01-02
浙江省科技厅项目(2015F30003;2015F50004)
王庚申(1988-),男,河南周口人,工程师,研究方向:海水健康养殖.E-mail:wgs-1988@163.com
许文军,教授级高级工程师.E-mail:xwenjun@sina.com
