新疆南部塔河流域柳树腐烂病病原菌的鉴定
2017-09-18邵延慧张王斌刘振亚朱宗财严海璘李亚鹏曹琦但红侠
邵延慧,张王斌,刘振亚,朱宗财,严海璘,李亚鹏,曹琦,但红侠
(塔里木大学/塔里木大学南疆农业有害生物综合治理重点实验室,新疆阿拉尔 843300)
新疆南部塔河流域柳树腐烂病病原菌的鉴定
邵延慧,张王斌,刘振亚,朱宗财,严海璘,李亚鹏,曹琦,但红侠
(塔里木大学/塔里木大学南疆农业有害生物综合治理重点实验室,新疆阿拉尔 843300)
【目的】通过对病原菌形态学和培养形状观察,结合分子生物技术ITS和β-tubulin序列分析,明确新疆南疆柳树腐烂病菌种类和遗传分化。【方法】采集柳树腐烂病并获得病菌纯培养,对腐烂病菌分生孢子器进行纵横切片并观察记录形态数据,将病原菌置于不同营养条件和环境条件下观察记录其培养形状,采用ITS和β-tubulin序列分析方法结合,构建系统发育树。【结果】分子孢子器和分子孢子形态特征与金黄壳囊孢菌(C.chrysospermaPer.ex Fr.)一致;菌丝生长最适温度20~30℃,最适pH值为5~6,最适碳氮源分别是葡萄糖和蛋白胨;经β-tubulin序列分析,与壳囊孢属不同种类的序列相似度为96%以上;在室内离体接种条件下病原菌均能成功侵染其他5种树木。【结论】引起新疆南部塔河流域柳树腐烂病原菌为金黄壳囊孢菌(C.chrysospermaper.exFr),不同来源的病原菌存在一定的遗传分化,在室内可侵染杨树、胡杨、枣树、梨和苹果,并可在胡杨和杨树上产生分生孢子器。
柳树腐烂病;形态特征;培养性状;ITS;β-tubulin
0 引 言
【研究意义】新疆南部荒漠生态环境极为脆弱,防护林为绿洲农业的种植提供了有效的保障[1]。近年来发现新疆主栽防护林柳树腐烂病越发严重,已经制约了新疆南部荒漠绿洲生态区特色果品的可持续发展[2-3],对柳树腐烂病菌进行鉴定,明确其种类和来源,对新疆南部荒漠绿洲生态区特色果园合理布局具有重要意义。【前人研究进展】邢红亮等[4]研究表明核桃腐烂病菌能够成功侵染杨树、柳树、苹果树、枣树、胡杨、梨树等多种树木枝条,说明该病菌的寄主广泛;桂腾茸等[5]对苹果树腐烂病菌的生物学特性及致病性进行了研究,腐烂病菌优势种是V.Ceraterapema,其种内出现了明显的菌群分化和致病力分化现象;王卫雄[6]通过形态学和rDNA-ITS、EF1-α以及β-tubulin三种基因序列分析,确定了甘肃省苹果树腐烂病菌分为2类;刘钰娇[7]通过rDNA-ITS基因序列分析,明确河北省苹果树腐烂病菌为Valsamali;唐俊煜[8]通过rDNA-ITS、EF1-α以及β-tubulin基因序列构建系统进化树发现杨树、柳树、枣树等林木腐烂病菌全部由C.chrysosperma(有性型V.sordida)引起。【本研究切入点】国内关于柳树腐烂病的研究较少,相关研究多集中于经济效益较高的果树,近些年关于柳树腐烂病的研究多涉及该病害的防治及病害的发生规律等[9]。研究柳树腐烂病菌进行鉴定以及培养性状。【拟解决的关键问题】研究柳树腐烂病菌的鉴定、培养性状及致病性,为柳树腐烂病的防治以及果树防护林树种的选择提供依据。
1 材料与方法
1.1材料
从温宿(N 41°16' 15.92",E 80°17' 50.63")、第一师阿拉尔市(N 40°32' 35.98",E 81°18' 0.69")、十二团(N 40°29' 46.95",E 81°16' 1.15")和三团(N 40°22' 1.37",E 80°3' 27.37")防护林选取柳树腐烂病的发病枝条和树皮4份,用采集刀刮取病皮( 含健康组织) 或用枝剪剪取病枝条,放于牛皮纸标本袋中备用。
1.2 方 法
1.2.1 病原菌的分离
用常规组织分离法获得病原菌纯培养[10],保存备用。
1.2.2 致病性测定
采用离体枝条接种法[11],置于25℃下培养,每隔24 h观察并记录病斑扩展、产孢体形成情况。
1.2.3 子实体形态特征观察
对从田间采集到病组织上的产孢体进行切片观察,在显微镜(13395H2X )下观察分生孢子器外部形态、腔室数量及纵横切面形态特征并测量其大小;观察分生孢子形态并测量孢子大小。
1.2.4 不同条件下病菌生长特性观察
1.2.4.1 不同环境条件下病菌生长特性观察
将供试菌株接于不同 pH 值(2、3、4、5、6、7、8、9、10 )和不同温度(5、15、20、25、30、35和40℃)条件下的 PDA 平板上,置于 25℃,暗培养,每处理 3 个重复,每隔24 h测量菌落直径,记录菌落形态特征。
1.2.4.2 不同营养条件(C/N源)下菌丝生长特性观察
以查彼克培养基为基础培养基,将供试菌株分别接于不同碳源(葡萄糖、麦芽糖、淀粉代替蔗糖和空白对照)、不同氮源(分别用等量( 0.13 g /L ) 的蛋白胨、甘氨酸、硝酸钾代替硝酸钠和空白对照)条件下,按照常规方法接种到培养基上,每皿接1个菌饼。每处理3次重复,25 ℃ 条件下培养,每隔24 h测量菌落直径并记录菌落的形态特征。
1.2.5 分子生物学鉴定
1.2.5.1 菌丝的培养和DNA提取
腐烂病菌菌丝的培养和基因组DNA的提取,参照王卫雄[6]的方法进行。
1.2.5.2 基因片段的 PCR 扩增
病原菌ITS序列扩增的引物ITS1和ITS4以及扩增条件参照殷辉等[12]的方法进行;应用引物Beta-F和Beta-R扩增β-tubulin序列,扩增条件均参照郭开发等[13]的方法进行。
将不同引物扩增的PCR产物送到华大基因科技有限公司进行测序。获得序列后,登陆NCBI网站,进行BLAST同源性比对分析,同时下载同源性序列,选取PCR扩增的500 bp片段,使用MEGA5.02软件以邻接法建立进化树。
2 结果与分析
2.1 子实体形态特征
对采集四份病原菌进行分离和鉴定,表明其分生孢子均呈腊肠形,无色,单胞,具有13~20个数量不等的腔室,腔室大小不一,形状不规则,共用腔室璧,仅一个孔口通向表皮外,大小为4.75 μm~5.89 μm×1.40 μm~3.21 μm。分生孢子横切面大小为2 380.78 μm~1 000.40 μm×2 268.13 μm~890.10 μm,纵切面大小3 318.20 μm~708.08 μm×670.49 μm~338.07 μm。根据《植物病原真菌学》[14]的描述和范鑫磊[15]、张庆涛[16]的研究结果,柳树腐烂病菌病原与金黄壳囊孢(C.chrysospermaPer.exFr.)相似度最高。图1~3
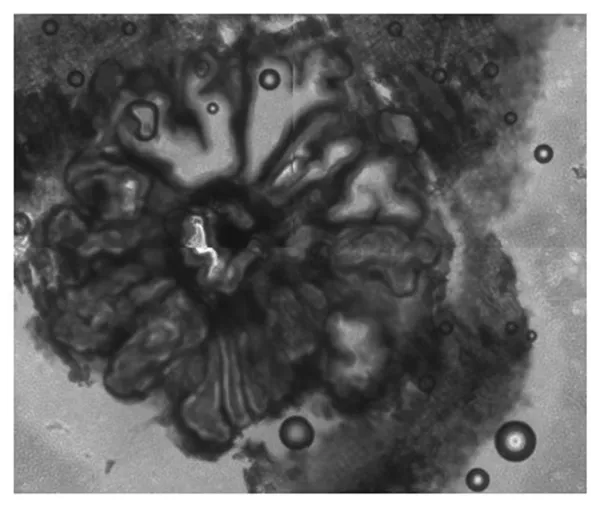
图1 分生孢子器横切片
Fig.1 Pycnidium cross section

图2 分生孢子器纵切片
Fig.2 Pycnidium longitudinal section
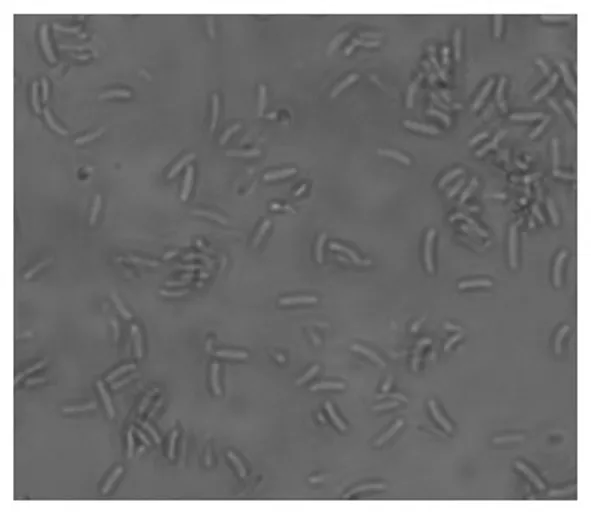
图3 分生孢子
Fig.3 Conidia
2.2 不同条件下菌丝生长特性
2.2.1 不同温度/pH对菌丝的长的影响
研究表明,从采集的四份柳树发病枝条分离获得柳树腐烂病菌在5~15℃时是低温抑制菌落的生长,35~40℃时是高温抑制菌落的生长。在20~30℃时菌落生长较快,菌丝质密且色素分泌较多,皿底呈浅褐色。在5~15℃、35~40℃时菌落的生长变化巨大且在两个抑制生长的温区菌落生长慢且产生色素少。图4
研究表明,病菌在pH 2~10均可生长,最适pH为5~7,在pH为6时生长最快,而且产孢最多。在pH小于5区间和pH大于7区间病菌生长受到不同程度的抑制,说明柳树腐烂病菌不适合在碱性条件下生长。图4

图4 温度/pH对柳树腐烂病菌菌落生长的影响
Fig.4 Effect of temperature/pH on the growth of rot pathogen of willow
2.2.2 碳源/氮源对菌丝生长的影响
研究表明,病菌对麦芽糖、淀粉利用率很低。在72 h内观察,不同碳源培养基上生长量存在差异,在葡萄糖为碳源的培养基上菌丝颜色乳白、匍匐生长、分泌色素较多,生长最快且产孢快,菌落呈圆形辐射状生长。以麦芽糖,淀粉为碳源的培养基上菌丝透明、匍匐生长、不分泌色素,产孢慢,菌落呈圆形辐射状生长。在无碳条件下菌落生长速度较为缓慢,菌丝稀疏,菌落为无色透明状,产孢慢,呈圆形辐射状生长。由此说明以葡萄糖为最适碳源。
病菌在以蛋白胨为氮源的培养基上生长最好,起初菌丝透明,后变为乳白色且匍匐生长,产孢快,色素为浅褐色。在以硝酸钾、甘氨酸为氮源的培养基上菌丝透明,色素分泌较少,生长较慢,菌落呈匍匐状生长,产孢慢。由此说明以蛋白胨为最适氮源。表1
表1 碳源/氮源下菌丝生长变化
Table 1 Effect of carbon/nitrogen sources on hyphae growth
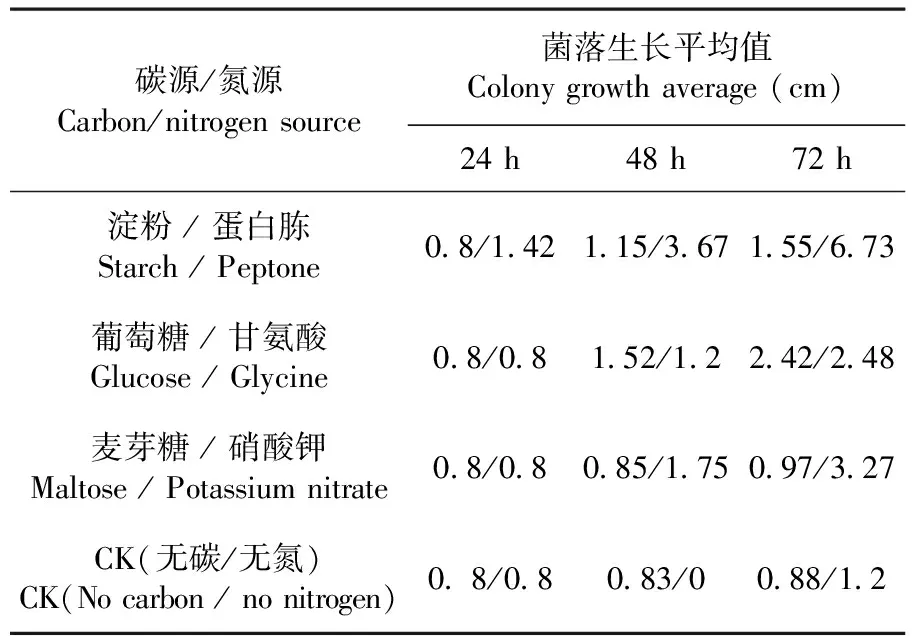
碳源/氮源Carbon/nitrogensource菌落生长平均值Colonygrowthaverage(cm)24h48h72h淀粉/蛋白胨Starch/Peptone0 8/1 421 15/3 671 55/6 73葡萄糖/甘氨酸Glucose/Glycine0 8/0 81 52/1 22 42/2 48麦芽糖/硝酸钾Maltose/Potassiumnitrate0 8/0 80 85/1 750 97/3 27CK(无碳/无氮)CK(Nocarbon/nonitrogen)0 8/0 80 83/00 88/1 2
2.3 DNA分子序列
利用ITS和β-tubulin两个基因引物腐烂病菌进行PCR扩增,均可获得明亮清晰的条带,片段大小为540和500 bp,将所得的序列在GenBank中通过Blast比对,下载相似性较高的序列并与4株菌株,以phomopsissp序列作为外群,利用MEGA5.02软件,采用邻接法构建系统发育树,进行系统发育分析。在基于ITS序列构建的系统发育树中,4株菌株与下载的序列聚在一支,说明4株菌株在ITS核苷酸序列上分化较小,ITS序列提供的信息位点不能区分供试病原菌的种间差异,因此无法将4株菌株鉴定到具体的种类;在基于β-tubulin序列构建的系统发育树中,菌株LI01与C.chrysosperma(KM034883、KF498689)、菌株LI02和LI03与C.chrysosperma(KM034885、JQ900372)、菌株LI04与C.chrysosperma(KF039741)聚在不同的分支,表明不同柳树的腐烂病菌株存在一定的遗传分化。图5~7
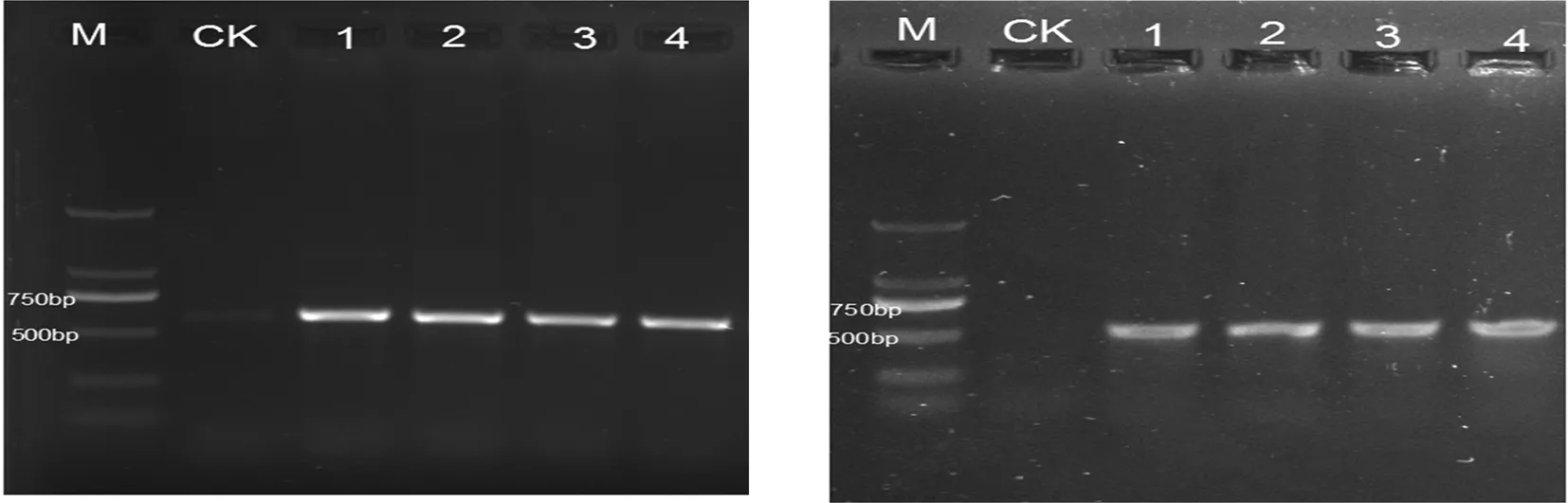
图5 柳树腐烂病菌PCR扩增结果
Fig.5 PCR products of ITS、β-tubulin of C.chrysosperma

图6 ITS序列构建的系统发育树(NJ)
Fig.6PhylogramusingITSsequencesdata
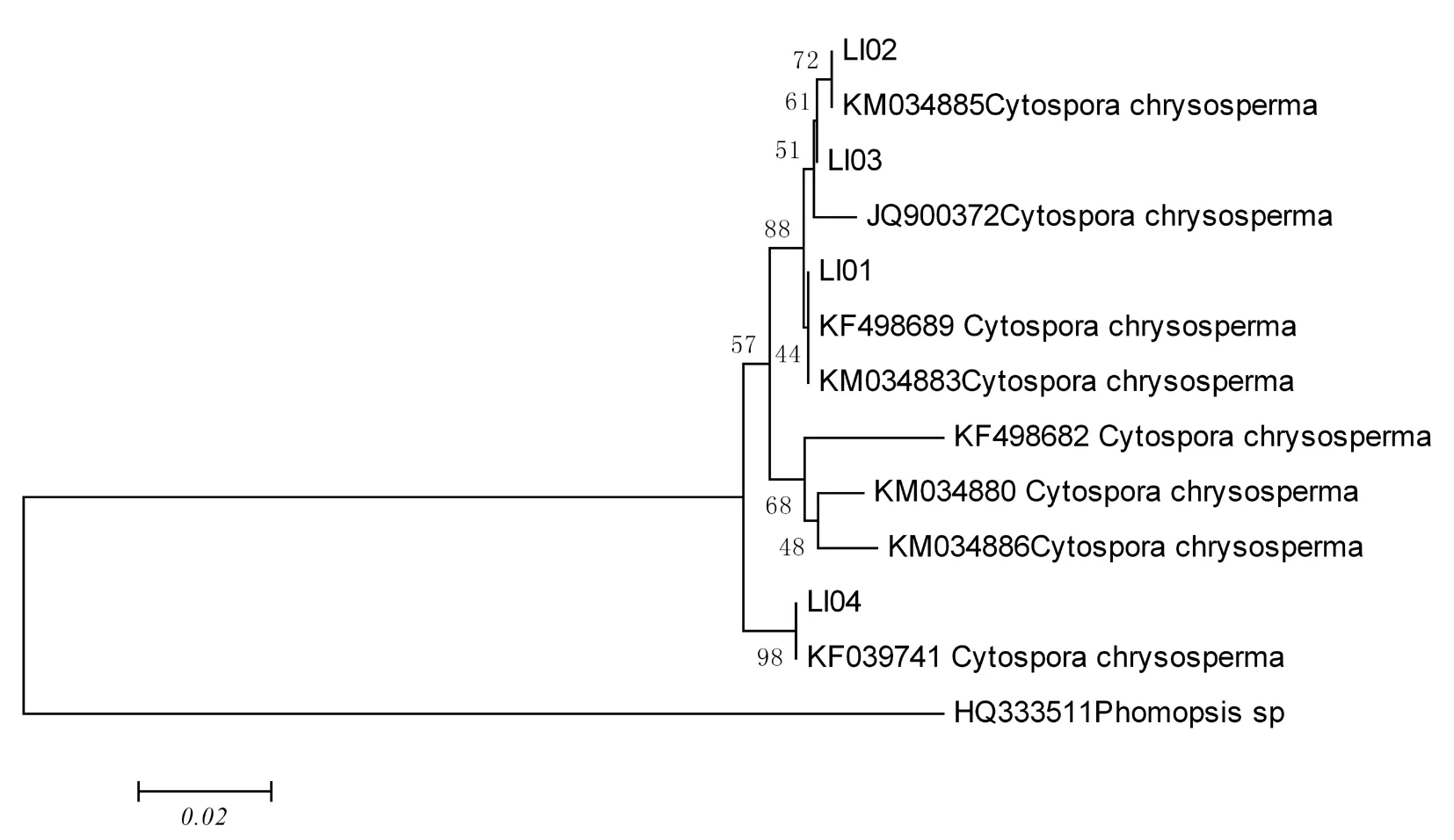
图7 β-tubulin序列构建的系统发育树(NJ)
Fig.7 Phylogram using β-tubulin sequences data
2.4病菌对新疆南部塔河流域主栽果树和防护林树木致病性
研究表明,将该病菌在25℃接种于6种寄主枝条,观察其病斑的扩展速度与病斑颜色间的差异及是否产生分生孢子角。杨树、胡杨最先出现病斑,胡杨的病斑扩展速度比杨树的快,柳树病斑的扩展速度较胡杨和杨树缓慢;胡杨最先产孢,分生孢子角为橘黄色,杨树枝条产孢晚于胡杨,分生孢子角为橘黄色,产孢时间略有差异,胡杨最快杨树次之,柳树最慢。苹果树、梨树和枣树均可产生病斑,但不产生分生孢子角。接种7 d后出现褐色、水渍状病斑,随后病斑向四周扩展,病斑部位表皮皱缩,病健交界明显。12 d能病斑表面均能产生小黑点。表2
表2 柳树腐烂病菌对不同寄主的致病性测定
Table 2 Measuring results of Willow valsa cankers rot pathogenicity of Fusarium moniliforme on different trees
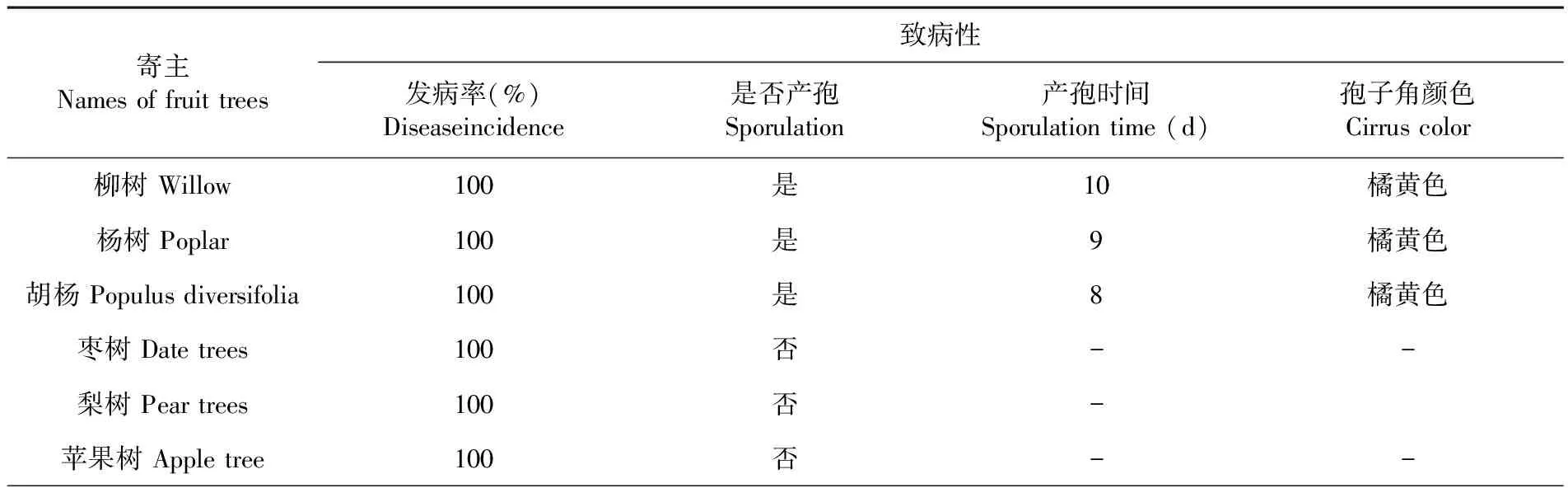
寄主Namesoffruittrees致病性发病率(%)Diseaseincidence是否产孢Sporulation产孢时间Sporulationtime(d)孢子角颜色Cirruscolor柳树Willow100是10橘黄色杨树Poplar100是9橘黄色胡杨Populusdiversifolia100是8橘黄色枣树Datetrees100否--梨树Peartrees100否-苹果树Appletree100否--
注:-表示未产孢
Note: - Indicates that no produce spores
3 讨 论
根据病原菌分生孢子器腔室形态、孔口数量和分生孢子等相对较稳定的形态特征对供试菌株进行鉴定,研究结果与范鑫磊[15]和张庆涛[16]对金黄壳囊孢菌形态特征的研究结果相近;根据前人研究发现培养性状也可以作为种类鉴定的辅助依据,因此,对供试菌株的培养性状进行研究,其菌丝生长最适温度20~30℃,最适pH值为5~6,最适碳氮源分别是葡萄糖和蛋白胨,研究结果与冀瑞卿等[17]对金黄壳囊孢菌的培养性状结果一致。结合形态特征和培养性状,初步确定供试菌株可能是金黄壳囊孢菌或是相似种。
壳囊孢属Cytospora种类较多,分布范围广,种间的形态特征和培养性状具有很高的相似性,仅依靠传统形态学准确鉴定十分困难[13]。因此,研究结合ITS和β-tubulin序列片段对病原菌进行鉴定,通过ITS序列分析发现,供试菌株ITS序列与C.chrysosperma(KP114125、KP114129、KP114134、KP114124)、C.tritici(KF293771)、V.sordida(AB369506)、V.nivea(KF293929)序列高度相似,系统发育树结果与之一致,因此,仅通过ITS序列片段无法将病原菌鉴定到具体的种类,说明在真菌分类中采用ITS序列片段进行分析时存在一定的局限性,在其序列片段上存在一定的保守区,难以区分壳囊属种间复杂的差异性,因此在研究系统发育时需使用多基因分析法,才能使结果更加准确,所以使用β-tubulin序列对供试菌株进行分析,发现4株菌株与不同登陆号的C.chrysosperma高度相似,在系统发育树也聚在不同的分支上,说明4株菌株亲缘关系存在差异,这种差异与菌株的地理来源有一定的关系。
致病性的测定中发现,柳树腐烂病接种在梨树植条上只产生病斑而不产生产孢器,与唐俊煜[8]在柳树植条上接种香梨腐烂病只产生病斑不产生产孢器,说明二者相互侵染的能力较弱,可以在梨园周围种植柳树作为防护林。
4 结 论
4.1 通过形态特征、培养性状、结合分子生物学辅助鉴定方法,将柳树腐烂病菌准确鉴定为金黄壳囊孢(C.chrysospermaPer.exFr.),采自不同地点的病原菌之间存在一定的遗传分化。
4.2 对供试菌株的培养性状进行研究,研究结果为供试菌株菌丝生长最适温度20~30℃,最适pH值为5~6,最适碳氮源分别是葡萄糖和蛋白胨。
4.3 柳树腐烂病病原菌可使杨树、胡杨发病严重;使枣树、梨树和苹果树发病但不产生分生孢子器。在生产实际中提倡在梨园、枣园和苹果园周围可以种植柳树为防护林,在防护林的建设时尽量减少柳树与胡杨和杨树混栽。
References)
[1] 李诚志.新疆土地沙漠化监测与预警研究[D].乌鲁木齐:新疆大学博士论文,2012.
LI Cheng-zhi.(2012).StudyontheDesertificationMonitoringandEarly-warninginXingjiang[D]. PhD Thesis. Xinjiang University, Urumqi. (in Chinese)
[2] 董立新.浅析冀北绿化柳树腐烂病发病原因及防治[J]. 中国农业信息,2013,(7):135.
DONG Li-xin. (2013).Analysis on the Causes and Prevention of Deciduous Disease of Willow in North Hebei Province [J].ChinaAgriculturalInformation, (7):135. (in Chinese)
[3] 董汉伟.北方地区果树病虫害环保防治对策[J]. 现代农业科技,2014,(13):155-156.
DONG Han-wei.(2014).Environmental Protection and Control Countermeasures of Fruit Trees and Pests in Northern China [J].ModernAgriculturalSciencesandTechnology, (13):155-156. (in Chinese)
[4] 邢红亮,张王斌,温善菊,等.新疆南疆桃树腐烂病菌生物学特性研究[J].新疆农业科学,2015,52(12):2 286-2 293.
XIN Hong-liang, ZHANG Wang-bin, WEN Shan-ju, et al.(2015).Biological Characteristics and Pathogenicity of Valsa Canker Pathogen of Peach Tree in Southern Xinjiang [J].XinjiangAgriculturalSciences, 52(12): 2,286-2,293. (in Chinese)
[5] 桂腾茸,孔宝华,马学林,等.云南苹果树腐烂病菌分离株生物学特性和致病性研究[J].西南农业学报,2015,28(5):2 096-2 102.
GUI Teng-rong, KONG Bao-hua, MA Xue-li, et al.(2015).Studies on Biological Characters and Pathogencity of Isolates of Cytospora from Apple Tree in Yunnan [J].SouthwestChinaJournalofAgriculturalSciences, 28(5):2,096-2,102. (in Chinese)
[6] 王卫雄.甘肃省苹果树腐烂病菌种类、遗传多样性及生防细菌培养条件优化研究[D].兰州:甘肃农业大学硕士论文,2015.
WANG Wei-xiong.(2015).StudyofpathogendiversityandantagonisticbacteriaofthecausativeagentofappletreecankerinGansuProvince[D]. Master Thesis. Gansu Agricultural University, Lanzhou. (in Chinese)
[7] 刘钰娇.河北省苹果树腐烂病菌种类分析及分子检测体系的建立[D].保定:河北农业大学硕士论文,2015.
LIU Yu-jiao.(2015).SpeciesanalysisandestablishmentofmoleculardetectionsystemonapplevalsacankerpathogensinHebeiProvince[D].Master Thesis. Agricultural University of Hebei, Baoding. (in Chinese)
[8]唐俊煜.新疆绿洲生态枣园林木腐烂病菌种类鉴定及交互侵染机制研究[D].阿拉尔:塔里木大学硕士论文,2016.
TANG Jun-yu.(2016).ResearchonTheIdentificationandCrossInfectionMechanismoftheValsaCankerintheOasisEcologicalJujubeOrchardinXinjiang[D]. Master Thesis. Tarim University, Alar. (in Chinese)
[9] 陈元宏.杨柳树腐烂病发生规律及生态防治方法研究[J].黑龙江科技信息,2014,(26):268.
CHEN Yuan-hong.(2014).Study on Occurrence and Ecological Control of Willow Trees [J].HeilongjiangScienceandTechnologyInformation, (26):268. (in Chinese)
[10] 王雪,于菁妮,马现成,等.东北高寒地区果树腐烂病病原菌分离、检测与抑制探究[J].生物灾害科学, 2016, 39(4): 224-227.
WANG Xue,YU Jing-ni, MA Xian-cheng, et al.(2016). Separation, Detection and Suppression of Pathogenic Bacteria of Fruit Rot Disease in Heavy Frigid Regions in Northeast China [J].BiologicalDisasterScience, 39(4): 224-227. (in Chinese)
[11] 李春艳,张王斌,苟巧,等.不同寄主来源腐烂病菌对枣树致病性研究[J].新疆农业科学,2016,53(1): 51-58.
LI Chun-yan, ZHANG Wang-bin, GOU Qiao, et al. (2016).Study on the Pathogenicity ofCytosporaspp. from Different Hosts to Jujube [J].XinjiangAgriculturalSciences, 53(1): 51-58. (in Chinese)
[12] 殷辉,周建波, 张志斌,等.山西省苹果树腐烂病菌的种群结构分析[J].菌物学报,2016,35(12):1 493-1 502.
YIN Hui, ZHOU Jian-bo, ZHANG Zhi-bin, et al. (2016).Population structure of Valsa species from Malus pumila in Shanxi Province [J].Mycosystema, 35(12): 1,493-1,502. (in Chinese)
[13] 郭开发,吴彩兰,方晓翠,等.塔里木河下游胡杨树腐烂病病原鉴定[J].中国森林病虫,2015,34(5):1-4.
GUO Kai-fa, WU Cai-lan, FANG Xiao-cui, et al.(2015).Identification of canker pathogen of Populus euphratica in the lower reaches of Tarim River [J].ForestPestandDisease, 34(5):1-4. (in Chinese)
[14] 陆家元.植物病原真菌学[M].北京:中国农业出版社,2000:5.
LU Jia-yuan. (2000).Plantpathogenicmycology[M]. Beijing: China Agriculture Press: 5. (in Chinese)
[15] 范鑫磊.中国黄河流域壳囊孢属的分类和系统学研究[D].北京:北京林业大学,2016.
FAN Xin-lei. (2016).PhylogenyandTaxonomyofCytosporainYellowRiverRegionofChina[D]. Master Thesis. Beijing Forestry University, Beijing. (in Chinese)
[16] 张庆涛.中国壳囊孢属真菌分类与系统发育研究[D].上海:上海交通大学博士论文,2014.
ZHANG Qing-tao. (2014).TaxonomyandphylogenyofCytosporainChina[D]. PhD thesis. Shanghai Jiaotong University, Shanghai. (in Chinese)
[17] 冀瑞卿,李玉,宋瑞清,等.金黄壳囊孢菌rDNAITS序列测定及生理学特性[J].吉林农业大学学报,2010,32(5):483-487.
JI Rui-qing, LI Yu, SONG Rui-qing, et al. (2010). Cultural Characteristics and rDNA ITS Sequence Analysis of Cytospora chrysosperma [J].JournalofJilinAgriculturalUniversity, 32(5):483-487. (in Chinese)
IdentificationofWillowValsaCankersPathogenintheTarimRiverBasininSouthernXinjiang
SHAO Yan-hui, ZHANG Wang-bin, LIU Zhen-ya, ZHU Zong-cai, YAN Hai-lin,LI Ya-peng, CAO Qi, DAN Hong-xia
(TarimUniversity/SouthernXinjiangKeyLaboratoryofIPMofTarimUniversity,AlarXinjiang843300,China)
【Objective】 Through the study on pathogenic bacteria culture morphology and shape observation, combined with analysis of molecular biology techniques ITS and β-tubulin sequence analysis, this project aims to make clear how many kinds of willow valsa cander pathogens there are and their genetic differentiation in southern Xinjiang.【Method】Willow valsa cankers pathogen was collected and pure culture of pathogen was obtained. The conidia of the pathogen of pathogenic bacteria were divided into longitudinal and vertical sections and the morphological data were observed. Using ITS and β--tubulin sequence analysis method, the phylogenetic tree was successfully constructed. The most optimum pathogen was studied after β-tubulin sequential analysis.【Result】The pycnidium and conidia characteristics were consistent with the reportedCytosporachrysosperma. The most optimum conditions for the growth of pathogenic bacteria were the temperature between 20℃-30 ℃, the PH was 5-6, and the suitable carbon and nitrogen sources were glucose and peptone, respectively. ITS and β-tubulin analysis showed the sequence was up to 96% identical withCytosporaspp.; Pathogenic bacteria could infect 5 kinds of trees successfully in the in vitro inoculation conditions.【Conclusion】Biological observation and molecular sequence analysis showed that the pathogen causing willow valsa cankers in Tarim River basin of southern Xinjiang isCytosporachrysosperma. There are some genetic differentiations among pathogens from different sources. These pathogens can infect poplar, populus euphratica, jujube tree, pear and apple, and produce conidia in populus euphratica and poplar inside the room.
the willow valsa cankers in Tarim River basin in southern Xinjiang; morphological character; culture characteristics; ITS;β-tubulin
ZHANG Wang-bin (1974- ), male, Chengcheng in Shanxi, Associate Professor, Master Instructor.research field: Plant pathology. (E-mail)zwbzky@163.com
10.6048/j.issn.1001-4330.2017.08.014
2017-06-02
国家自然科学基金项目“新疆枣园林果树木腐烂病菌遗传多样性及致病力分化研究”(31660034);国家级大学生创新项目(201610757006)
邵延慧(1992-),女,硕士研究生,研究方向为果树优质高效栽培生理,(E-mail)18399576075@163.com
张王斌(1974-),男,陕西澄城人,副教授,硕士生导师,研究方向为植物病理学,(E-mail)zwbzky@163.com
S-43;S763.1
:A
:1001-4330(2017)08-1481-08
Supported by: National Natural Science Foundation of China "Genetic Diversity and Pathogenicity Differentiation in Rotting Jujube Trees in Xinjiang"(31660034)and National College Students Innovation Program (201610757006)
