黄淮麦区小麦种质资源矮秆基因分布及其与农艺性状的关系
2017-09-16周晓变陈建辉王永彦张香粉闫雪芳董中东崔党群
周晓变,赵 磊,陈建辉,阳 霞,王永彦,张香粉,闫雪芳,董中东,崔党群,陈 锋
(河南农业大学农学院/河南粮食作物协同创新中心/省部共建小麦玉米作物学国家重点实验室,河南郑州 450002)
黄淮麦区小麦种质资源矮秆基因分布及其与农艺性状的关系
周晓变,赵 磊,陈建辉,阳 霞,王永彦,张香粉,闫雪芳,董中东,崔党群,陈 锋
(河南农业大学农学院/河南粮食作物协同创新中心/省部共建小麦玉米作物学国家重点实验室,河南郑州 450002)
为了进一步阐明多个矮秆基因的分布及其与小麦农艺性状的关系,运用分子标记对来自我国黄淮麦区的246份小麦种质资源中6个矮秆基因位点( Rht1、 Rht2、 Rht4、 Rht8、 Rht9及 Rht12)分别进行了检测,同时连续3年调查参试材料株高、穗长、穗下节长、小穗数、旗叶长、旗叶宽、穗粒数、粒长、粒宽和千粒重共10个农艺性状,分析了不同矮秆基因位点对小麦农艺性状的影响。结果表明,6个矮秆基因在黄淮麦区小麦中均具有广泛分布,其中含有 Rht1和 Rht2基因的小麦品种分布最广。分析矮秆基因位点对小麦农艺性状的影响发现,在 Rht1位点, Rht1-B1a和 Rht1-B1b两种基因型间的株高没有显著差异;在 Rht2位点,拥有 Rht2-D1b类型的小麦品种所有年份间的株高和穗下节长较低,但千粒重较高,为相对优良的基因型。排除 Rht1和 Rht2基因效应后, Rht4、 Rht8、 Rht9和 Rht12位点对黄淮麦区小麦品种不同农艺性状均具有重要影响,其中, Rht4基因位点主要对小麦株高和千粒重具有重要影响,且 Rht4-B1b类型为相对优良的基因型; Rht8基因位点主要对小麦穗下节长、穗长和千粒重具有重要影响,且 Rht8-D1b类型为相对优良的基因型; Rht9基因位点主要对小麦株高和千粒重具有重要影响,且 Rht9-A1a类型为相对优良的基因型; Rht12基因位点主要对小麦千粒重和穗长具有重要影响,且 Rht12-A1a类型为相对优良的基因型。进一步分析发现,6个位点中对株高影响最大的是 Rht2基因,其次是 Rht4基因;有4个位点( Rht1、 Rht2、 Rht8、 Rht12)对千粒重有显著影响,其中 Rht2基因的影响最大。分析除 Rht1外其他5个位点优良基因型在不同时期小麦品种中的分布发现,从早期历史品种、近期历史品种到现代品种,不同位点优良基因型分布比例总体呈现上升趋势,表明优良矮秆基因型在黄淮麦区小麦品种选育中的利用逐渐增加,尤其是82.9%的现代小麦品种已含有 Rht2-D1b类型。
普通小麦;矮秆基因;分子标记;农艺性状;优良基因型
自20世纪60年代诺曼布劳格发起第一次“绿色革命”[1],国内外相关学者对小麦矮秆基因的研究越来越多,且矮化育种已成为小麦高产育种的一个重要途径。近年来,分子标记的迅速发展为重要功能基因的鉴定和筛选提供了便利,能够缩短育种年限,加快育种进程。因此,分子标记辅助选择与常规育种有机结合将成为今后小麦育种的主要发展方向。迄今为止,已报道和命名大量矮秆基因[2-3],但是对生产影响较大且较常用的主要有 Rht1( Rht1-B1b)、 Rht2( Rht2-D1b)、 Rht3、 Rht4、 Rht8、 Rht9、 Rht10、 Rht12和 Rht17[4-6]。其中, Rht1( Rht1-B1b)和 Rht2( Rht2-D1b)是在农林10号中被发现,在矮化育种中应用最广,分别定位于4BS与4DS染色体上[7-8]。进一步研究表明[9], Rht1和 Rht2位点均包含多种等位变异,但主要有 Rht1-B1a和 Rht1-B1b、 Rht2-D1a和 Rht2-D1b,其中,拥有 Rht1-B1b和 Rht2-D1b类型的小麦品种株高较矮,综合农艺性状相对优良。Ellis等[10]开发出了 Rht-B1b和 Rht-D1b的功能标记,可以明确的区分出 Rht-B1a和 Rht-B1b及 Rht-D1a和 Rht-D1b。 Rht4是从小麦品种Burt的后代BurtM937中鉴定出来的,定位在2BL染色体上,与标记WMC317紧密连锁,在Vigour 18的后代中扩增出大小为150 bp的条带,在Burt的后代中扩增出大小为170 bp的条带[11]。 Rht8和 Rht9均来源于赤小麦,其中, Rht8位于2DL染色体上,与标记WMS261紧密连锁[12-13]; Rht9位于5AL染色体上,与标记BARC151紧密连锁,在川麦18中可以检测到大小为230 bp的条带,在Mara中检测到大小为220 bp的条带[11]。 Rht12来源于品种Karcagi的辐射突变体,位于5AL染色体上,与标记WMC410紧密连锁,在Vigour 18的后代中扩增出大小为112 bp的条带,在Mercia 12的后代中扩增出大小为114 bp的条带[14]。国内外多名学者对不同矮秆基因的作用进行了分析,Ellis等[15]发现, Rht1-B1b、 Rht2-D1b、 Rht4、 Rht9和 Rht12在植物的早期生长过程中均能显著降低株高。Asghar等[16]发现, Rht8对株高和其他农艺性状均具有一定影响。Akman等[17]和Wang等[18]发现, Rht2-D1b和 Rht8分别能够降低小麦株高的16%~30%和3.8%~12%,同时 Rht2-D1b能够显著降低植株节间长度。多个研究表明, Rht12既能显著降低小麦株高与旗叶长,又与小麦开花期有关[19-21]。Guedira等[22]发现,美国软粒冬小麦品种中, Rht1-B1b和 Rht2-D1b的分布频率分别为28%和45%。Mccartney等[23]对小麦重要农艺性状QTL定位发现,矮秆基因 Rht1-B1b和 Rht2-D1b与穗粒数和千粒重等农艺性状的QTL位点相重合。综上可知, Rht1-B1b与 Rht2-D1b除了对株高有影响外,对小麦千粒重等农艺性状也有影响。
关于我国小麦品种的主要矮秆基因型分布的研究也取得了一定的进展。Zhang等[24]研究表明, Rht2-D1b和 Rht8在我国小麦品种中的分布频率分别为45.5%与46.8%,是最为常见的2种基因型,而 Rht1-B1b所占频率较低,仅为24.5%。Yang等[25]也发现,我国小麦品种中 Rht2-D1b的分布比例显著大于 Rht1-B1b。林旭群等[26]和许 琦等[27]分别分析了 Rht1-B1b、 Rht2-D1b和 Rht8对株高的影响,结果表明, Rht2-D1b的降秆作用最大, Rht1-B1b次之, Rht8降秆作用最小,且3个基因的联合降秆作用显著大于2个基因,2个基因作用显著大于单个基因。
前人研究大多数是对主要矮秆基因( Rht1-B1b、 Rht2-D1b和 Rht8等)在不同类型小麦品种中的分布进行检测和效应分析,本试验对黄淮麦区不同时期的小麦种质资源中6个矮秆基因位点( Rht1、 Rht2、 Rht4、 Rht8、 Rht9及 Rht12)进行了检测,并对不同矮秆基因型与包括株高在内的多个小麦农艺性状的关系进行多年度分析,以期阐明不同矮秆基因位点对小麦农艺性状的影响,并筛选出农艺性状相对优良的矮秆基因型组合,为黄淮麦区进一步合理利用矮秆基因资源和培育高产小麦新品种提供参考。
1 材料与方法
1.1 试验材料
本试验所用材料为来自于我国黄淮麦区当前主栽小麦品种、历史品种和农家品种,共计246份。所有材料分别于2012-2013、2013-2014和2014-2015年度种植于河南农业大学郑州科教园区,田间四周设保护行,采用完全随机排列,常规田间管理。每个品种种植2行,行长2 m,行间距20 cm,株间距10 cm。农家品种采用防倒网支撑,所有材料均未有倒伏发生。小麦抽穗后(4月中下旬),每个品种选取10株均匀一致、具有代表性的植株,在收获前调查株高、穗长、穗下节长、可孕小穗数、不孕小穗数、旗叶长及旗叶宽(2013年收获的材料未调查旗叶长和宽)。成熟后,选取上述标记的单株进行收获、脱粒及室内考种,分别测量粒长(10粒总长)、粒宽(10粒总宽)、穗粒数及千粒重。
1.2 DNA提取及PCR扩增
每个品种取3粒有代表性的种子,参照Chen等[28]的方法分别提取其基因组DNA。利用1.2%的琼脂糖凝胶电泳以及紫外分光光度计检测DNA的质量和浓度,并根据检测结果将样品DNA稀释至浓度为50 ng·μL-1的工作液,置于-20 ℃条件下保存备用。
用于鉴定 Rht1-B1a和 Rht1-B1b以及 Rht2-D1a和 Rht2-D1b的引物按照Ellis等[10]提供的序列进行合成。用于鉴定 Rht8与 Rht12的引物按照Korzun等[12,14]报道的序列进行合成。用于鉴定 Rht4与 Rht9位点的引物参照Ellis[11]公布的序列进行合成。上述具体引物序列见表1。
PCR反应体系:10×Taqbuffer 2.5 μL,10 mmol·L-1的dNTPs 0.5 μL,10 pmol·μL-1的引物各0.4 μL,2.5 U·μL-1的TaqDNA聚合酶0.4 μL,DNA模板2.0 μL,加ddH2O补足至25 μL。PCR扩增程序:95 ℃预变性5 min;94 ℃变性30 s,57~63 ℃退火30 s(详见表1),72 ℃延伸30 s,38个循环;72 ℃延伸10 min。扩增产物用1.5%的琼脂糖凝胶进行电泳检测,150 V的电压下电泳30 min,经溴化乙锭染色,在凝胶成像系统扫描仪下观察并照相。
1.3 统计分析
利用Excel 2007对不同基因型农艺性状进行描述性统计及作图,利用SPSS 19.0对不同基因型农艺性状进行差异显著性分析。
2 结果与分析
2.1 黄淮麦区小麦品种不同位点矮秆基因型分布
各位点的检测结果表明,共有242份材料含有 Rht1基因,其中,有140份材料为 Rht1-B1a 类型,占供试材料的56.19%,有102份材料为 Rht1-B1b类型,占供试材料的41.46%。共有242份材料含有 Rht2基因,其中,有86份材料为 Rht2-D1a类型,占供试材料的34.96%,有156份材料为 Rht2-D1b类型,占供试材料的63.41%。共有198份材料含有 Rht4基因。依据其扩增出的两条不同带型,进一步将 Rht4基因型分别命名为 Rht4-B1a(低带型:150 bp)和 Rht4-B1b(高带型:170 bp),其中,有164份材料为 Rht4-B1b类型,占供试材料的66.67%,有34份材料为 Rht4-B1a类型,占供试材料的13.82%。共有175份材料含有 Rht8基因。依据其扩增出的两条不同带型,进一步 Rht8基因型分别命名为 Rht8-D1a(低带型:165 bp)和 Rht8-D1b(高带型:192 bp),其中,有69份材料是 Rht8-D1a类型,占供试材料的28.05%,有106份材料是 Rht8-D1b类型,占供试材料的43.09%。共有183份材料含有 Rht9基因。依据其扩增出的两条不同带型,进一步将 Rht9基因型分别命名为 Rht9-A1a(低带型:220 bp)和 Rht9-A1b(高带型:230 bp),其中,有100份材料为 Rht9-A1a类型,占供试材料的40.65%,有83份材料为 Rht9-A1b类型,占供试材料的33.74%。共有191份材料含有 Rht12基因。依据其扩增出的两条不同带型,进一步将 Rht12基因型分别命名为 Rht12-A1a(低带型:114 bp)和 Rht12-A1b(高带型:210 bp),其中,有106份材料为 Rht12-A1a类型,占供试材料的43.09%,有85份材料为 Rht12-A1b类型,占供试材料的34.55%。

表1 检测不同位点矮秆基因的引物序列及扩增条件Table 1 Primer sequences and their amplification conditions for identification of Rht genes in bread wheat
综上所述,在246份供试材料中,有4份农家种小麦未发现任何矮秆基因(6个位点),但有111份材料在这6个矮秆基因位点均呈现出植株相对较矮的等位基因型,占总供试材料的45.12%,表明矮秆基因在黄淮麦区小麦品种中已有广泛应用。
2.2黄淮麦区小麦品种不同位点矮秆基因型对农艺性状的影响
由表2可以看出,在 Rht1位点, Rht1-B1a与 Rht1-B1b类型的小麦品种在株高等多个农艺性状上均没有显著差异。在 Rht2位点, Rht2-D1a类型小麦品种的株高和穗下节长在所有年份均显著高于 Rht2-D1b类型,而千粒重却显著低于 Rht2-D1b类型; Rht2-D1a类型小麦品种的旗叶长在部分年份显著高于 Rht2-D1b类型,而粒长和粒宽却显著低于 Rht2-D1b类型。在 Rht4位点, Rht4-B1a类型小麦品种的株高、穗下节长、穗粒数与旗叶长在部分年份显著高于 Rht4-B1b类型。在 Rht8位点, Rht8-D1a类型小麦品种的穗长在所有年份均显著高于 Rht8-D1b类型,而穗粒数、株高、旗叶长仅在部分年份显著高于 Rht8-D1b类型; Rht8-D1a类型小麦品种的千粒重在部分年份显著低于 Rht8-D1b类型。在 Rht9位点, Rht9-A1a与 Rht9-A1b类型小麦品种各农艺性状差异在所有年份均不显著。在 Rht12位点, Rht12-A1a类型小麦品种的穗长在所有年份均显著低于 Rht12-A1b类型,而小穗数和旗叶长仅在部分年份显著低于 Rht12-A1b类型; Rht12-A1a类型小麦品种的粒宽和千粒重在部分年份显著高于 Rht12-A1b类型。未发现任何矮秆基因(6个位点)的小麦品种的株高、穗长及穗下节长在所有年份间均显著高于其他含有矮秆基因的小麦品种,小穗数(除2013年外)、穗粒数和千粒重均显著低于其他含有矮秆基因的小麦品种。
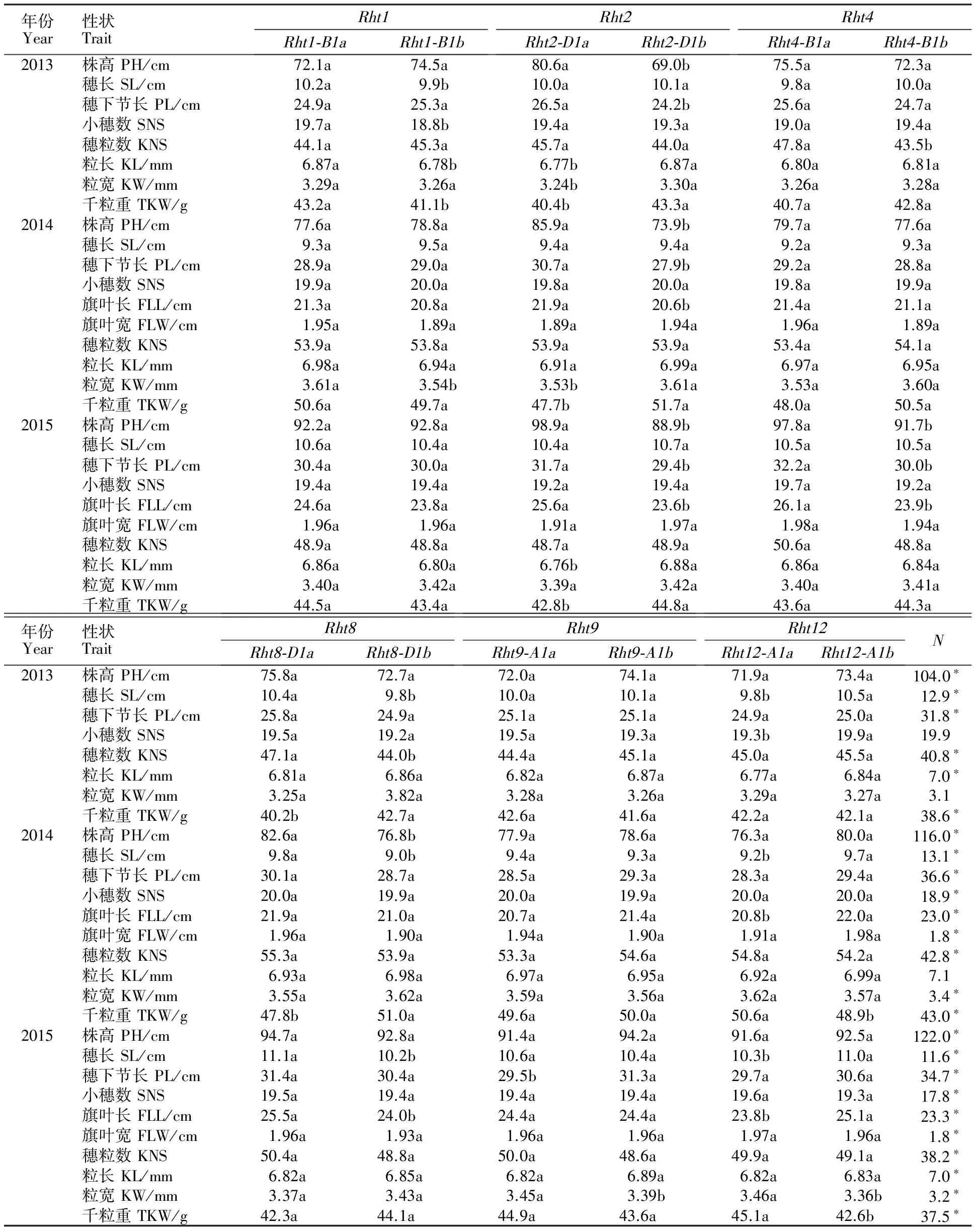
表2 不同位点矮秆基因型间农艺性状的比较Table 2 Comparison of agronomic traits of cultivars with different Rht genes
N:所检测的6个位点不含矮秆基因;数据后不同字母表示等位变异的农艺性状差异在0.05水平上显著;*表示与含有矮秆基因小麦品种的农艺性状差异在0.05水平上显著。下同。
N:No dwarf gene at the six loci; Different letters following data show the difference of agronomic traits was significant at 5% probability level between allelic variations;* indicated the difference of agronomic traits was significant at 5% probalility level among the cultivars not containing dwarf genes and the cultivars containing dwarf genes;PH:Plant height; SL:Spike length; PL:Peduncle length; SNS:Spikelet number per spike; FLL:Flag leaf length; FLW:Flag leaf width; KNS:Kernel number per spike; KL:Kernel length; KW:Kernel width; TKW:Thousand-kernel weight.The same below.
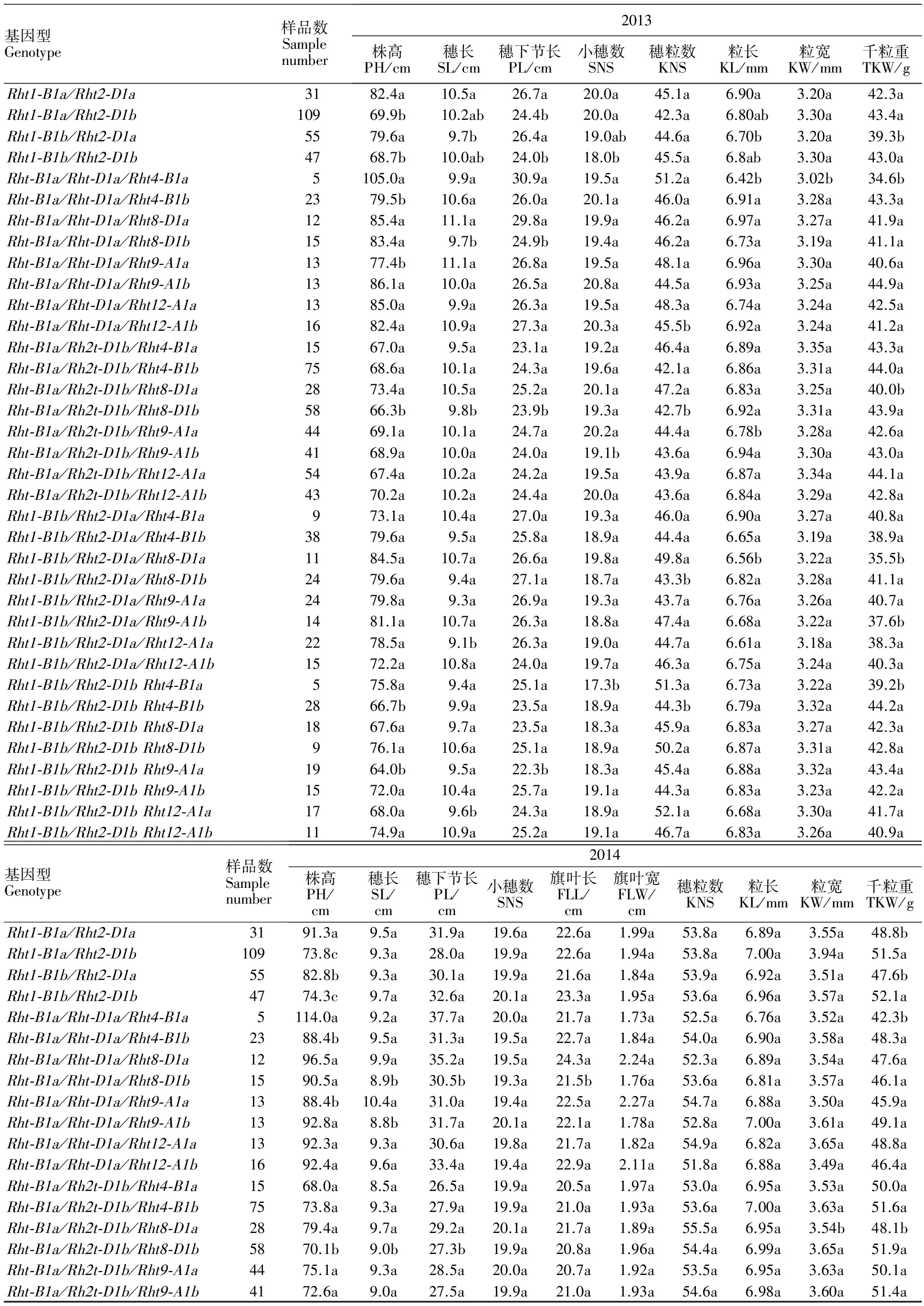
表3 各年份不同位点矮秆基因组合间农艺性状的比较Table 3 Comparison of agronomic traits of cultivars with different Rht gene combinations in three years
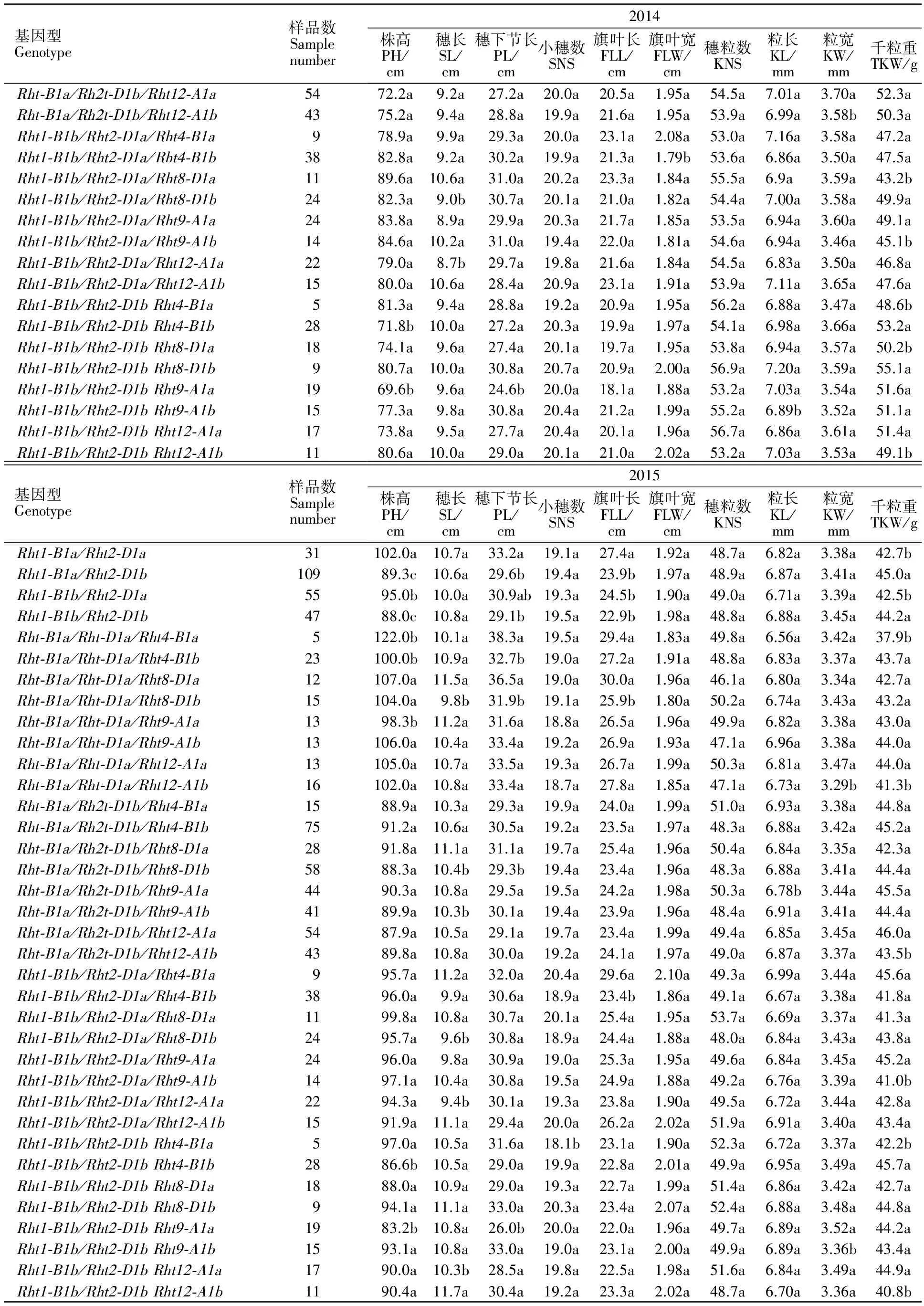
(续表3 Continued table 3)
从以上结果可知, Rht2、 Rht4、 Rht8对参试的黄淮麦区小麦品种株高均有一定影响,其中,影响最明显的是 Rht2基因; Rht1、 Rht2、 Rht8、 Rht12对参试的黄淮麦区小麦品种千粒重均有一定影响,其中,影响最明显的也是 Rht2基因。
2.3Rht1/Rht2基因组合对农艺性状的影响
本研究将 Rht1与 Rht2位点不同基因型进行组合,共得到 Rht1-B1a/ Rht2-D1a、 Rht1-B1a/ Rht2-D1b、 Rht1-B1b/ Rht2-D1a及 Rht1-B1b/ Rht2-D1b四种组合类型。其中 Rht1-B1a/ Rht2-D1b组合类型所占比例较高,共有109份,占45.04%;其次为 Rht1-B1b/ Rht2-D1a类型,共55份,占22.73%; Rht1-B1b/ Rht2-D1b和 Rht1-B1a/ Rht2-D1a类型分别为47份和31份,所占比例分别为19.42%和12.81%。进一步分析4种不同组合类型对黄淮麦区小麦农艺性状的影响,结果(表3)表明,各年份部分基因型的株高、穗长、穗下节长、小穗数、粒长、旗叶长、千粒重之间具有显著差异。其中, Rht1-B1b/ Rht2-D1b和 Rht1-B1a/ Rht2-D1b类型小麦品种的株高在调查的所有年份均显著低于 Rht1-B1b/ Rht2-D1a与 Rht1-B1a/ Rht2-D1a类型小麦品种, Rht1-B1a/ Rht2-D1a类型小麦品种三年间均表现出最高的株高; Rht1-B1b/ Rht2-D1b和 Rht1-B1a/ Rht2-D1b类型小麦品种的千粒重在所有年份间均高于 Rht1-B1b/ Rht2-D1a与 Rht1-B1a/ Rht2-D1a类型小麦品种,其中在2014和2015年度差异均达显著水平。
2.4Rht1和Rht2位点基因型相同的情况下Rht4、Rht8、Rht9和Rht12位点对黄淮麦区小麦品种农艺性状的影响
由前人研究得知, Rht1和 Rht2的效应较大(但在本试验 Rht1效应较小),为了进一步分析去除掉 Rht1和 Rht2效应之后 Rht4、 Rht8、 Rht9和 Rht12位点对黄淮麦区小麦品种农艺性状的影响,本研究分析了具有相同 Rht1/ Rht2组合但 Rht4、 Rht8、 Rht9或 Rht12位点基因型不同的小麦品种农艺性状间的差异,结果详见表3。 Rht4位点的分析结果表明,在 Rht1-B1a/ Rht2-D1a和 Rht1-B1b/ Rht2-D1b 两个基因组合位点, Rht4-B1b类型小麦品种株高在所有年份间均显著低于 Rht4-B1a类型的品种,但前者的千粒重在所有年份间则均显著高于后者;另外,在 Rht1-B1a/ Rht2-D1a基因型组合位点,两种基因型小麦品种部分年份间的粒长、粒宽和穗下节长存在显著差异;在 Rht1-B1b/ Rht2-D1b基因型组合位点,两种基因型小麦品种部分年份间的小穗数、穗粒数存在显著差异。 Rht8位点分析的结果表明,在 Rht1-B1a/ Rht2-D1a和 Rht1-B1a/ Rht2-D1b 两个基因组合位点, Rht8-D1a类型小麦品种穗下节长和穗长在所有年份间均显著高于 Rht8-D1b类型的品种;在 Rht1-B1a/ Rht2-D1b、 Rht1-B1b/ Rht2-D1a和 Rht1-B1b/ Rht2-D1b 三个基因组合位点, Rht8-D1b类型小麦品种千粒重在所有年份均高于 Rht8-D1a类型的品种,其中大部分年份间差异达到了显著水平。 Rht9位点分析结果表明,在 Rht1-B1a/ Rht2-D1a和 Rht1-B1b/ Rht2-D1b 两个基因组合位点, Rht9-A1a类型小麦品种株高在所有年份间均显著低于 Rht9-A1b类型的品种;在 Rht1-B1b/ Rht2-D1a基因组合位点, Rht9-A1a类型小麦品种千粒重在所有年份间均显著高于 Rht9-A1b类型的品种。 Rht12位点分析结果表明,在 Rht1-B1a/ Rht2-D1a、 Rht1-B1a/ Rht2-D1b和 Rht1-B1b/ Rht2-D1b基因组合位点, Rht12-A1a类型小麦品种千粒重在所有年份间均高于 Rht12-A1b类型的品种,且在2015年度达到了显著水平;另外,在 Rht1-B1b/ Rht2-D1a和 Rht1-B1b/ Rht2-D1b基因组合位点, Rht12-A1a类型小麦品种穗长均短于 Rht12-A1b类型的品种,且在2013和2015年度达到了显著水平。
因此,可以看出,排除 Rht1和 Rht2基因效应后, Rht4、 Rht8、 Rht9和 Rht12位点对黄淮麦区小麦品种不同农艺性状均具有重要影响,其中 Rht4基因位点主要对小麦株高和千粒重具有重要影响,且 Rht4-B1b类型为相对优良的基因型; Rht8基因位点主要对小麦穗下节长、穗长和千粒重具有重要影响,且 Rht8-D1b类型为相对优良的基因型; Rht9基因位点主要对小麦株高和千粒重具有重要影响,且 Rht9-A1a类型为相对优良的基因型; Rht12基因位点主要对小麦千粒重和穗长具有重要影响,且 Rht12-A1a类型为相对优良的基因型。
2.5 不同矮秆基因位点优良基因型分布
根据上述分析结果,可以看出不同矮秆基因位点的相对优良基因型分别为 Rht2-D1b、 Rht4-B1b、 Rht8-D1b、 Rht9-A1a和 Rht12-A1a。依据参试小麦品种审定时间,分别将其分为早期历史品种(1980年之前审定)、近期历史品种(1980-2000之间审定)和现代品种(2000之后审定),其样品数分别为40份、107份和70份,剩余材料为农家种或引进品种,因为数目较少,未作更多分析。进一步分析5个矮秆位点优良基因型在不同时期小麦品种中的分布,结果(图1)表明,从早期历史品种、近期历史品种到现代品种,5个位点优良基因型分布比例大多呈现上升趋势,尤其是 Rht2-D1b类型分布比例上升幅度最大,从48.8%(早期历史品种)、59.8%(近期历史品种)到82.9%(现代品种),表明5个矮秆基因位点在黄淮麦区小麦品种的利用均在逐渐增加,尤其是 Rht2位点的优良基因型在黄淮麦区的绝大多数现代品种中均已含有。
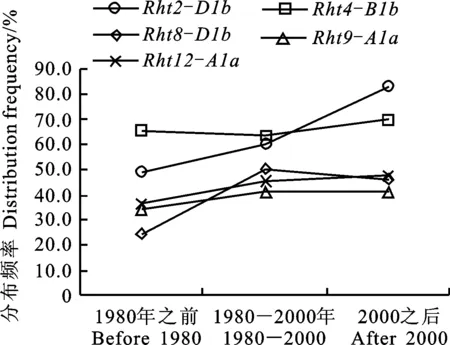
图1 不同时期小麦品种中5个矮秆基因位点优良基因型的分布频率
3 讨 论
自20世纪60年代农林10号中 Rht1( Rht1-B1b)和 Rht2( Rht2-D1b)基因运用到小麦育种并取得显著效果以来,矮秆基因的研究在全世广泛开展。矮秆在小麦育种中的推广和应用,不仅降低了小麦株高,还大幅度提高了小麦产量。随着矮秆基因研究的逐步深入,越来越多的矮秆基因被定位和克隆[9,11,29],但前人研究多集中在对 Rht1和 Rht2的研究,本试验则全面地分析了 Rht1、 Rht2、 Rht4、 Rht8、 Rht9与 Rht12这6个位点矮秆基因在黄淮麦区小麦中的分布及其对多个农艺性状的影响。据Evans等[30]调查,全世界小麦品种中约有70%以上携带有 Rht1-B1b或 Rht2-D1b。我国小麦品种中,矮秆基因分布较广的是 Rht1-B1b、 Rht2-D1b及 Rht8[31-32]。前人关于 Rht4、 Rht9与 Rht12这三个位点的基因研究相对较少。因此,进一步更为全面地了解不同矮秆基因的分布,并阐明其对小麦农艺性状的影响,可为高产小麦育种提供有价值的信息。本试验检测的246份小麦品种不同位点矮秆基因分布表明,携带 Rht1与 Rht2的小麦品种分布最为广泛,6个位点均不含有矮秆基因的小麦品种分布比例最低,主要集中在1980年之前审定的早期历史品种。Zhang等[24]和慕美财等[34]研究发现, Rht1与 Rht2位点的 Rht1-B1b与 Rht2-D1b在我国小麦品种中分布均较为广泛。本试验除了发现 Rht1与 Rht2基因在我国黄淮麦区小麦品种中具有广泛分布外,还发现 Rht4、 Rht8、 Rht9与 Rht12基因在黄淮麦区小麦品种中也具有广泛分布。
通过分析不同位点矮秆基因对黄淮麦区小麦品种农艺性状的影响,结果发现,对株高影响最大的是 Rht2基因,其次是 Rht4基因。进一步分析其对千粒重的影响,发现所调查的6个矮秆基因位点中,有4个位点( Rht1、 Rht2、 Rht8、 Rht12)对黄淮麦区小麦千粒重有显著影响(部分年份间差异达到显著水平),携带以上位点优异基因型的材料至少一年的千粒重差异达到显著水平,其中, Rht2基因的影响最大,调查的所有年份间差异均达到了显著水平。同时发现, Rht4基因主要对小麦株高和千粒重具有重要影响, Rht8基因主要对小麦穗下节长、穗长和千粒重具有重要影响, Rht12基因主要对小麦千粒重和穗长具有重要影响,且相对优良基因型分别为 Rht2-D1b、 Rht4-B1b、 Rht8-D1b 和 Rht12-A1a。因此, Rht4、 (〗Rht8、)〗 Rht12可能是直接控制株高的微效基因或者与控制株高的主效基因产生互作进而间接影响株高,其调控机制有待进一步深入研究。分析优良基因型在不同时期小麦品种中的分布可以得出,大多数优良基因型在黄淮麦区小麦品种中的利用频率从早期历史品种到现代品种均呈现上升趋势,还发现黄淮麦区现代小麦品种中绝大多数品种已经拥有 Rht2-D1b(82.9%)和 Rht4-B1b(70.0%)两种优良基因型。因此,今后对黄淮麦区小麦品种产量相关性状进行改良时,应优先考虑效应相对较大的 Rht2-D1b和 Rht4-B1b两种基因型,但由于绝大多数小麦品种已经含有这两种基因型,所以还应进一步考虑利用 Rht8-D1b 和 Rht12-A1a这两种优良基因型。同时,在黄淮麦区小麦育种中,若以提高千粒重为主要育种目标,可优先重点考虑利用 Rht2-D1b、 Rht1-B1a、 Rht8-D1b和 Rht12-A1a四种优良基因型;若以降低株高为主要改良目标,可主要考虑 Rht2-D1b 和 Rht4-B1b两种优良基因型;若以增加穗长为主要育种目标,可主要考虑 Rht8-D1b 和 Rht12-A1a两种优良基因型。
由于不同基因间常常存在互作效应,本研究还分析了 Rht1和 Rht2矮秆基因组合的分布及其对农艺性状的影响,发现 Rht1-B1a/ Rht2-D1b组合类型在黄淮麦区小麦品种中分布频率最高,达到了45.0%,其余三种组合类型 Rht1-B1a/ Rht2-D1a、 Rht1-B1b/ Rht2-D1a、 Rht1-B1b/ Rht2-D1b尽管也有一定分布,但比例分别仅为12.8%、22.7%和19.4%,这与梁 丹等[35]的研究结果相一致。这可能是由于 Rht1-B1a和 Rht2-D1b分别在 Rht1和 Rht2位点是相对优良的基因型,而在小麦育种过程中这两种优良基因型经历人工选择造成的。 Rht1-B1a/ Rht2-D1b与 Rht1-B1b/ Rht2-D1b类型的小麦品种具有更低的株高和更高的千粒重,具有相对优良的综合农艺性状,可考虑在黄淮麦区小麦育种中利用。
由于农艺性状受环境影响较大,不同地区或单位的育种目标也不尽相同,本试验研究分析了6个矮秆基因位点对黄淮麦区小麦品种农艺性状的影响,并找到了相对优良的优异基因型或基因型组合,但在针对具体需要改良的农艺性状时,还需要结合田间表型及其他基因位点的效应进行综合选择。
[1]KHUSH G S.Green revolution:The way forward [J].NatureReviewsGenetics,2001,2:815.
[2]MCINTOSH R A,DUBCOVSKY J,ROGERS W J,etal.Catalogue of gene symbols for wheat.2013-2014 supplement [ER/OL].[2015-04-09](2016-10-10).http://www.shigen.nig.ac.Jp/wheat/komugi/genes/symbolClassList.jsp.
[3]CHEN S L,GAO R H,WANG H Y,etal.Characterization of a novel reduced height gene ( Rht23) regulating panicle morphology and plant architecture in bread wheat [J].Euphytica,2015,203:583.
[4]ELLIS M H,REBETZKE G J,CHANDLER P,etal.The effect of different height reducing genes on the early growth of wheat [J].FunctionalPlantBiology,2004,31:583.
[5] 赵 和.小麦矮秆基因研究和利用性状[J].河北农业科学,2008,8(4):96.
ZHAO H.Advances in researches and applications of dwarfing genes in wheat [J].JournalofHebeiAgriculturalSciences,2008,8(4):96.
[6]CASEBOW R,HADLEY C,UPPAL R,etal.Reduced height(Rht) alleles affect wheat grain quality [J].PLoSOne,2016,11(5):e0156056.
[7]GALE M D,LAW C N,WORLAND A J.The chromosomal location of a major dwarfing gene from Norin 10 in British semi-dwarf wheats [J].Heredity,1975,35:417.
[8]GALE M D,MARSHALL G A.The chromosomal location ofGailand Rht1 genes for gibberellins insensitivity and semi-dwarfism,in a derivative of Norin 10 wheat [J].Heredity,1976,37:283.
[9]PENG J R,RICHARDS D E,HARTLEY N H,etal.‘Green revolution’ genes encode mutant gibberellin response modulators [J].NatureReviewsGenetics,1999,400(6731):256.
[10]ELLIS M H,SPIELMEYER W,GALE R,etal.“Perfect” markers for the Rht-B1b and Rht-D1b dwarfing genes in wheat [J].TheoreticalandAppliedGenetics,2002,105(6):1038.
[11]ELLIS M H,REBETZKE G J,AZANZA F.Molecìlar mapping of gibberell in-responsive dwarfing genes in bread wheat [J].TheoreticalandAppliedGenetics,2005,111:423.
[12]KORZUN V,RODER M S,GANAL M W,etal.Genetic analysis of the dwarfing gene (Rht8) in wheat.Part I.Molecular mapping of Rht8 on the short arm of chromosome 2D of bread wheat (TriticumaestivumL.) [J].TheoreticalandAppliedGenetics,1998,96:1104.
[13]WORLAND A J,KORZUN V,RODER M S,etal.Genetic analysis of the dwarfing gene( Rht8) in wheat.Part II.The distribution and adaptive significance of allelic variants at the Rht8 locus of wheat as revealed by microsatellite screening [J].TheoreticalandAppliedGenetics,1998,96:1110.
[14]KORZUN V,RODER M,WORLAND A J,etal.Intrachromosomal mapping of genes for dwarfing ( Rht1) and vernalization response ( Vrn1) in wheat using RFLP and microsatellite markers [J].PlantBreeding,1997,116:227.
[15]ELLIS M H,REBETZKE G J,CHANDLER P,etal.The effect of different height reducing genes on the early growth of wheat [J].FunctionalPlantBiology,2004,31:583.
[16]ASGHAR B G,LI C,HU Y G.The effects of dwarfing gene Rht-8 on plant height and other agronomic traits in common wheat [J].AgriculturalResearchintheAridAreas,2014,32(1):252.
[17]AKMAN H,BRUCKNER P.Marker assisted selection for Rht8 and Rht-D1b dwarfing genes in winter wheat breeding program [J].NewBiotechnology,2012,29:S139.
[18]WANG Y S,CHEN L,DU Y Y,etal.Genetic effect of dwarfing gene Rht13 compared with Rht-D1b on plant height and some agronomic traits in common wheat (TriticumaestivumL.) [J].FieldCropsResearch,2014,162:39.
[19]WORLAND A J,SAYERS E J,BORNER A.The genetics and breeding potential of Rht12,a dominant dwarfing gene in wheat [J].PlantBreeding,1994,113(3):187.
[20]WOJCIECHOWSKI T,GOODING M J,RAMSAY L,etal.The effects of dwarfing genes on seedling root growth of wheat [J].JournalofExperimentalBotany,2009,60(9):2565.
[21]CHEN L,PHILLIPS A L,CONDON A G,etal.GA-responsive dwarfing gene Rht12 affects the developmental and agronomic traits in common bread wheat [J].PloSOne,2013,8(4):e62285.
[22]GUEDIRA M,BROWN-GUEDIRA G,VAN SANFORD D,etal.Distribution ofRhtgenes in modern and historic winter wheat cultivars from the eastern and central USA [J].CropScience,2010,50(5):1811.
[23]MCCARTNEY C A,SOMERS D J,LUKOW O,etal.QTL analysis of quality traits in the spring wheat cross RL4452בAC Domain’ [J].PlantBreeding,2007,11(29):2607.
[24]ZHANG X K,YANG S J,ZHOU Y,etal.Distribution of the Rht-B1b, Rht-D1b and Rht8 reduced height genes in autumn-sown Chinese wheats detected by molecular markers [J].Euphytica,2006,152:109.
[25] 杨松杰,张晓科,何中虎,等.用STS标记检测矮秆基因 Rht-B1b和 Rht-D1b在中国小麦中的分布[J].中国农业科学,2006,39(8):1680.
YANG S J,ZHANG X K,HE Z H,etal.Distribution of dwarfing genes Rht-B1b and Rht-D1b in Chinese bread wheats detected by STS marker [J].ScientiaAgriculturaSinica,2006,39(8):1680.
[26] 林旭群,李韵芳,鲁 璐,等.小麦矮秆基因 Rht-B1b、 Rht-D1b、 Rht8的检测及其对株高的影响[J].应用与环境生物学报,2014,20(1):80.
LIN X Q,LI Y F,LU L,etal.Identification of wheat dwarfing genes Rht-B1b, Rht-D1b, Rht8 and their effects on plant height [J].ChineseJournalofAppliedandEnvironmentalBiology,2014,20(1):80.
[27] 许 琦,杨 娜,柴永峰,等.中国小麦主要矮秆基因的分布及其对株高的影响[J].西北农业学报,2014,23(5):59.
XU Q,YANG N,CHAI Y F,etal.Distribution and impact on plant height of major wheat dwarfing genes in China [J].ActaAgriculturaeBoreali-occidentalisSinica,2014,23(5):59.
[28]CHEN F,XU H X,ZHANG F Y,etal.Physical mapping of puroindoline b-2 genes and molecular characterization of a novel variant in durum wheat(TriticumturgidumL.) [J].MolecularBreeding,2011,28:153.
[29]PEARCE S,SAVILL R,VAUGHAN S P,etal.Molecular characterization of Rht-1 dwarfing genes in hexaploid wheat [J].PlantPhysiology,2011,157(4):1820.
[30]EVANS L T.Feeding the ten billion:Plant and population growth [J].TheQuarterlyReviewofBiology,2000,75:2.
[31] 郭保宏,宋春华,贾继增.我国小麦品种的 Rht1、 Rht2矮秆基因鉴定及分布研究[J].中国农业科学,1997,30(5):56.
GUO B H,SONG C H,JIA J Z.Distribution and detection of dwarf genes Rht1 and Rht2 in wheat in China [J].ScientiaAgriculturaSinica,1997,30(5):56.
[32] 唐 娜,李 博,闵 红,等.分子标记检测矮秆基因 Rht-B1b和 Rht-D1b及 Rht8在我国小麦中的分布[J].中国农业大学学报,2012,17(4):21.
TANG N,LI B,MIN H,etal.Distribution of dwarfing genes Rht-B1b, Rht-D1b and Rht8 in Chinese breed wheat cultivars detected by molecular markers [J].JournalofChinaAgriculturalUniversity,2012,17(4):21.
[33] 张德强,宋晓朋,冯 洁,等.黄淮麦区小麦品种矮秆基因 Rht-B1b、 Rht-D1b和 Rht8的检测及其对农艺性状的影响[J].麦类作物学报,2016,36(8):975.
ZHANG D Q,SONG X P,FENG J,etal.Detection of dwarf genes Rht-B1b, Rht-D1b and Rht8 in Huang-huai valley winter wheat areas and their influences on agronomic characteristics [J].JournalofTriticeaeCrops,2016,36(8):971.
[34] 慕美财,刘 勇,郭小丽,等.山东小麦品种中矮秆基因 Rht-B1b、 Rht-D1b分布的分子鉴定[J].分子植物育种,2005,3(4):473.
MU M C,LIU Y,GUO X L,etal.Distribution of Rht-B1b and Rht-D1b in wheat cultivars in Shandong detected by molecular markers [J].MolecularPlantBreeding,2005,3(4):473.
[35] 梁 丹,杨芳萍,何中虎,等.利用STS标记检测CIMMYT小麦品种(系)中 Lr34/Yr18、 Rht-B1b和 Rht-D1b基因的分布[J].中国农业科学,2009,42(1):17.
LIANG D,YANG F P,HE Z H,etal.Characterization of Lr34/Yr18, Rht-B1b, Rht-D1b genes in CIMMYT wheat cultivars and advanced lines using STS markers [J].ScientiaAgriculturaSinica,2009,42(1):17.
DistributionofDwarfGenesandTheirAssociationwithAgronomicTraitsinBreadWheatfromtheYellowandHuaiWheatRegion
ZHOUXiaobian,ZHAOLei,CHENJianhui,YANGXia,WANGYongyan,ZHANGXiangfen,YANXuefang,DONGZhongdong,CUIDangqun,CHENFeng
(Agronomy College,Henan Agricultural University/Collaborative Innovation Center of Henan Grain Crops/National Key Laboratory of Wheat and Maize Crop Science,Zhengzhou,Henan 450002,China)
Identification of distribution of dwarf genes and their influence on agronomic traits in bread wheat could contribute to reasonable use of dwarf genes and could provid evaluable information for wheat breeding program. Different molecular markers at sixRhtloci ( Rht1, Rht2, Rht4, Rht8, Rht9 and Rht12) were used to detect distribution of dwarf genes in 246 common wheat from the Yellow and Huai wheat region.A total of ten agronomic traits,including plant height,spike length,peduncle length,spikelet number per spike,flag leaf length,flag leaf width,kernel number per spike,kernel length,kernel width and thousand-kernel weight were investigated for three years and their relationship with theRhtloci were analyzed. The results showed that six dwarf genes showed an extensive distribution in the Yellow and Huai wheat region,especially for the Rht1 and Rht2 genes. Association analysis of the six dwarf genes with the ten agronomic traits in bread wheat showed that at Rht1 locus cultivars with Rht1-B1a and Rht1-B1b alleles did not show significant difference for plant height. At Rht2 locus,cultivars with Rht2-D1b had significantly lower plant height,peduncle length but higher thousand-kernel weight in the three years,suggesting that Rht2-D1b is the superior allele. Excluding the effect of Rht1 and Rht2 genes,due to their strong influence on agronomic traits,the four loci of Rht4, Rht8, Rht9 and Rht12 had also important effects on different agronomic traits in wheat cultivars from the Yellow and Huai valley. Among the four loci, Rht4 played an important role in plant height and thousand-kernel weight and thus Rht4-B1b is the relatively superior allele; Rht8 had important effects on spike length,peduncle length and thousand-kernel weight and thus Rht8-D1b is the relatively superior allele; Rht9 had important effects on plant height and thousand-kernel weight and thus Rht9-A1a is the relatively superior allele; Rht12 had important effects on spike length and thousand-kernel weight and thus Rht12-A1a is the superior allele. Further analysis showed that Rht2 showed the most important effect on plant height among the six loci,which was followed by Rht4. Four ( Rht1, Rht2, Rht8 and Rht12) of the six loci showed significant effect on thousand-kernel weight,of which Rht2 also showed the most significant effect.Analysis of the distribution of the six superior alleles in cultivars from different periods indicated percentage of the superior alleles generally increased from early historical cultivars,recent historical cultivars to modern cultivars in the Yellow and Huai valley,and especially 82.9% of modern cultivars had Rht2-D1b.
Bread wheat; Dwarf genes; Molecular marker; Agronomic traits; Superior alleles
时间:2017-08-08
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170808.0911.002.html
2017-01-13
2017-06-12
国家重点研发计划项目(2016YFD101802);河南省科技创新杰出青年基金项目(174100510001);河南省重大科技专项(161100110500);河南省高校科技创新团队项目(14IRTSTHN010)
E-mail:954336282@qq.com
陈 锋(E-mail:chf0088@163.com)
S512.1;S330
: A
:1009-1041(2017)08-0997-11
