鼎湖山不同森林类型土壤纤毛虫群落比较研究①
2017-09-03徐润林
王 超,徐润林
鼎湖山不同森林类型土壤纤毛虫群落比较研究①
王 超,徐润林*
(中山大学生命科学学院,广州 510275)
为了丰富对鼎湖山自然保护区生物多样性的研究,了解不同植被类型下土壤原生动物群落的特征,本文对采自鼎湖山自然保护区7种不同植被下的土壤样品进行了土壤纤毛虫定性和定量分析。结果显示:在全部土壤样品中,共检出44属70种土壤纤毛虫。在全部纤毛虫种类中,仅有4种出现在全部土壤样中,它们分别是、、和。依据物种出现频率高低,鼎湖山自然保护区土壤纤毛虫中,广布种和特有种较少,而常见种较多。不同植被类型下土壤纤毛虫群落丰度介于1 730 ~ 29 200 ind./g,表现为针叶林-马尾松林<山顶灌草丛<针阔叶混交林<河岸常绿阔叶林<沟谷常绿阔叶林<温带季风常绿阔叶林<山地常绿阔叶林的特点。纤毛虫群落相似性分析显示:各样点土壤纤毛虫群落间处于中等不相似(0.25 ~ 0.5)到中等相似(0.5 ~ 0.75)的水平。从本研究结果看,土壤纤毛虫的分布更符合Foissner的“生物地理模型”。相关性分析显示:土壤含水量与土壤纤毛虫丰度之间有极显著的相关性(<0.01),而土壤NO– 3-N含量与纤毛虫丰度也有显著的相关性(<0.05)。而多元相关分析表明鼎湖山土壤纤毛虫丰度变化与多个土壤理化因子组合共同作用间存在相关性,其中土壤含水量均作为一个重要组合元素出现。CCA分析显示不同的土壤理化因子对一具体纤毛虫种类的作用存有差异。
土壤纤毛虫;群落结构;动物地理模式;鼎湖山自然保护区
森林是以乔木为主体并包括下面的灌木、草被、草木枯落物、林中的土壤、水面、动物、各种微生物等等所构成的生态系统,是维持陆地良好生态环境的关键因素之一。它具有丰富的物种,复杂的结构,多种多样的功能,高生产力、生态系统稳定,现存量高[1]。森林生态系统是地球陆地生态系统的主体,也是陆地上最为复杂的生态系统[2]。森林对土壤的作用是极其复杂的,主要表现在森林所造成的郁闭环境、乔灌木根系对土壤的影响、森林凋落物的归还和枯枝落叶层的形成以及它们的分解对土壤的影响等。土壤原生动物就是生活在土层中或土壤表面凋落物中的原生动物,是起源于淡水水体由淡水环境迁徙到土壤中并不断演化形成的能够适应特定土壤环境的特定原生动物群落[3]。
鼎湖山国家级自然保护区是我国第一个自然保护区,位于广东省肇庆市鼎湖区,23°10′N,112°31′E。该地区属南亚热带季风湿润型气候,夏长冬短,热量丰富,雨量充沛,阳光充足[4],年平均气温21.0 ℃,极端最高气温38.1 ℃,极端最低气温–0.2 ℃,年均降水量为1 927 mm,受季风气候影响,4—9月为雨季,降雨量占全年的80.38%,且多见暴雨[5]。该保护区总面积约1 133 hm2,主要保护对象为南亚热带地带性森林植被类型——季风常绿阔叶林及其丰富的生物多样性。保护区内生物多样性丰富,生长着约占华南地区植物种类1/4的高等植物,其中,桫椤、紫荆木、土沉香等国家保护植物达22种,鼎湖冬青、鼎湖钓樟等华南特有种和模式产地种多达30种,药用植物更是多达1 000种以上[6],动物亦1 000余种,是华南地区生物多样性最富集的地区之一,被生物学家称为“物种宝库”和“基因储存库”[7]。鼎湖山国家级自然保护区的森林生态系统具有显著的区域代表性,有重要的科研价值[8]。长期以来吸引着众多研究人员的目光,研究的内容涵盖森林群落的组成、结构、功能、演替动力学及其演替模型的构建、群落与土壤的理化等指标的关系、土壤营养状况等众多方面的研究,也有关于当地土壤动物在内的生物调查[9-16],但是目前尚未见到关于鼎湖山森林生态系统中土壤原生动物方面的报道。
本文通过对鼎湖山自然保护区内不同土壤样品中纤毛虫的定性和定量分析探讨该保护区内纤毛虫的区系组成、生物多样性以及与土壤环境因子间的关系,为进一步了解保护区内生物多样性和生态环境的现状提供科学依据。
1 材料与方法
1.1 采样点设置与采样
本研究的目的除对鼎湖山土壤纤毛虫群落进行调查外,还希望就土壤原生动物的分布模式进行探讨,故设置采样点时,使各采样点间的距离尽可能短以控制本研究所涉及的面积尽可能小,尽可能消除因采样点的距离而导致的群落结构差异。参照前人对鼎湖山自然保护区植被的调查结果[9, 17],分别在7种不同的森林群落类型中设定了采样点。它们代表的植被类型分别是:沟谷常绿阔叶林(A点)、河岸常绿阔叶林(B点)、温带季风常绿阔叶林(C点)、针阔叶混交林(D点)、针叶林-马尾松林(E点)、山地常绿阔叶林(F点)和山顶灌草丛(G点)。其中E点是人工植被,D点是由之前种植的马尾松林不断演替形成的半人工植被,其他各样地是自然植被。
1.2 土壤样品的采集和前期处理
在每一森林群落类型样点(1 m × 1 m样方)的四角和中央等量采集表层(0 ~ 5 cm)土壤,剔除较大的凋落物,混匀后做好标签,带回实验室。将每个土壤样品分成两份,一份用于测试土壤样品理化指标,一份风干处理。
1.3 土壤样品的理化指标测试
本研究中测定的土壤理化指标包括土壤含水量、总有机质(TOC)、全氮(TN)、全磷(TN)、全钾(TK)、硫酸根(SO2– 4)、NH4+-N(AN)、NO– 3-N(NN)含量及土壤pH和电导率(TDS)。各指标的测定方法均参照国家相关标准进行。
1.4 土壤纤毛虫的定性分析
本研究中对纤毛虫定性研究采用“非淹没培养法”结合蛋白银染色法[18-19]。纤毛虫种类鉴定依据相关文献的描述[20-25]。
1.5 土壤纤毛虫的定量分析
取已风干好的样品,采用“3级10倍”法[26-27]。根据预实验,本研究中采用的是103、104、105三级稀释倍数。
1.6 数据处理
群落多样性指数选择Shannon-Wiener指数(′)[28]。
本研究涉及的数据处理、群落相似性分析(ANOSIM)、等级聚类分析(CLUSTER)以及典型相关分析(Canonical Correlation Analysis,CCA)分别利用SPSS11.5统计软件、Microsoft Excel 2000软件和PRIMER 5.0软件进行。
2 结果分析
2.1 鼎湖山自然保护区不同类型植被下土壤的理化性质
本研究得到的7个采样点的土壤理化性质见表1。比较各土壤样品可以发现,尽管全部样品均采自一个面积范围不大的区域,但土壤样品间的理化指标还是存在一定程度的差别。
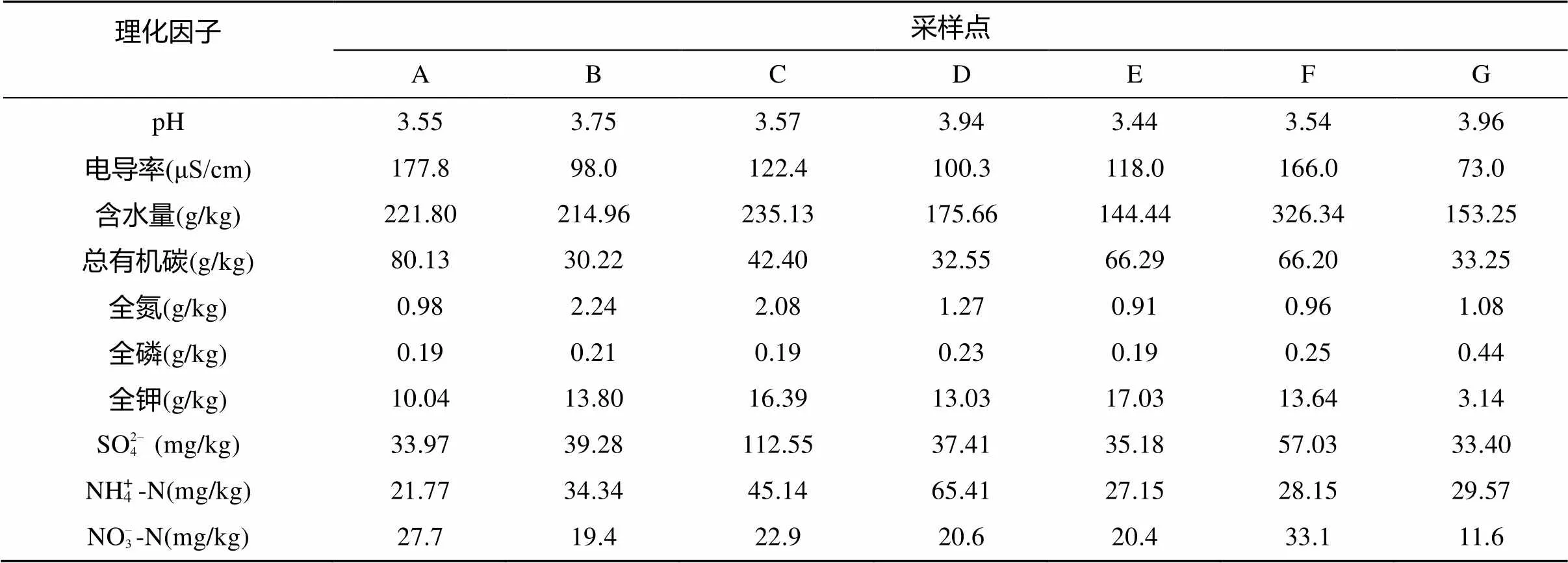
表1 鼎湖山自然保护区不同类型植被下土壤的理化性质
注:A:沟谷常绿阔叶林;B:河岸常绿阔叶林;C:温带季风常绿阔叶林;D:针阔叶混交林;E:针叶林-马尾松林;F:山地常绿阔叶林;G:山顶灌草丛。下同。
2.2 鼎湖山自然保护区不同类型植被下土壤纤毛虫的群落特征
2.2.1 不同类型植被下土壤纤毛虫的群落组成 从全部土壤样品中共发现土壤纤毛虫70种,隶属44个属(表2)。其中,有16种在至少一半以上的样点中出现,其中、、和等4种在7个样点中均有发现。根据物种在各样点出现的频率划分,这些种类包括广布种10种(分布频率0.83 ~1.0),占物种总数的14.3%;常见种44种(分布频率0.50 ~ 0.67),占物种总数的62.9%;特有种16种(分布频率0.17 ~ 0.33),占物种总数的22.9%。由此可见,在鼎湖山自然保护区土壤纤毛虫中,广布种和特有种较少,而常见种较多。
在全部44属中,属中出现的物种最多(8种),其次是属和属(各4种),属出现了3种。这4属出现的种类占了纤毛虫总种数的27.1%(共19种)。
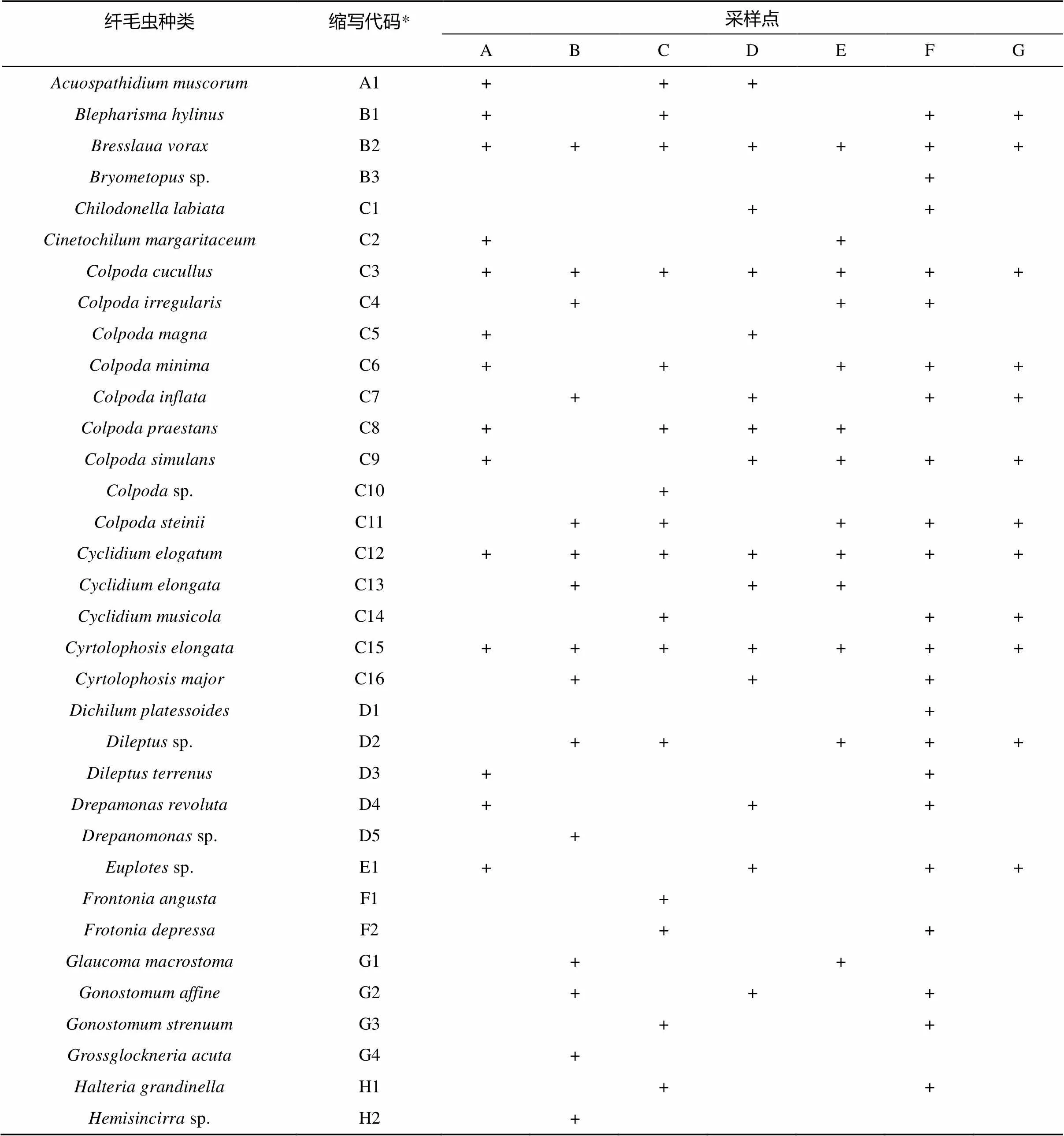
表2 鼎湖山自然保护区不同类型植被下土壤纤毛虫的种类
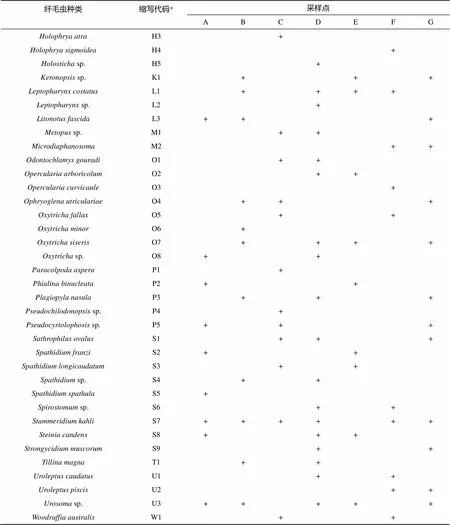
续表
注:“+”表示该种存在;“*”显示在图2中的标识代码。
2.2.2 不同类型植被下土壤纤毛虫群落基本结构 表3列出了鼎湖山自然保护区土壤纤毛虫群落的基本信息。在群落的种类组成上,鼎湖山自然保护区各土壤样品中纤毛虫的种类数介于22 ~ 33(平均27种)。从群落的丰度讲,鼎湖山自然保护区山地常绿阔叶林(样点F)下土壤纤毛虫的丰度最大(29 200 ind./g),针叶林–马尾松林(样点E)和山顶灌草丛(样点G)下土壤纤毛虫的丰度均较低,分别只有1 730 ind./g和2 160 ind./g。在反映生物群落结构复杂性和稳定性的生物多样性指数上,针阔叶混交林(样点D)下土壤纤毛虫群落的Shannon-Wiener指数(′)最高(3.798),而针叶林–马尾松林(样点E)下土壤纤毛虫群落的指数最低(2.349)。
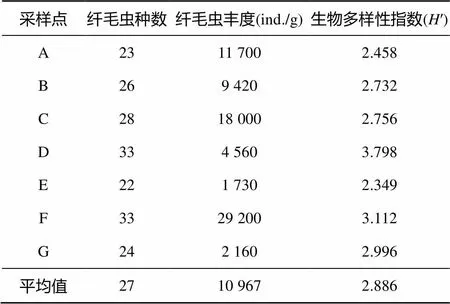
表3 鼎湖山自然保护区不同类型植被下土壤纤毛虫群落的基本参数
2.2.3 不同类型植被下土壤纤毛虫群落间的相似性 根据鼎湖山自然保护区土壤纤毛虫在7个样点的分布特征,计算得各个样点间土壤纤毛虫群落的相似性(表4)。各样点土壤纤毛虫的相似性系数位于0.286(样点A与样点B之间)到0.560(样点B与样点G之间)。按照群落相似性评价原理和标准[28],可以判定鼎湖山自然保护区内土壤纤毛虫群落间的相似性处于中等不相似(0.25 ~ 0.50)到中等相似(0.50 ~ 0.75)的水平。
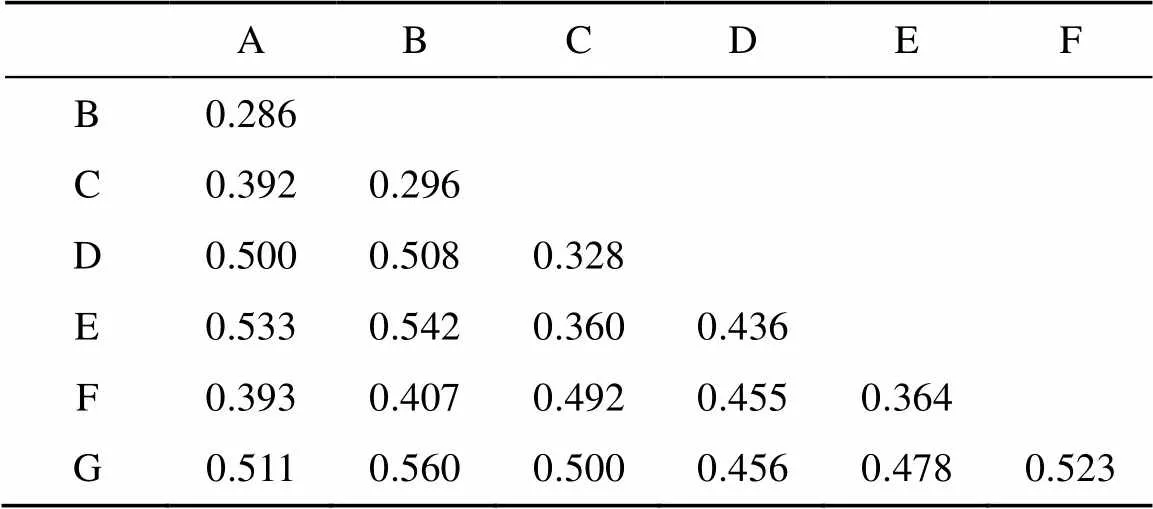
表4 鼎湖山自然保护区不同类型植被下土壤纤毛虫群落间的相似性
对各个样点间的土壤纤毛虫的群落相似性进行聚类分析显示:在鼎湖山自然保护区,河岸常绿阔叶林(B点)与山顶灌草丛(G点)下的土壤纤毛虫群落相似性最高;沟谷常绿阔叶林(A点)与针叶林-马尾松林(E点)下的土壤纤毛虫群落相似性也很高较,而温带季风常绿阔叶林(C点)和山地常绿阔叶林(F点)与其他5种林地类型下的土壤纤毛虫群落相似性相对较低(图1)。
2.3 鼎湖山自然保护区土壤理化性质与纤毛虫群落结构的关系
对鼎湖山自然保护区各种林地土壤理化因子与相对应的土壤纤毛虫物种数间的Pearson相关性分析显示:土壤含水量与土壤纤毛虫种类数间存在极显著的相关性(=0.984,<0.001);土壤NO– 3-N含量与土壤纤毛虫种类数间存在着显著的相关性(= 0.825,<0.005)。而其他理化指标与土壤纤毛虫种类数间并无显著相关性(表5)。在纤毛虫群落丰度方面,土壤含水量与纤毛虫群落丰度间有极显著的相关性(<0.01),而土壤NO– 3-N与纤毛虫群落丰度有显著的相关性(<0.05)。
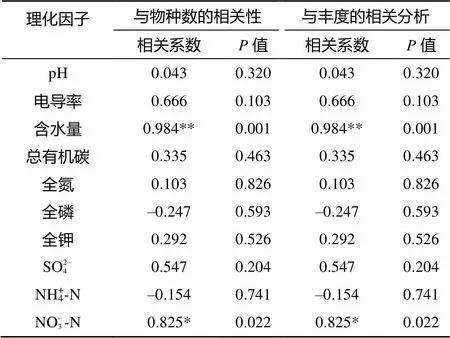
表5 鼎湖山自然保护区土壤纤毛虫群落种类数和丰度与土壤理化因子的Pearson相关分析
注:** 表示在<0.01 水平显著相关(双尾检验);* 表示在<0.05水平显著相关(双尾检验);–表示负相关。
为考虑多个土壤理化因子对土壤纤毛虫群落的综合作用,本研究进行的多元相关分析表明,鼎湖山土壤纤毛虫丰度变化与多个土壤理化因子共同作用间存在相关性。土壤纤毛虫丰度与土壤pH + 含水量组合间的相关性最大(= 0.916);土壤含水量 + 全氮含量组合以及土壤pH + 含水量 + 全氮含量组合与土壤纤毛虫丰度间也存有较高的相关性(表6)。
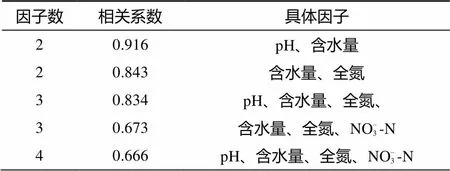
表6 鼎湖山自然保护区土壤理化因子组合与纤毛虫丰度的相关性
通过对各种土壤理化因子与各纤毛虫种类丰度间典型相关分析(CCA),可以发现不同的土壤理化因子对一个具体的纤毛虫种类作用是有差异的(图2)。
3 讨论
到目前为止,我国有关土壤纤毛虫的研究主要集中在分类和区系的调查,涉及的地域南北均有[29-36]。对比这些文献,如果单从本研究中共发现土壤纤毛虫70种,且各样点土壤纤毛虫群落由22 ~ 33种组成这一结果看,基本与国内外其他地区土壤纤毛虫研究结果相近。但如对各群落组成做进一步的比较,可以发现,土壤纤毛虫群落组成不仅在大尺度地域间存在明显差异,即使在小尺度距离间也存有明显差异,即土壤纤毛虫群落间的相似性处于中等不相似(0.25 ~ 0.50) 至中等相似(0.50 ~ 0.75)的水平。群落的相似性与该类动物的分布模式存在很大关系。对于原生动物的动物地理分布模式,国际上存在着两种截然对立的观点。以Finlay为代表的观点认为:人们已基本了解了大多数自由生活的原生动物种类;在多数情况下自由生活原生动物是以包囊形式存在;由于包囊扩散非常容易,因此它们是全球性分布的;自由生活原生动物的区域物种多样性与全球物种多样性呈很高的比值,故它们在各地的分布并没有明显的差异。这一观点被称为自由生活原生动物的“普遍分布模型”(ubiquity model)理论[37-39]。而以Foissner为代表的“生物地理模型”(biogeography model)理论则认为:大多数自由生活原生动物的分布受区系地理的限制;由于技术手段上的局限性,原生动物的鉴定存在很多困难;鉴定时很多少见种被数量较大的常见种所掩盖,因而人们得到的结果被严重误导[40-42]。根据本研究结果并结合国内其他相关报道,本研究认为土壤纤毛虫的分布更符合“生物地理模型”。
作为土壤生态系统中的重要组成部分,土壤原生动物与土壤环境间的关系也是大家非常关注的问题。大量的文献表明:一定的时空范围内,影响土壤动物群落的因素很多。小型土壤动物数量对环境变化的反应比大型土壤动物敏感,对环境影响的依赖性强于大型土壤动物。植被为土壤动物提供生存环境的同时,也对土壤动物群落的组成产生影响,植被类型与土壤动物的分布变化密不可分,对土壤动物群落的组成具有一定的影响[43]。鼎湖山自然保护区不同植被类型下土壤纤毛虫群落丰度介于1 730 ~ 29 200 ind./g,且具有针叶林-马尾松林<山顶灌草丛<针阔叶混交林<河岸常绿阔叶林<沟谷常绿阔叶林<温带季风常绿阔叶林<山地常绿阔叶林的特点,也验证了此观点。
有关环境因子对土壤原生动物群落的影响虽有报道,但尚不系统。尽管如此,有关土壤理化因子对原生动物的作用还是存在一些共识。如土壤水分和土壤的机械组成是影响土壤纤毛虫丰度的最主要因素[27]。土壤中的水是土壤内部物理、化学和生物过程不可缺少的介质。土壤原生动物是起源于淡水,且经过多次向土壤集群后形成的具有特定适应性、独立的类落[3]。纤毛虫的各种生命过程均离不开水,因此土壤含水量也就成为土壤原生动物的重要限制因子[32, 44]。对鼎湖山自然保护区内土壤纤毛虫群落与土壤理化因子的相关性分析结果也说明了土壤含水量在纤毛虫的分布上起着极为重要的作用(表5和表6)。
但由于土壤系统的复杂性,任何栖息在此的生物均会受到其周围多种理化因子的影响。多元相关分析与典型相关分析(CCA)均显示鼎湖山自然保护土壤纤毛虫群落均受到多种环境因子的影响,每种因子对于每个具体纤毛虫种可产生不同的效应,其综合结果就导致了各种植被类型下土壤纤毛虫群落结构上的差异。
4 结论
在鼎湖山自然保护区采集的7种植被类型土壤样品中,共检出44属70种土壤纤毛虫,但仅有少数种类出现在了全部土壤样中;以广布种和特有种的较少,而常见种较多。不同植被类型下土壤纤毛虫群落的种类数介于22 ~ 33(平均27种),其丰度介于1 730 ~ 29 200 ind./g。各样点土壤纤毛虫群落间处于中等不相似(0.25 ~ 0.50)到中等相似(0.50 ~ 0.75)的水平。
统计分析显示:土壤含水量与土壤纤毛虫种类数间存在极显著的相关性(=0.984,<0.001);土壤NO– 3-N含量与土壤纤毛虫种类数间存在着显著的相关性(= 0.825,<0.005)。同时,土壤纤毛虫丰度变化与多个土壤理化因子共同作用间存在相关性。
本研究结果表明鼎湖山土壤纤毛虫的分布更符合Foissner提出的“生物地理模型”。
[1] 蔡晓明. 生态系统生态学[M]. 北京:科学出版社, 2002
[2] 马定国, 舒晓波, 刘影, 等. 江西省森林生态系统服务功能价值评估[J].江西科学,2003, 21(3): 211–216
[3] Foissner W. Soil protozoa: Fundamental problems, ecological significance, adaptations in ciliates and testaceans, bioindicators, and guide to the literature[J]. Progress of Protistology, 1987, 2: 69–212
[4] 肖光明. 鼎湖山旅游地生命周期的分析与调控[J]. 热带地理, 2003, 23(3): 180–184
[5] 黄展帆, 范征广. 鼎湖山的气候[J]. 热带亚热带森林生态系统研究, 1982(1): 11–23
[6] 张宏伟, 马骥. 鼎湖山药用植物资源调查分析[J]. 广西植物, 2005, 25(6): 539–543
[7] 莫萍. 北回归线上的绿宝石——鼎湖山[J]. 南方国土资源, 2009, 2(1): 37–39
[8] 彭少麟. 南亚热带森林群落动态学[M]. 北京: 科学出版社, 1996
[9] 曹洪麟, 黄忠良, 张林艳, 等. 鼎湖山自然保护区植被类型[J]. 热带亚热带森林生态系统研究, 2002(9): 1–9
[10] 黄文娟, 刘菊秀, 唐旭利, 等. 鼎湖山5种森林土壤的无机氮和有效磷含量[J]. 应用与环境生物学报, 2009, 15(4): 441–447
[11] 何宜庚. 广东省鼎湖山自然保护区的土壤[J]. 华南师范大学学报 (自然科学版), 1983, 1(1): 87–96
[12] 李耀泉. 鼎湖山常见昆虫调查初报[J]. 热带亚热带森林生态系统研究, 1984, 2: 69–81
[13] 廖崇惠. 鼎湖山森林土壤动物研究Ⅰ:区系组成及其特征[J]. 热带亚热带森林生态系统研究, 1989(5): 83–96
[14] 林英华, 张夫道, 张俊清, 等. 鼎湖山不同自然植被土壤动物群落结构时空变化[J]. 生态学报, 2005, 25(10): 2616–2622
[15] 陈茂乾, 廖崇惠. 鼎湖山森林土壤动物研究,Ⅱ:不同生境的群落组成[J]. 热带亚热带森林生态系统研究, 1990(7): 90–99
[16] 徐国良, 黄忠良, 欧阳学军, 等. 广东鼎湖山自然保护区动物物种多样性[J]. 四川动物, 2002, 21(2): 72–75
[17] 王铸豪, 何道泉, 宋绍敦. 鼎湖山自然保护区之植被[J]. 热带亚热带森林生态系统研究, 1982(1): 77–141
[18] Foissner W. Estimating the species richness of soil protozoa using the “flooded petridish method”[A]// Lee J J, Soldo A T. Protocols in Protozoology. Lawrence: Allen Press, 1992: 1–2
[19] Foissner W. Basic light and scanning electron microscopic methods for taxonomic studies of ciliated protozoa[J]. European Journal of Protistology,1991, 27(2): 313–330
[20] Kahl A. Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria)[M]. Jena: Verlag von Gustav Fischer, 1935
[21] Foissner W. Colpodea (Ciliophora)[M]. Stuttgart, Germany, Jena, New York: Gustav Fischer Verlag, 1993
[22] Berger H. Monograph of the Oxytrichidae (Ciliophora, Hypotrichia)[M]. Dordrecht: Kluwer Academic, 1999
[23] Foissner W, Agatha S, Berger H, et al. Soil ciliates (,) from Nambibia (Southwest Africa), with enphasis on two contrasting environments, the Etosha Region and the Nambia Desert[M]. Austria: Oberosterreichisches Landesmuseum Lina, 2002
[24] Lynn D H. The ciliated protozoa, characterization, classification, and guide to the literature[M]. 3rd ed. London: Springer, 2008
[25] 沈韫芬. 原生动物学[M]. 北京: 科学出版社, 1999
[26] Darbyshire J F, Wheatley R E, Greaves M P, Inkson R H E. A rapid micromethod for estimating bacterial and protozoan populations in soil[J]. Review Ecology and Biology of Soil, 1974, 11(4): 465–475
[27] Ekelund F, Rønn R. Notes on protozoa in agricultural soil with emphasis on heterotrophic flagellates and naked amoebae and their ecology[J]. FEMS Microbiological Review, 1994, 15: 321–353
[28] 赵志模, 郭依泉. 群落生态学原理与方法[M]. 北京: 科学技术文献出版社, 1990
[29] 崔振东. 长白山北坡针阔混交林土壤原生动物的生态分布[J]. 生态学杂志, 1986, 5(2): 1–5
[30] 宁应之, 沈韫芬. 珞珈山森林土壤原生动物生态学研究及土壤原生动物定量方法探讨[J]. 动物学研究, 1996, 17(3): 225–232
[31] 宁应之, 沈韫芬. 中国典型地带土壤原生动物调查Ⅰ: 区系特征与种类分布[J]. 动物学报, 1998, 44(1): 5–10
[32] 宁应之, 沈韫芬. 中国典型地带土壤原生动物调查Ⅱ: 生态学研究[J]. 动物学报, 1998, 44(3): 271–276
[33] 陈素芳, 徐润林, 王勇军, 等. 化学防除薇甘菊对内伶仃岛土壤原生动物群落的影响[J]. 应用与环境生物学报, 2003, 9(3): 422–428
[34] 王慧, 桂娟, 刘满强, 等. 稻草和三叶草分解对微型土壤动物群落的影响[J]. 土壤学报, 2015, 52(5): 1124–1134
[35] 宁应之, 王娟, 刘娜, 等. 甘肃天水麦积山风景名胜区土壤纤毛虫的物种多样性[J]. 动物学研究, 2007, 28(3): 367–373
[36] 徐润林, 孙逸湘. 大鹏半岛土壤纤毛虫的群落特点[J]. 应用生态学报, 2000, 11(3): 428–430
[37] Finlay B J, Corliss J O, Esteban G. Biodiversity at the microbial level: the number of free-living ciliates in the biosphere[J]. Quarterly Review of Biology, 1996, 71(2): 221–237
[38] Finlay B J, Esteban G F, Clarke J K. Biodiversity of terrestrial protozoa appears homogeneous across local and global spatial scales[J]. Protist, 2001, 152(3): 355–366
[39] Finlay B J. Global dispersal of free-living microbial eukaryote species[J]. Science, 2002, 296(5570): 1061–1063
[40] Foissner W. Protist diversity: estimates of the near-imponderable[J]. Protist, 1999, 150(3): 363–368
[41] Foissner W. Biogeography and dispersal of micro- organisms: a review emphasizing protists[J]. Acta Protozoology, 2006, 45(2): 111–136
[42] Foissner W. Dispersal and biogeography of protists: recent advances[J]. Japan Journal Protozoology, 2007, 40(1): 1–16
[43] 武海涛, 吕宪国, 杨青, 等. 土壤动物主要生态特征与生态功能研究进展[J]. 土壤学报, 2006, 43(2): 314–323
[44] Stout J. Reaction of ciliates to environmental factors[J]. Ecology, 1956, 37(1): 178–191
Comparative Study on Soil Ciliates Community Structures Under Different Vegetation Types in Dinghu Mountain
WANG Chao, XU Runlin*
(School of Life Sciences, Sun Yat-Sen University, Guangzhou 510275, China)
In order to expand the research of biodiversity in Dinghu Mountain Nature Reserve and understand further and more about the characteristics of soil protozoa community under different vegetation types, soil ciliates from 7 soil samples under different vegetations were cultured with the qualitative and quantitative methods. From all soil samples, 70 ciliates species, belonged to 44 genus, were found. In all ciliates, only,,andappeared inall soil samples. It could be found that widely distributed species and endemic species of ciliates were less, otherwise, common species of ciliates were more according to the species observed in all samples. The abundances of soil ciliates communities under different vegetation types ranged from 1 730 to 29 200 ind./g, and followed the order as conifer and pinus forest < grass near mountain shrub < coniferous and broad-leaved mixed forest < evergreen broad-leaved forest near riparian < evergreen broad-leaved forest in ravine < evergreen broad-leaved forest in temperate monsoon < evergreen broad-leaved forest in montane. The similarity of soil ciliates communities in different vegetation types displayed from moderate dissimilar (0.25 to 0.5) to moderate similar (0.5 to 0.75) between each other. The results implies that the distribution of soil ciliates was more consistent with the Foissner’s “biogeography model”. Correlation analysis revealed an extremely significant correlation (< 0.01) existed between soil moisture and ciliates abundance, and a significant correlation between soil NO– 3-N and ciliates abundance (< 0.05). The multiple correlation analysis showed correlations existed between soil ciliates abundance and the combinations of soil physico-chemical factors, and soil moisture appeared in each combination as an important element. CCA analysis showed that different soil physico-chemical factors had different effects on a certain ciliate species.
Soil ciliates; Community structure; Zoogeographic pattern; Dinghu Mountain Nature Reserve
10.13758/j.cnki.tr.2017.04.013
Q89;S154.36
A
国家自然科学基金项目(31030015)资助。
(xurunlin_1960@yahoo.com)
王超(1989—),男,山东人,硕士,主要研究方向为动物生态学。E-mail: wangchao@qq.com
