辣椒全基因组TCP转录因子家族生物信息学分析
2017-09-03冯羽娥
冯羽娥
(西北农林科技大学园艺学院,陕西咸阳712100)
辣椒全基因组TCP转录因子家族生物信息学分析
冯羽娥
(西北农林科技大学园艺学院,陕西咸阳712100)
利用生物信息学方法对辣椒TCP转录因子家族成员、基因分类、基因结构、染色体定位、系统进化关系和结构域序列保守性进行了预测和分析。结果表明,辣椒TCP基因家族包含30个成员,CaTCP蛋白含有95~550个氨基酸,等电点为6.30~10.91。进化树分析将其分为11个不同的亚族。辣椒TCP家族进化树以及基因结构的进一步分析,显示辣椒TCP家族可分为两大类,即ClassI类和ClassII类,而ClassII类可分为两个不同的亚类,即CIN和CYC/TB1亚类。位于同一亚家族的大多数TCP基因具有相似的内含子和外显子结构。此外,染色体定位分析表明,辣椒TCP基因不均衡的分布于辣椒的10条染色体上,其中3号染色体上最多,有7个。
辣椒;TCP;转录因子;全基因组分析
1 材料与方法
在Pfam数据库(http∶//pfam.xfam.org/)下载TCP结构域序列(PF03634),用于在辣椒全基因组数据库(PGD,http∶//peppergenome.snu.ac.kr/,CM334 and Zunla-1 genomes)中检索辣椒TCP基因。同时,拟南芥AtTCP蛋白序列也作为诱饵序列在PGD和NCBI数据库中进行检索。所有检索出的基因(E-value≤0.001)都利用PFAM及NCBI-CDD工具进行蛋白结构预测,删除不含TCP结构域的基因。利用在线程序Ex-PASy ProtParam(Artimo et al.2012;http∶//web. expasy.org/protparam)对辣椒CaTCP基因编码蛋白的等电点和分子量进行分析。
利用ClustalW程序对拟南芥和辣椒TCP蛋白序列进行多序列比对,将多序列比对结果利用MEGA6.0软件进行构建,采用邻接法(Neighbor-Joining,NJ),重复1 000次。利用GSDS程序(GuoAY et al.2007;Gene Structure Display Server∶http∶//gsds.cbi.pku. edu.cn/)对辣椒CaTCP基因结构进行分析。
利用DNAMAN软件对辣椒CaTCP蛋白序列进行多序列比对分析。利用MapDraw工具对辣椒CaTCP家族成员进行染色体定位作图。
2 结果与分析
2.1 辣椒CaTCP转录因子家族的鉴定与分类
通过检索后再进行结构域查找,共从辣椒基因组中鉴定得到30个CaTCP转录因子家族成员。根据其所在染色体的位置,对30个辣椒CaTCP基因进行了系统编号。CaTCP对应的基因编号、基因ID、所在染色体位置、编码序列长度、氨基酸序列长度、分子量以及等电点等特征见表1;对辣椒CaTCP基因编码蛋白进行了理化性质的分析,详细情况见表1。辣椒CaTCP基因编码蛋白所含氨基酸个数差异较大,最少编码95个氨基酸(CaTCP8和CaTCP10),最多编码550个(CaTCP20),平均为315个。分子量在59008.4Da(CaTCP20)到17 892.7Da(CaTCP23)之间;CaTCP蛋白的等电点基本都大于7,平均值为8.34,只有6个CaTCP蛋白(CaTCP1、4、11、17、29和30)的等电点小于7(分别为6.51、6.30、6.79、6.80、6.67和6.96),这说明大多数CaTCP蛋白含有较多的碱性氨基酸。
2.2 辣椒与拟南芥TCP家族系统进化及保守结构域分析
图1为辣椒上的30个CaTCP蛋白序列与24个拟南芥AtTCP蛋白序列构建的进化树。有的进化枝上的数值相较其他大部分数值比较低,但这在前人的研究中也出现过。由图1可以看出,根据其TCP结构域特点,TCP转录因子家族可以分为11个亚家族(Group A到Group K)。其中Group A、B和C属于class II类亚家族中的CIN类,而Group D和E属于class II类亚家族中的CYC/TB1类;而其余的(Group F、G、H、I、J和K)则属于class I类亚家族成员。这两个物种中的TCP基因基本上在所有进化枝上都有分布。许多拟南芥TCP基因在辣椒中拥有2个或更多的复本,表明CaTCP基因的复制发生于辣椒与拟南芥分离之后。如Group D中包含7个辣椒CaTCP基因,但却只有3个拟南芥AtTCP基因;Group C中包含5个辣椒CaTCP基因,但却只有3个拟南芥AtTCP基因。
许多具有相似功能的拟南芥TCP基因可聚集到一簇中,这可能意味着在同一进化枝上的辣椒TCP基因具有相似的功能。如Group I和Group K中与5个CaTCP基因聚为一簇的所有AtTCPs(AtTCP8、AtTCP14、AtTCP15、AtTCP22、AtTCP23)基因在通过介导细胞周期以及维持顶端分生组织调控叶片发育方面起重要作用。Group D中的所有AtTCPs在调控植株侧枝发育中起重要作用。Group B和Group C中与7个CaTCP基因聚为一簇的所有AtTCP基因(AtTCP2、AtTCP3、AtTCP4、AtTCP10、AtTCP24)受miRNA319负调控,且在调控叶片边缘发育中负调控细胞增殖。
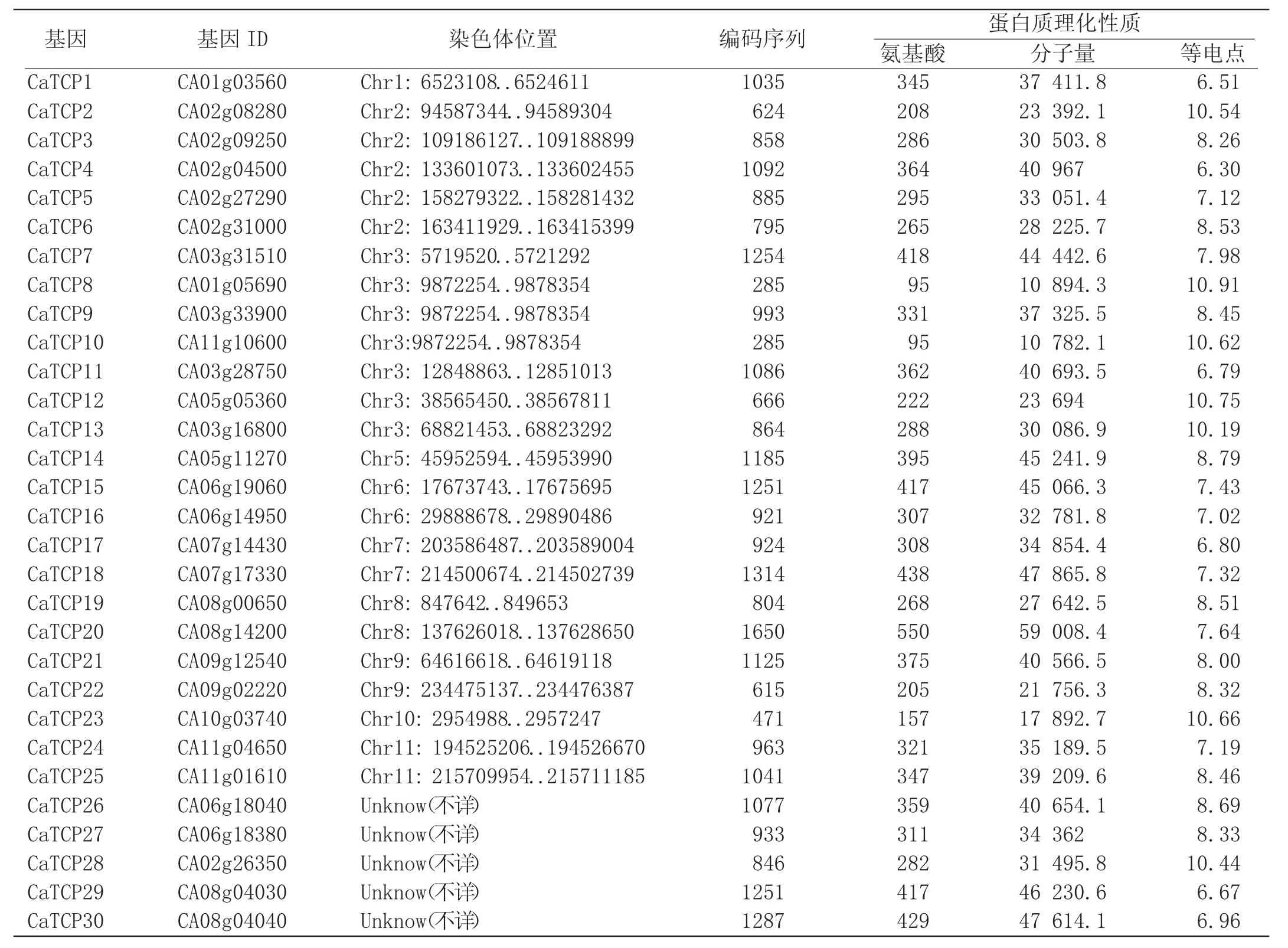
表1 辣椒CaTCP基因家族鉴定
对预测的30个辣椒CaTCP转录因子蛋白序列进行保守结构域序列比对(图2),结果发现,辣椒CaTCP蛋白中均包含有TCP结构域,即TCP转录因子的特征序列—bHLH结构域。
2.3 辣椒CaTCP家族系统进化及基因结构分析
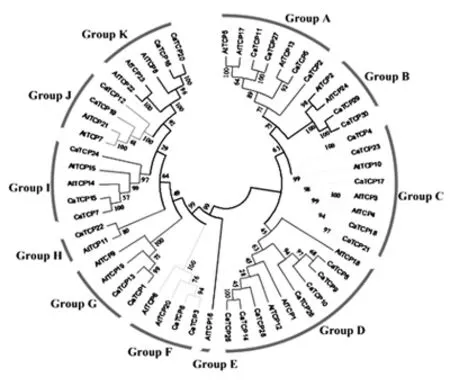
图1 拟南芥和辣椒TCP蛋白家族的进化分析

图2 辣椒TCP转录因子家族多重序列比对
为了进一步了解CaTCP基因的进化关系,通过其cDNA序列以及对应的DNA序列进行比对,对每个CaTCP基因的外显子/内含子结构进行分析研究(图3)。如图3 b可以看出,大多数CaTCP的基因结构较为简单,30个CaTCP基因中27个没有内含子,其他3个(CaTCP1、CaTCP16和CaTCP20)都只有1个内含子。此外,为验证CaTCP基因的外显子/内含子结构是否与其进化树亚家族一致,对CaTCP的蛋白序列进行了进化树分析(图3 a)。如图3 a可以看出,大部分位于同一亚家族的CaTCP基因拥有相似的外显子/内含子分布结构(外显子长度和内含子数量)。例如,在A、B、C、F和I亚家族的大部分CaTCP基因只有一个具有相似长度的外显子,但没有内含子;然而,在K亚家族中仅有的2个CaTCP基因都包含一个内含子。
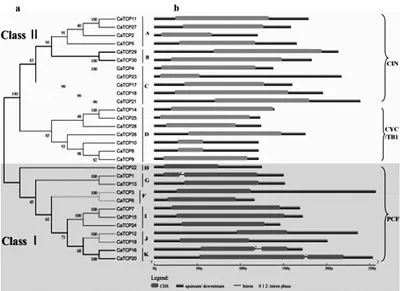
图3 辣椒CaTCP基因的系统进化关系及外显子/内含子结构
2.4 辣椒CaTCP基因的染色体分布
对辣椒CaTCP基因进行染色体定位分析,30个CaTCP基因只定位到25个(CaTCP26、27、28、29和30未定位到染色体)。如图4所示,在辣椒的12条染色体中,25个辣椒CaTCP基因定位到10条染色体上,第4号和第12号染色体上没有CaTCP基因分布。一般而言,每条染色体上CaTCP基因分布不均,数量从0到7不等。其中,第3号染色体上的CaTCP基因数目最多,分布7个CaTCP基因,占总基因数量的23.3%,其次为2号染色体,分布5个CaTCP基因;而第1、5和10号染色体上都只有1个CaTCP基因。
3 讨论与结论
TCP转录因子在植物中分布较广,各物种间的分布数量存在一定差异。拟南芥TCP转录因子家族有24个成员,水稻有22个成员(Yao et al.2007;Cubas et al.2002),本研究鉴定了30个辣椒TCP家族成员,通CaTCP蛋白含有95~550个氨基酸,等电点为6.30~10.91。进化树分析将其分为11个不同的亚族。辣椒TCP家族进化树以及基因结构的进一步分析显示辣椒TCP家族可分为两大类,即Class I类和Class II类,而Class II类可分为两个不同的亚类,即CIN和CYC/TB1亚类。位于同一亚家族的大多数TCP基因具有相似的内含子和外显子结构。染色体定位分析表明辣椒TCP基因不均匀的分布于辣椒的10条染色体上,其中3号染色体上最多,有7个。
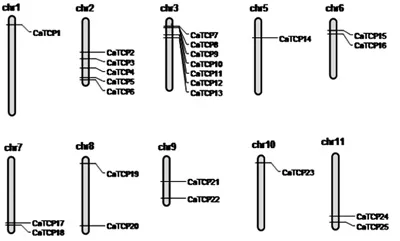
图4 辣椒CaTCP基因的染色体定位
通过系统进化分析发现,辣椒和拟南芥中的TCP家族基因分散在进化树的各个分支,且拟南芥中的许多TCP基因在辣椒中拥有2个或2个以上的复本,这表明辣椒TCP家族的扩展可能是由于基因组重组(如片段重复、串联重复和转座子)导致的。CaTCP8、CaTCP9、CaTCP10、CaTCP14、CaTCP25、CaTCP26和CaTCP28与AtTCP1、AtTCP12和AtTCP18的同源性较高,说明他们有可能在调控植株侧枝发育中起重要作用(Aguilar-Martinez et al.2007);而与拟南芥AtTCP基因(AtTCP2、AtTCP3、AtTCP4、AtTCP10、AtTCP24)聚为一簇的Group B和Group C中的7个CaTCP基因有可能作为miRNA319的靶基因参与辣椒叶片的发育(Palatnik et al.2003)。
借鉴拟南芥、水稻及其他物种中的TCP转录因子在植物生长发育和相关代谢过程中的功能研究,探索CaTCP转录因子在辣椒生长发育尤其是叶片和侧枝、某些激素代谢过程中的功能及其相互作用机制,是今后辣椒TCP基因相关研究的重点。
1005-2690(2017)08-0152-03
S641.3
B
2017-07-13)
