广西野生蕉种质资源调查与鉴定
2017-08-08秦献泉安振宇黄伟雄尧金燕
龙 兴,秦献泉,方 仁,邓 彪,安振宇,黄伟雄,尧金燕
(广西农业科学院园艺研究所,广西 南宁 530007)
广西野生蕉种质资源调查与鉴定
龙 兴,秦献泉,方 仁,邓 彪,安振宇,黄伟雄,尧金燕*
(广西农业科学院园艺研究所,广西 南宁 530007)
【目的】较全面地收集并鉴定广西野生蕉种质资源的亲缘关系和分类地位,为广西野生蕉资源的保护和利用提供理论依据。【方法】采用染色体制片压片法、形态学分类法和荧光AFLP标记分别对广西境内的14份野生蕉种质进行染色体倍性、形态学分类和亲缘关系的鉴定与研究。【结果】形态学分类鉴定表明,供试的14份野生蕉种质资源染色体倍性均为二倍体,2n=22(x=11)。其中2份鉴定为M.acuminatasubsp.malaccensis种,其余12份鉴定为M.itinerans种,种间形态学性状差异较大。基于AFLP标记的鉴定结果表明广西野生蕉资源具有丰富的遗传多样性,与形态学鉴定结果基本一致。【结论】供试的14份野生蕉均属于芭蕉属真蕉组(Eumusa),其中2份归属为小果野蕉亚种,12份归属为阿宽蕉,该种在广西分布较普遍。AFLP鉴定结果显示广西野生蕉种质资源间遗传差异较大,与栽培蕉亲缘关系也较远。
野生蕉;种质资源;分类鉴定;形态学标记;AFLP;广西
【研究意义】野生蕉是芭蕉属野生近缘种植物的统称,我国地处全球主要香蕉野生种质资源分布区域的北缘,云南、海南、福建、广东及广西等省(区)均拥有丰富的野生蕉资源[1-3]。广西地处低纬,气候环境非常适宜野生蕉种质资源繁衍,在许多市县乡镇均发现了野生蕉的分布,资源十分丰富,然而缺乏系统深入的鉴定及遗传关系研究,致使在优异基因资源挖掘与利用上受到很大限制。因此开展广西野生蕉种质的调查和鉴定研究对于资源的保护和利用具有重要意义。【前人研究进展】李锡文[3]、刘爱忠[4]和Hakkinen[5]等学者对分布于我国云南等地区的香蕉野生种质资源均有系统研究和报道。关于广西野生蕉类资源的研究目前仅有少量报道[6-7],有待鉴定与发掘利用。 在香蕉种质鉴定的诸多方法中,形态学标记一直被视为蕉类种质资源分类的主体方法。自从1947年Cheesman[8]依据染色体数目和形态学性状等将芭蕉属植物划分为四个组蕉类形态学标记分类系统经过多位学者[9-10]的补充完善,已具有较好的规范性和合理性。与形态学标记相比DNA分子标记在种质资源鉴定应用中具有更稳定高效的优越性。目前RFLP、RAPD、SSR、ISSR及AFLP等分子标记技术在果树亲缘关系与品种鉴定研究中大量应用,其中AFLP标记技术因具有稳定性高、重复性好、多态性检测效率高的特点,在国内外香蕉品种鉴定和遗传多样性的研究上广泛成功应用[11-15],有助于解决形态学分类方法的不足。【本研究切入点】由于目前许多形态标记遗传表达易受环境和发育期的影响[1],且对于组间杂交种或特异种质的鉴定适应性不强[15],因此结合分子标记技术鉴定能更利于种质资源鉴定和遗传关系研究。【拟解决的关键问题】长期着力于对广西境内野生蕉资源的收集,并利用形态学分类和AFLP标记技术对其进行鉴定,旨在探讨广西野生蕉的遗传背景,为其在香蕉遗传改良上的应用提供理论依据。
1 材料与方法
1.1 野生蕉的调查和收集
从2013年4月至2016年8月,在广西境内14个地县开展野生蕉类种质资源分布情况的调查与收集工作,观察记录其在分布地的各种性状,并采集野生种质的吸芽。
1.2 野生香蕉种质资源的鉴定
1.2.1 染色体倍性鉴定 ①材料准备:广西农业科学院园艺研究所香蕉资源圃内挖取野生种质的第二代吸芽吸芽(种质名称和来源见表1),取高40~70 cm的剑叶芽假植于沙床中,25~30 ℃条件下进行培养生根,待新根长出后(3~5 d)取长约1 cm、粗约0.2 cm的新鲜不定根用于染色体制片和分析。② 染色体制片:参照李懋学[16]压片法,结合去壁低渗法,略作修改。步骤为:预处理、前低渗、固定、酸解、后低渗、染色压片。取材时间为早上7:00-8:00, 预处理用0.02 M 8-羟基喹啉+0.2 %秋水仙素混合液处理4 h。用0.075 M KCl溶液前低渗30 min,酸解用1 mol/L的盐酸60 ℃水浴中解离10 min,再转入0.075M KCl溶液后低渗10 min。用蒸馏水冲洗3次,最后染色,压片。③染色体计数和倍性的确定。用Olympus显微镜在10倍目镜,100/1.35油镜下观察。每个样品观察5个根尖,挑选至少30个完整、染色体清晰、分散好的细胞进行计数,以占优势细胞染色体数目作为该种的染色体数目。蕉类存在随体与小染色体混在一起,二倍体(2x=22)蕉类多1~2条,三倍体(3x=33)多1~3条[17]。所以将染色体数目为23、24的细胞和34、35、36的细胞分别计算在二倍体和三倍体细胞范围内。
1.2.2 形态学鉴定 形态学描述参考INIBAP1995年修订的《Descriptors for Banana(Musaspp.)》[10],形态学分类采用Simmonds(1962)[9]分类法则。
1.2.3 AFLP分子标记鉴定 样品选取与准备。为鉴别广西野生蕉与栽培蕉之间的亲缘关系,在AFLP标记鉴定中加入10个栽培种(包括AAA、ABB和AA3个基因型)和1个长梗蕉(BB基因型),供试种质如表2所示。选取健壮、无病虫危害种质的嫩叶置于保鲜薄膜袋,装入冰壶中3 h内携回实验室,以自来水和无菌水冲洗后立即提取DNA,或吸水纸吸干叶面水份后保存于-80 ℃超低温冰箱中备用。
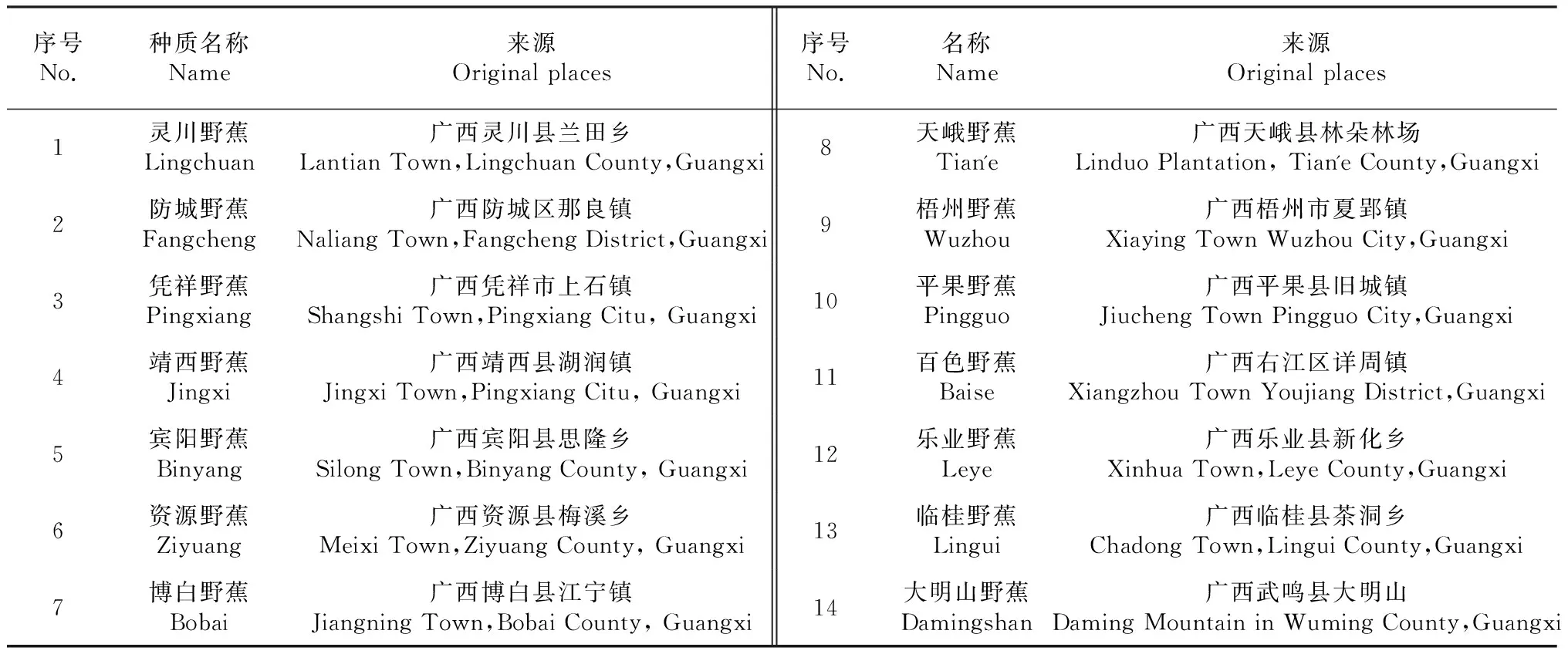
表1 野生蕉类材料及来源

表2 AFLP分析所用香蕉种质材料
注:字母Z开头的种质资源为栽培蕉类型,字母Y开头的表示野生蕉类型,字母后面数字为泳道编号。 *为了验证分子标记的准确性,特设两对相同的种质(泳道1,2和泳道13,14)做标记。 Note: The germplasms of the letter Z forMusacultivation and the letter Y for wildMusa, the figures behind the letter for the lane number.*In order to verify the accuracy of molecular markers, ad hoc two pairs of the same germplasm(lane1,2 and lane 13,14)mark.
DNA提取、AFLP扩增及产物检测。总DNA提取采用改进的CTAB法。AFLP分析试剂盒(EcoRI/MseI型)从北京鼎国生物技术有限公司购买。从试剂盒提供的64对引物中筛选8对多态性好的引物进行扩增。反应程序及详细步骤参照文献[18]。利用GeneScan 3.1软件将8对荧光引物产生的电泳胶图转换为(0,1)矩阵。用NTSYSpc-2.11F软件进行数据分析。对原始矩阵用SimQual程序求DICE相似系数矩阵,并获得相似系数矩阵。用 SHAN程序中的 UPGMA方法进行聚类分析,并通过 Tree plot模块生成聚类图。
2 结果与分析
2.1 广西野生蕉的分布与收集情况
广西境内野生蕉资源丰富且跨域大,北至资源县,南至凭祥县,西至天峨县,东至博白县都发现有野生蕉分布,其中在百色县、博白县、防城港市防城区仍保存着面积达130~450 hm2,分布密度较大的蕉群。广西野生蕉种质多数集中生长在海拔260~700 m土壤湿润的原始林山谷中,少数生长在坡面,甚至海拨较高的山坡上,如大明山野蕉分布在海拨1000 m以上的高处,百色市右江区野生蕉分布则从山谷一直延伸至山顶。由于广西境内野生蕉分布区域气候变化较大,分布在桂南地区海拨较低的部分野生蕉可全年开花,而分布在桂北或高海拨地区的野生蕉花期主要集中在7-9月,但植株能承受冬季零度左右的寒冷气温。可见广西野生蕉分布地形复杂,生态环境差异较大,可能携带着不同的抗性基因,可用作香蕉遗传改良的育种材料。本研究共收集到14种野生蕉种质资源样品。
2.2 广西野生蕉的形态特征
从表3可以看出,广西野生蕉多数具有明显的匍行茎,部分在假茎高度、雄蕾形状、苞片颜色及果指色泽等性状上有明显的差别,如不同野生蕉资源抽蕾时假茎高度最小的为112 cm,最大的达589 cm;雄蕾形状有陀螺状等4个类型;苞片背部颜色以紫红色为主,多数带彩纹或褪色线,以黄,绿或黄绿条纹为主。果指青果色泽有紫红、黄、青色等3种。
2.3 广西野生蕉的鉴定
2.3.1 染色体倍性的鉴定 从表4可知:供试的14份广西野生蕉种质染色体二倍体细胞百分比都在90.00 %以上,平果野蕉最高占98.61 %,因此可以确定供试的14份野生蕉染色体为二倍体,染色体数目2X=22,部分野生蕉细胞染色体图像情况见图1所示。
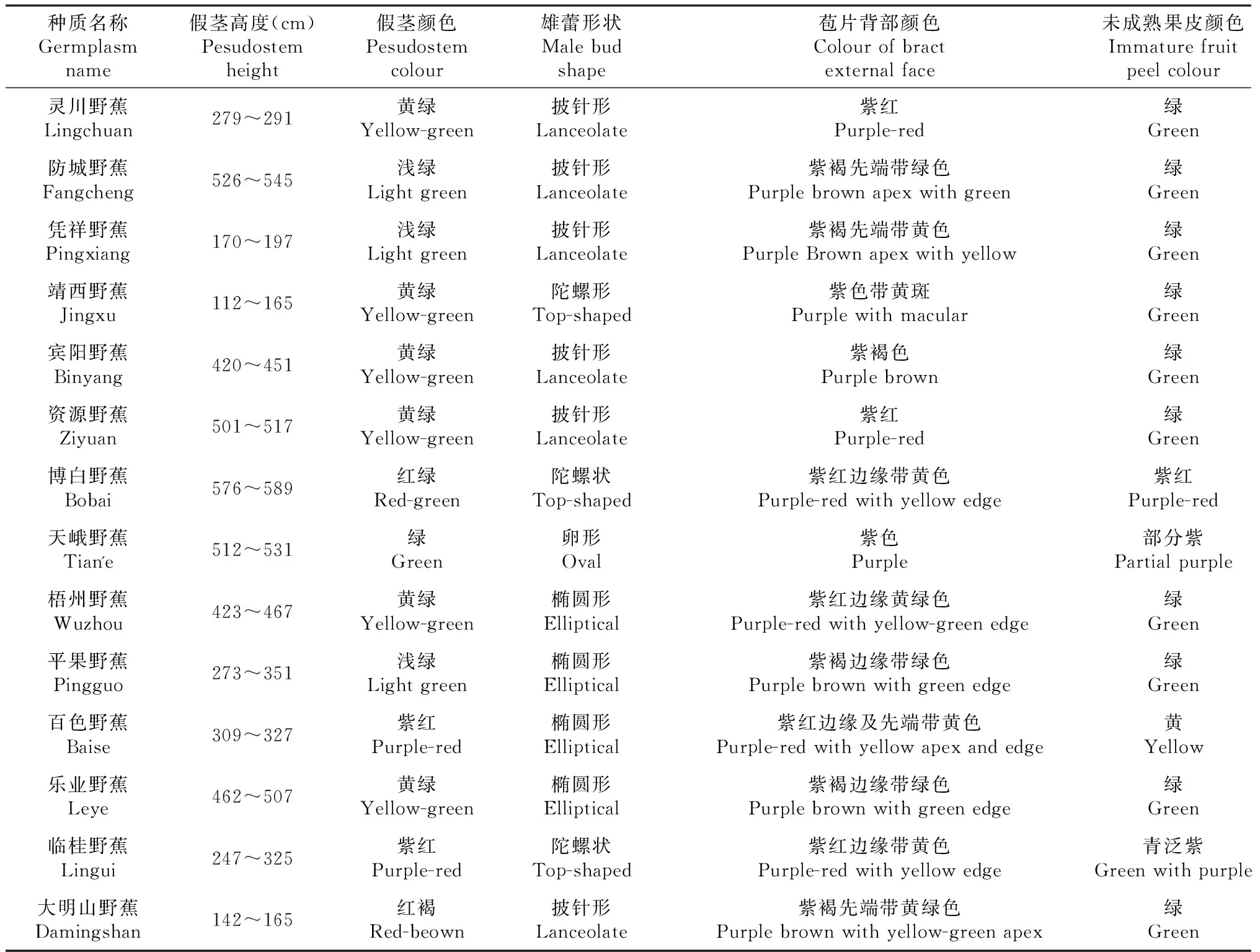
表3 广西野生蕉种质资源形态学描述部分性状
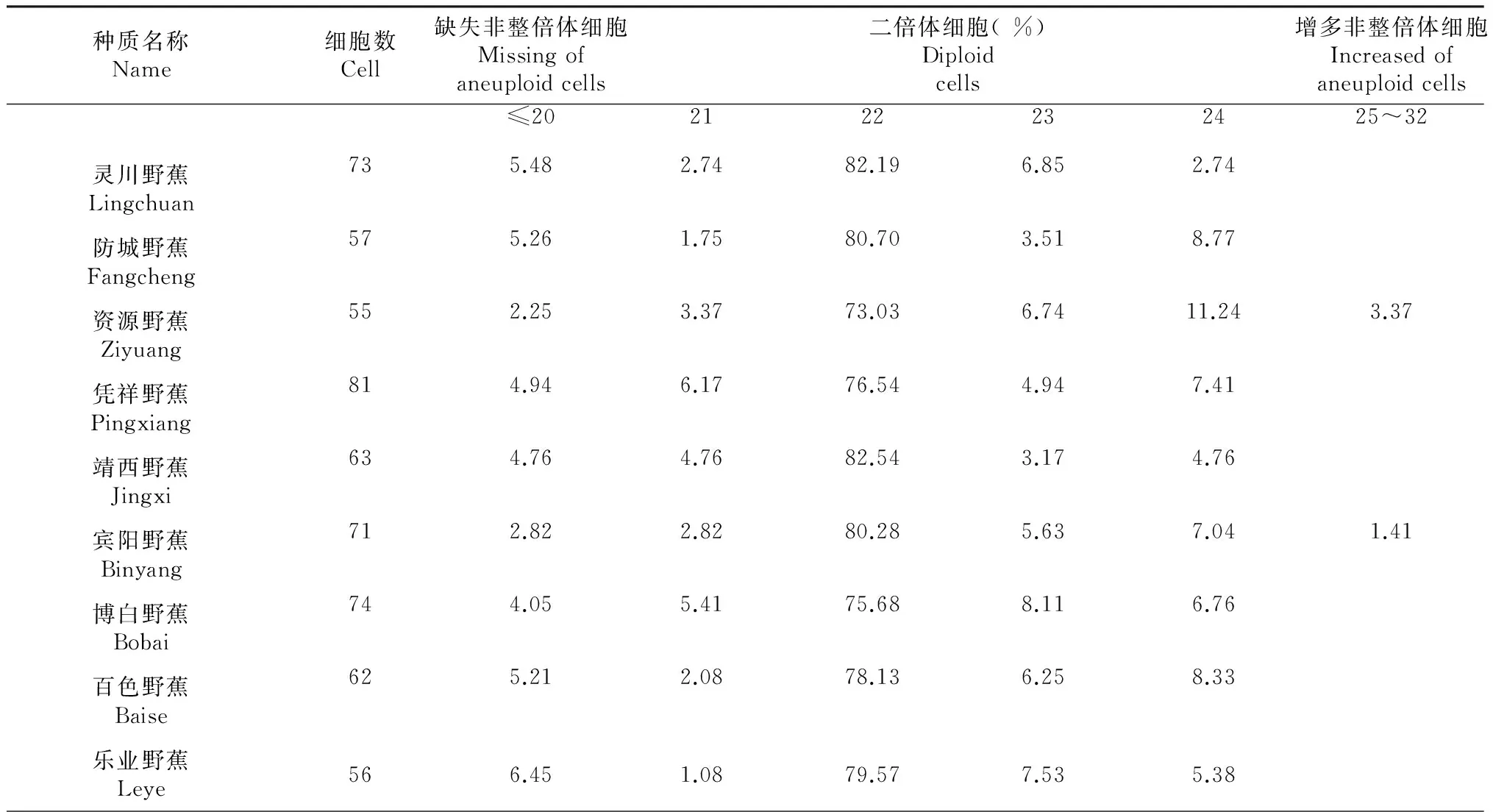
表4 供试野生蕉染色体数目不同的细胞百分比例(%)
续表4 Continued table 4

种质名称Name细胞数Cell缺失非整倍体细胞Missingofaneuploidcells二倍体细胞(%)Diploidcells增多非整倍体细胞Increasedofaneuploidcells临桂野蕉Lingui663.031.5286.364.554.55大明山野蕉Damingsha-n886.844.2773.505.987.691.71天峨野蕉Tian'e634.722.8378.305.666.600.94梧州野蕉Wuzhou934.302.1581.725.386.45平果野蕉Pingguo721.392.7881.948.335.56
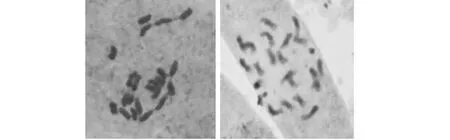
图1 染色体倍性为2n=22(x=11)的灵川野蕉(左)和凭祥野蕉(右)Fig.1 The Lingchuan (left) and Pingxiang (right) wild Musa chromosome ploidy are 2n=22(x=11)
2.3.2 广西野生蕉形态学鉴定 供试的14种广西野生蕉种质染色体倍性均为2n=22(x=11),根据Simmonds[9]分类法则中A-Y25个性状对14份广西野生蕉的性状鉴定归类到表5。形态性状归类结果表明本研究的14份广西野生蕉种质资源均属于芭蕉属(Musa)。

表5 供试野生蕉形态学性状归类
续表5 Continued table 5

材料名Name染色体数Chromosomenumbers各性状归类(A-Y根据表)Thetraitsclassified(A-Y)A/B/C/D/E/F/G/H/I/J/K/L/M/N/O/P/Q/R/S/T/U/V/W/X/Y所属的种Speices大明山野蕉Damingshan223/2/2/1/1/1/2//3/2/2/2/2/2/2/1/1/2/2//2/1/2/1/3/1/2M.intinerans天峨野蕉Tian'e223/1/3/1/1/2/2//2/3/2/2/2/1/2/1/1/2/3//2/1/2/1/3/1/2M.intinerans梧州野蕉Wuzhou223/1/3/1/1/2/2//2/3/2/2/2/1/2/1/1/2/3//2/1/2/1/3/1/2M.intinerans平果野蕉Pingguo223/1/3/1/1/2/2//2/3/2/2/2/1/2/1/1/2/3//2/1/2/1/3/1/2M.intinerans
(1)芭蕉属中组的分类。根据芭蕉属中组的分类系统,除凭祥野蕉、靖西野蕉和大明山野蕉外,其余野生资源均与分类法则一致,可确定为真蕉组(Eumusa),这3份野生蕉在分类检索的假茎高度性状处与形态学分类系统描述不一致,无法按法则进一步分类,其假茎高度均低于2 m(表3),且根状茎都小于1 m,但三者花序梗为水平或悬垂,基部花排列双列,因此兼有Eumusa组和Rhodoclamys组的形态特性,在对它们25(A-Y)个形态性状权重考量后,本研究认为与Eumusa更为相符,因此,将这3份野生蕉资源也归属为Eumusa组。
(2)野生蕉种的分类与鉴定。参照野生蕉真蕉组的分类标准,凭祥野蕉与靖西野蕉可归属为M.acuminatasubsp.malaccensis种;百色野蕉种的形态学分类鉴定中存在分类结果与客观不符的情况,按分类检索法则,百色野蕉因具有“果实成熟时开裂”的性状,可划归为M.schizocarpai种,但M.schizocarpa种的显著特征为具水绿色假茎以及雄蕾苞片绿色,与百色野蕉假茎黄绿色及雄蕾苞片紫褐色特征不一致。百色野蕉的25(A~Y)个形态学特征中除了果指开裂性状外,其余与M.itinerans种的形态特征高度相符,鉴于此,本研究认为其应归属为M.itinerans种更合理。大明山野蕉根状茎小于1 m,在真蕉组分类检索根状茎处与分类法则不一致。因根状茎大于1 m是真蕉组的主要特征,但在组的分类上,大明山野蕉是份比较特殊的种质,其兼有真蕉组和观赏蕉组的主要特征,综合考量了其它性状后将其归属于真蕉组,大明山野蕉根状茎性状在真蕉组中的分类权重应当弱化,依据其它主要形态学性状大明山野蕉在此也将其归属于M.itinerans种;其余野生蕉资源均与M.itinerans种分类性状相符(图2~3)。

图2 广西野生蕉形态标记分类图Fig.2 The dendrogram of Guangxi wild banans based on morphological traits

A:凭祥野蕉;B:靖西野蕉;C:大明山野蕉;D:百色野蕉A:Wild Musa from Pingxiang; B: Wild Musa from Jinxi; C: Wild Musa from Damingshan;D: Wild Musa from Baise图3 4份特殊广西野生蕉种质Fig.3 Four special wild banana germplasm in Guangxi
引物组合Primercombination总带数No.oftotalband多态性带数No.ofpolymorphicband多态性位点百分率(%)PercentageofpolymorphicbandAAC/CTA16115898.1AAC/CTG15415399.4AAC/CTT16616599.4ACT/CAA17316696ACT/CAC16416298.8ACT/CTG15114092.7ACT/CTT15715095.5ACG/CTG979496.9总计Total12231188平均Average152.9148.597.1
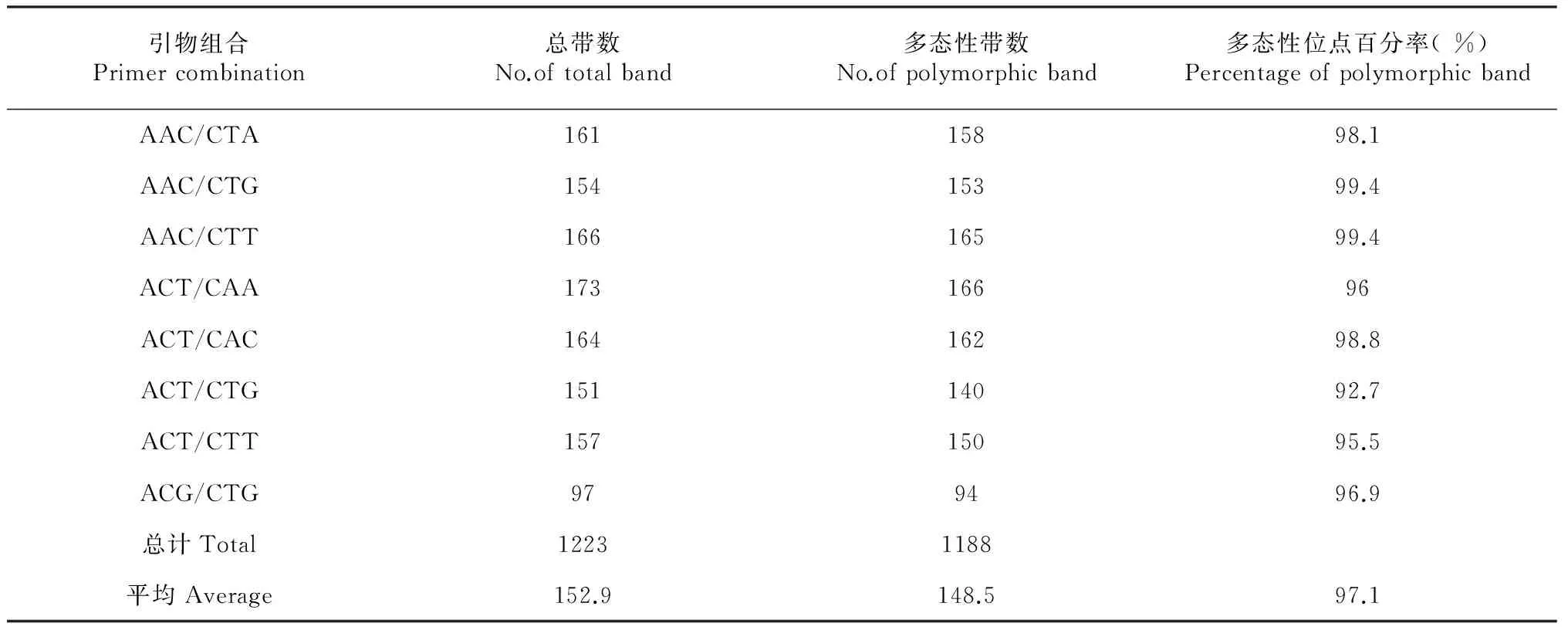
2.3.3 广西野生蕉种质AFLP分子标记鉴定 (1) AFLP多态性。本研究选取8对引物组合(表6)对25份蕉类种质进行AFLP分析,各份种质均获得较好的扩增效果,产生的谱带清晰,多态性高。引物组合AAC/CTG、AAC/CTT、AAC/CTA 和ACT/CAC扩增效果相对较好,其中引物对E-ACT/M-CAC的扩增结果见图4(自左而右1~30代表泳道编号,编号所代表的种质如表2所列)。8对引物平均多态性比率为97.1 %。特异条带占23.2 %, 可见,本研究供试蕉类种质的AFLP扩增谱带多态性非常高,充分体现供试蕉类种质间具有丰富的遗传多样性。
(2)供试香蕉种质的聚类分析。根据所选用的8对引物组合扩增后得到的1188条多态性条带进行亲缘关系和UPGMA聚类分析,获得25份供试香蕉材料的亲缘关系树状图(图5)。25个样品的成对相似系数在0.39~0.78。依阀值0.53的水平,可以将25份香蕉种质划分为4个类群。从树状图可以看出供试蕉类各类群之间的亲缘关系,相似系数越小表明其亲缘关系越远,供试广西野生蕉相互间相似系数比较低,介于0.39~0.65,说明广西野生蕉遗传多样性丰富外,种质间遗传差距较大。其中除了靖西野蕉、凭祥野蕉、灵川野蕉、博白野蕉和防城野蕉遗传距离较远外,其余广西野生蕉资源相似系数为0.58~0.65,亲缘关系相对较近。广西野生蕉与栽培蕉各类群在阀值0.41处相聚,表明它们的遗传距离非常明显。栽培蕉在树状图中被划分为两个类群,其中类群Ⅰ中包含1份AA基因型和5份AAA基因型的香牙蕉品种,相似系数为0.60~0.70,表明亲缘关系较近。类群Ⅲ包括1份长梗蕉和4份ABB基因型的粉蕉,类群内相似系数为0.62~0.78,遗传关系较近。但两个栽培蕉类群间在相似系数0.41处相聚,表明它们的遗传差异十分明显。依据不同阀值可将25份蕉类材料进一步细分,从树状聚类图可知,利用AFLP标记对蕉类种群的划分,跟前人的研究结论及本研究形态学标记分析结果基本一致,说明形态学标记分类系统具有一定的合理性。

图4 由引物对E-ACT/M-CAC扩增产生的荧光AFLP指纹图谱 Fig.4 The fluorescence AFLP fingerprint produced by primer combinations E-ACT/M-CAC
(3) 供试蕉类各种质亲缘关系分析。从聚类图(图5)可知,供试的14个广西野生蕉材料相互之间相似系数为0.39~0.65,在分子水平上体现了丰富的物种多样性。在相似系数0.53和0.54处分别划分为类群Ⅱ和类群Ⅳ两个类群,与其它栽培蕉类群区分明显。在类群Ⅱ中,仅有凭祥野蕉和靖西野蕉2份种质,其在阀值0.39处与其它广西野生蕉类群相聚,达到种一级分类单位差异的水平,在供试材料中亲缘关系最远。这与形态鉴定此两种野生蕉种质为M.acuminatasubsp.malaccensis种鉴定结果一致。在类群Ⅳ中,各野生蕉资源表现出丰富的多态性,遗传相似系数为0.53~0.65,聚类有趋向于来源地理相近分布位置的倾向。在阀值0.58处灵川野蕉、防城野蕉及博白野蕉分别独立成组,这3种野蕉种质在植物学形态特征上均有与其它野生蕉不同之处有关,如博白野蕉的幼果为青色,随着果实的发育,逐渐转为紫红色,成熟时整个果指都转为紫红色,成熟后颜色变化不大。灵川野蕉和防城野蕉叶柄基部对称且有稀疏的斑块,果序梗上毛少,苞片外表面红紫色,有黄色线,游离瓣片顶部有皱折,与其它野生蕉明显不同,表现出明显的形态差异性。在栽培蕉类群Ⅰ中,除了海贡蕉(Musa.AA)为AA基因组类型,其余均为AAA基因组型的香牙蕉品种, 它们都含有A基因组。在栽培蕉类群Ⅲ中,含有B基因组的长梗蕉(BB.M.balbisiana)和基因型为ABB类型的粉蕉的聚为一类,亲缘关系上与B基因组靠近。供试的14种广西野生蕉种质均有大量种子,它们与长梗蕉在相似系数0.42处相聚,分类差异明显,表明它们之间的亲缘关系较远。本试验电泳图谱中,泳道1,泳道2都是海贡蕉,泳道13,泳道14都是灵川野蕉,他们的AFLP指纹图谱完全一致(图4),充分说明本试验的AFLP分子标记结果重复性好,其所体现的聚类结果是可靠的。

图5 根据AFLP分子标记聚类分析产生的树状分支Fig.5 A dendrogram based on AFLP markers generated by UPGMA
3 讨 论
3.1 关于几份特殊种质的分类地位探讨
依芭蕉属分组特征,与靖西野蕉、凭祥野蕉和大明山野蕉染色体倍性相符的有Rhodochlamys组与Eumusa组。从形态学上看,它们假茎高度都在2 m以下、根状茎短的形态学性状与Rhodoclamys组相符;而花序梗呈悬挂,果指垂直于果穗,每个苞片间花多个、排成两列,苞片外表面无光泽、紫褐色等形态性状又与Eumusa相符。由于它们的形态性状与分类系统描述不一致,依照形态学分类方法显然无法将它们进行鉴别分组。鉴于这3种野生蕉的形态特性,或存在它们是两组间自然回交或渐进渗透的可能,也存在受生长环境影响而发生变异的可能。大明山野蕉长期生长在海拨1000 m以上的山上,存在为适应高海拨和低温影响而在假茎高度及根状茎上发生变异的可能。对于这些较为特殊的野生蕉种质,目前形态学分类并无标准的分类法则可依。根据Simmonds[9]对野生蕉的分类检索标准,假茎高度和花序梗着生位置处于同一权重水平,但蕉类假茎高度性状易受生长环境和生长年限的因素影响,而花序梗着生位置与果指排列是稳定遗传性状。本研究认为,在这3份野生蕉种质的分组形态学鉴定中应侧重考虑花序梗着生位置和果指排列的权重,并综合考量其它25(A-Y)个形态学性状。因此,本研究在态学分类上将这3份野生蕉归属为真蕉组(Eumusa)。
在AFLP亲缘关系分析中,靖西野蕉和凭祥野蕉在阀值0.55处聚为一类群,与其它阿宽蕉在阀值0.39处相聚,说明它们亲缘关系非常远。大明山野蕉与其它阿宽蕉在阀值0.62处相聚,在亲缘关系上与阿宽蕉较为接近,鉴定结果与形态学分类相符。这三种形态学特征介于真蕉组与美蕉组的野生蕉在AFLP标记中的分析结果支持了本研究的形态学鉴定结论。
关于百色野蕉的形态学鉴定,在本研究的前期研究中[19]因其具有“没有或很少蜡粉、果实成熟时开裂”的形态性状而依据形态学分类法则鉴定为M.schizocarpai种。M.schizocarpai种的显著形态特征是具水绿色假茎以及雄蕾苞片绿色,目前只在巴布亚新几内亚发现被报道过,与百色野蕉的显著形态学特征差异明显。在形态学性状分类中存在给哪个性状优先权的问题,对于形态学性状与形态学分类系统描述不一致的特殊种质,应当综合权衡其分类性状的优先权才能得出最接近客观的鉴定结果。如果仅因“没有或很少蜡粉、果实成熟时开裂”的形态性状将百色野蕉鉴定为M.schizocarpai种显然依据不够科学。在综合考量其它形态学性状后,本研究将其鉴定为M.itinerans种,该鉴定结论在AFLP分析聚类图(图5)中也得到了印证。
3.2 本研究中形态学标记分类与AFLP标记分析结果的比较
本研究中形态学标记鉴定结果表明形态学分类系统基本能将供试蕉类种质资源鉴定到种、个别亚种或组群的水平。但对于一些形态学性状与分类系统描述不一致的特异种质很难按形态学分类法则进行鉴定。野生蕉类由于生长环境等方面的影响,形态特征与遗传物质间并无严格的对应关系,且在形态学性状分类检索中存在给哪个性状优先权与权重衡量的问题。此时需要全面考量并对主要形态性状的优先权进行衡量取舍才能得出合理的鉴定结果。如依照分类法则对百色野蕉的鉴定结论显然与客观事实不符。此外,本研究结果发现形态学标记鉴定均为M.Itinerans种的野生蕉种质资源,在形态学特征上推测本应遗传关系较为相近,但在AFLP聚类图上显示它们的相似系数为0.53~0.65,可能原因是表型特征不能完全反映DNA水平差异,此现象与姜燕[20]相关研究观点一致。可见,形态学标记分类方法不足以完全明确蕉类种质资源的亲缘关系,且对一些特异的种质鉴别存在一定的不足。因此,形态学分类如能结合分子标记等分类手段确定形态学性状的权重,相互印证鉴定结果将更为准确。
AFLP分子标记在国内外蕉类种质资源的亲缘关系和分类鉴定中已取得广泛的成功应用。易干军等[11-12]应用AFLP技术对粉蕉、大蕉、龙牙蕉品种进行遗传多样性分析表明仅参照形态学分类方法简单归为AAB或ABB等类型不能反映品种之间的亲缘关系。WONG C etc[13]应用AFLP技术对芭蕉属下属的5个组间亲缘关系进行了分析,揭示了具有不同染色体数目的分类组间的多态性和亲缘关系。G Ude etc[14]对39份香蕉种质进行了AFLP遗传多样性和亲缘关系分析,研究结果表明AFLP不仅可以有效鉴定芭蕉属不同组间亲缘关系,也可以进一步将组内不同类群差异加以区分。本研究的AFLP分子标记在阀值0.53处将供试蕉类划分为4个类群,与香(大)蕉传统的形态、农艺形状分类基本一致,并能够反映基因组的相似性及杂交种之间的渗入程度。在广西野生蕉的亲缘关系分析上,用AFLP技术可以区分所有种质,并能揭示近缘品种的差异以及特殊种质的系统位置。如对百色野蕉、凭祥野蕉、靖西野蕉及其余阿宽蕉的亲缘关系和种内各份种质的差异是形态学鉴定无法达到的。因此,本研究认为AFLP分子标记技术应用于蕉类分类,相比于Simmonds分类系统,更能反映出种质间基因型的差异。蕉类形态学标记分类法借助于分子标记等分类手段互相印证才能更可靠的反映蕉类的亲缘关系与进化途径。
3.3 广西地区野生蕉类资源的遗传多样性和资源保护
广西壮族自治区属亚热带季风气候区,具有适合野生蕉类生长的光、温、水等自然条件。经考察发现,广西野生蕉类资源十分丰富。北到与湖南交界的资源县、灌阳县等地,南到防城、凭祥等地,西北部的天峨县及东部的贺州等地均发现了野生的蕉类资源。这些蕉类资源主要分布在含水量较为丰富的山麓及沟谷边, 地理条件比较恶劣,在进化过程中积累了较多的遗传变异,形态表现丰富,果皮色、叶柄沟槽、苞片形状和苞片皮外颜色等差异均较大,按果实和花蕾颜色可分为4个基本类型:青果红蕾、青果黄蕾、红果红蕾、黄果红蕾。本研究对14份广西野生蕉种质的AFLP标记结果表明广西野生蕉类具有非常高的遗传多样性,或含有抗寒、抗病虫等优异基因。但是,针对广西丰富的蕉类资源来说,目前的收集和研究还十分有限,仍需要进一步加强实地考察,收集更多的野生蕉资源,以建立完善的广西野生蕉种质资源圃,为广西野生蕉的亲缘关系与种质资源分类鉴定提供更多材料和利用依据。同时,随着野生蕉分布区的开垦和不合理开发利用,造成广西野生蕉面积不断缩减和种质流失,有的甚至成为濒危物种,因此,开展相关的收集保存和创新利用研究具有重要意义,应当引起必要的重视。
4 结 论
形态学分类研究结果表明供试的广西野生蕉均属于芭蕉属真蕉组(Eumusa),其中凭祥野蕉、靖西野蕉和大明山野蕉与形态学分类系统描述不完全一致,无法按法则进一步进行分类。本研究在综合考量这3种野生蕉的形态学性状及权重后,将凭祥野蕉和靖西野蕉归属为M.acuminatasubsp.malaccensis种,大明山野蕉归属为M.itinerans种,其余11种野生蕉可依分类法则鉴定为M.itinerans种,其在广西分布较普遍。AFLP鉴定结果表明广西野生蕉种质间遗传差异较大,具有丰富的遗传多样性,且与栽培蕉亲缘关系较远。
[1]冯慧敏,陈 友,邓长娟,等. 香蕉野生种质资源系统分类研究进展[J].热带农业科学,2011,31(5):38-44.
[2]尧金燕,彭宏祥,秦献泉,等. 广西蕉类种质资源概况及其育种创新利用前景[J].广西农业科学,2008,39(4):527-529.
[3]李锡文.云南芭蕉科植物[J].植物分类学报,1978,16(3):54-64.
[4]刘爱忠,李德铢.树头芭蕉的名实辩证[J].云南植物研究,2000,23(3):175-176.
[5]Hakkinen M.Musaitinerans(Musaceae) and its intraspecific taxa in China[J]. Novon,2008,18(1):50-60.
[6]秦献泉,彭宏祥,尧金燕,等.广西博白野生蕉植物学性状观察及分类地位[J].亚热带植物科学,2008,37(4):9-11.
[7]龙 兴,尧金燕,秦献泉,等.广西野生蕉花粉活力及贮藏特性研究[J].广西农业科学,2010,41(12):1325-1328.
[8]Cheesman E E. Classification of the Bananas:The GenusMusaL.[J]. Kew Bulletion,1947(2):106-117.
[9]Simmonds N W. The evolution of the Bananas[M]. London England: Tropical science series,Longmans,1962:532-560.
[10]Descriptors for Banana(Musa) [EB/OL],1996. http://www.bioversityinternational.org/fileadmin/user_upload/online_library/publications/pdfs/326.pdf
[11]易干军,余晓英,霍合强,等. 应用AFLP进行香牙蕉品种(系)的鉴别与分类[J].果树学报,2002,19(4):247-251.
[12]易于军,于晓英,霍合强,等. 粉蕉、大蕉和龙牙蕉的AFLP分类研究[J].园艺学报,2002,29(5):413-417.
[13]Wong C,Kiew R. Genetic diversity of the wild bananaMusaacuminata colla in Malaysiaas evidenced by AFLP[J].Annals of Botany,2001,88:1017-1025.
[14]Ude G,Pillay M,Nwakanma D,et al. Analysis of genetic diversity and sectional relationship inMusausing AFLP markers[J].Theoretical and Applied Genetics,2002,104(8):1239-1245.
[15]Wong C,Kiew R,Argent G,et al. Assessment of the validity ofthe sections inMusa(Musaceae) using AFLP[J]. Annals of Botany.2001,90:231-238.
[16]李懋学.植物染色体研究技术[M]. 哈尔滨:东北林业大学出版社,1991:13-14.
[17]陈豫梅.中国香蕉种质资源形态学与细胞学鉴定[D].华南农业大学硕士学位论文,2002: 13-14.
[18]易干军,于晓英,霍合强,等. 香蕉种质资源的AFLP鉴别与分类中DNA模板的制备[J].果树学报,2001,18(6):345-348.
[19]秦献泉. 广西野生蕉资源调查、分类及遗传多样性研究[D].广西大学硕士学位论文,2009: 33-34.
[20]姜 燕.中国主要蕉类形态学与分子系统学研究[D].西南农业大学硕士学位论文,2005:41-42.
(责任编辑 汪羽宁)
Investigation and Identification of Wild Musa Germplasm Resources in Guangxi Province
LONG Xing, QIN Xian-quan, FANG Ren, DENG Biao, AN Zhen-yu, HUANG Wei-xiong, YAO Jin-yan*
(Horticultural Research Institute,Guangxi Academy of Agricultural Sciences, Guangxi Nanning 530007,China)
【Objective】The wildMusagermplasms resources were roundly collected in Guangxi province and their genetic relationship and classification status were clarified to provide a theoretical basis for protection and utilization of the wildMusagermplasm.【Method】Based on chromosome tabletting, morphological taxonomy and AFLP labeled with fluorescence methods, genetic relationship of 14 wildMusagermplasms from different regions in Guangxi Province were investigated and studied.【Result】The morphological identification showed that the chromosome ploidy of the 14 wildMusagermplasms were diploid, 2n=22(x=11).2 of them were identified asM.acuminatasubspmalaccensis, and other 12 were identified asM.itinerans, The interspecific morphological traits exhibited relatively difference. The results of AFLP markers revealed a rich genetic diversity among the wildMusagermplasm in Guangxi province, which were consistent with morphological classification.【Conclusion】According to the morphological classification, all the 14 wildMusagermplasms belonged toEumusa. 2 of them were classified intoM.acuminatasubspmalaccensis, the other 12 were classified intoM.itineranswith wide distribution in Guangxi. The results of AFLP showed that significant difference existed among the genetic relationship of the wildMusagermplasms in Guangxi. And they had a distant relationship with culturalMusa.
WildMusagermplasm; Germplasm resource; Classification identification; Morphological marker; AFLP;Guangxi
1001-4829(2017)6-1284-10
10.16213/j.cnki.scjas.2017.6.008
2017-03-01
广西自然科学基金项目(2014GXNSFBA118099);广西农业科学院青年科技专项项目(桂农科2014YQ29);广西农业科学院基本科研业务专项团队项目(2015YT48)
龙 兴(1980-),男,广东电白人,硕士,助理研究员,研究方向为果树育种与栽培,*为通讯作者:尧金燕,E-mail:jinyan.yao@gxaas.net。
S668.1
A
