棉花MAGIC群体后代株系对赤霉素和缩节胺互作的生态响应及其敏感性鉴定
2017-08-08石治鹏林忠旭李晓方李定国
李 敏,石治鹏,林忠旭,李晓方,李定国*
(1.长江大学农学院作物遗传育种研究所,湖北 荆州 434025;2.长江大学园艺园林学院,湖北 荆州 434025;3.华中农业大学植物科学技术学院作物遗传改良国家重点实验室,湖北 武汉 430070)
棉花MAGIC群体后代株系对赤霉素和缩节胺互作的生态响应及其敏感性鉴定
李 敏1,2,石治鹏1*,林忠旭3,李晓方1,李定国1**
(1.长江大学农学院作物遗传育种研究所,湖北 荆州 434025;2.长江大学园艺园林学院,湖北 荆州 434025;3.华中农业大学植物科学技术学院作物遗传改良国家重点实验室,湖北 武汉 430070)
【目的】以70份棉花多基因型种群(MAGIC)后代株系为材料,研究株系主要性状对花铃期喷施缩节胺、缩节胺+赤霉素的生态响应,鉴定其敏感性。【方法】试验设缩节胺、缩节胺+赤霉素2个处理,每个处理1种浓度,即缩节胺(30 g/hm2,兑水225 kg/hm2)、缩节胺+赤霉素(30 g/hm2+0.5 g/hm2,兑水225 kg/hm2),以喷清水为对照。9月15日调查棉花株式图,测定单铃重、衣分、子指、理论皮棉产量。【结果】株高、5果枝长、10果枝长、顶部果枝长、单铃重、子指、皮棉产量较对照对缩节胺、缩节胺与赤霉素互作的响应达到极显著水平,与缩节胺处理相比,缩节胺与赤霉素互作对株高、果枝数、总果节、5果枝长、10果枝长的影响达到极显著水平,单株结铃数达到显著水平;除子指外,对其他10个指标均有促进作用。缩节胺与赤霉素互作可显著改变株高、果枝数、5果枝长和10果枝长的生态响应度,而对单株结铃数、单铃重、子指和皮棉产量的生态响应度无显著改变。基于表型生态响应度数据的聚类分析将群体分为3大类,并构建了3个材料池:钝感材料池、中间材料池、敏感材料池。【结论】在本试验条件下,棉花MAGIC群体后代株系性状对缩节胺、缩节胺与赤霉素互作产生了比较明显的响应,为生产中采用赤霉素缓和缩节胺对棉花的不良调控效应提供了依据。极端材料池的构建可以明确群体材料对环境的敏感性,为完善棉花生态响应机制奠定了基础。
棉花MAGIC群体;赤霉素;缩节胺;生态响应;极端材料池
【研究意义】多基因型种群品种是利用MAGIC(multi-parents advanced generation intercross)杂交等技术创造大量稳定的或者可精确繁殖的不同基因型重组体,形成基础群体,再选择表现型稳定一致,使其符合抗性、优质、高产、稳产和一致性等商品性目标,按照比例搭配成多基因型种群,进一步开展种群品种在生态区的评价,确定种群内各种基因型对生态环境的响应是否一致,进而获得更好的稳定性和丰产性[1]。棉花生态响应是指在田间生态条件的改变后,棉花群体的生理生育特性也随着变化,表现为敏感性和钝感性。如何在育种过程中了解材料对不同环境的敏感性并进行筛选和淘汰显得非常重要, 尤其是面对大量的育种材料。同时,激素与棉花的生育关系密切,外源激素和生长调节剂可以协调棉株体内激素的平衡,诱导棉株的正常发育[2-3]。可见,研究植物生长调节剂和外源激素及其互作对棉花生育的影响具有重要意义。【前人研究进展】有关缩节胺和赤霉素在棉花上的应用报道较多,其中缩节胺是赤霉素生物合成抑制剂[4],通过减少棉花内生赤霉素水平,从而抑制细胞的伸长和扩大,进而影响棉花的生长,塑造良好株型,改善生理特性、产量、纤维品质[5-10]。有关DPC对棉花产量及其构成因子调节效应的研究存在不一样的结果。Zhao等[11]和Pettigrew等[12]研究结果显示喷施DPC与对照比降低产量或无显著影响,但是徐新霞等[13]和Gwathmey等[14]结果表明喷施DPC可以提高单位面积结铃数和皮棉产量。而赤霉素是调节植物生长发育的重要植物激素之一,对植物的生长发育,如种子的萌发、茎的延展、叶片的生长以及植物的花和种子的发育等生理具有重要的调控作用[15-16]。棉花具有无限生长的特性,其生殖生长与营养生长的关系往往不平衡,根据作物补偿机制中的得失补偿和差异补偿[17],人们对生长抑制调节剂缩节胺在棉花生育生理中的作用进行了比较系统的深入研究,并在棉花生产中起到了巨大作用,而关于赤霉素与缩节胺互作对棉花生育的调节效应还未见报道。【本研究切入点】本试验在大田条件下叶面喷施缩节胺和赤霉素,研究缩节胺及其与赤霉素互作对棉花MAGIC群体后代株系生育的影响。【拟解决的关键问题】探讨棉花对缩节胺及其与赤霉素互作的生态响应,以便为棉花生长的调控、生态响应机制的完善和棉花育种提供理论依据。
1 材料与方法
1.1 试验材料
试验于2014年在长江大学农学院试验基地进行,土壤肥力中等,肥力均匀。供试材料为70份棉花MAGIC群体后代株系。播种期为2014年4月27日,移栽日期2014年5月16日,株距40 cm,行距1 m,每个小区面积为2 m2。本试验设置3个处理,处理Ⅰ:叶面喷施DPC,用量为30 g/hm2,兑水225 kg/hm2;处理Ⅱ:叶面喷施DPC+赤霉素,配合用量为30 g/hm2+0.5 g/hm2,兑水225 kg/hm2,处理III:以喷清水为对照。采用随机区组设计,每小区种1份材料,1行,每行5株,各处理2次重复。其他同一般大田管理。
1.2 试验方法
试验自棉花进入初花期后,分别于7月9日、7月18日、7月25日、8月6日、8月19日叶面喷施缩节胺,DPC+赤霉素。在9月15日开始调查棉花的株式图,包括株高、果枝数、果枝长、铃数和脱落数等;取样,室内考种,根据株数/667m2、单株成铃数和单铃重,计算理论皮棉产量。
1.3 生态响应度计算
为了表示生态响应试验结果,本试验提出了“生态响应度”这一概念。生态响应度是指一个群体或品种的各性状对不同环境因素调控的敏感程度的一种表示指标。计算公式如下:
根据各个性状的表型数据的平均值和生态响应度,数据采用DPS分析软件进行处理与分析,Excel2010作表。
2 结果与分析
2.1 棉花MAGIC群体后代株系主要性状对缩节胺与赤霉素互作的响应
表1为不同处理棉花MAGIC群体后代株系各性状均值的差异显著性分析。与对照相比,棉花MAGIC群体后代株系的株高、5果枝长、10果枝长、顶部果枝长、单铃重、子指、皮棉产量对缩节胺、缩节胺与赤霉素互作的响应达到极显著水平,果枝数、总果节对缩节胺的响应达到极显著水平且对缩节胺与赤霉素互作没有达到显著水平,单株结铃数和衣分对缩节胺、缩节胺与赤霉素互作的响应没有达到显著水平。与喷施缩节胺相比,棉花MAGIC群体后代株系的株高、果枝数、总果节、5果枝长、10果枝长对缩节胺与赤霉素互作的响应达到极显著水平,单株结铃数对缩节胺与赤霉素互作的响应达到显著水平,其他5个性状均没有达到显著水平,其中缩节胺与赤霉素互作除了对子指有降低的趋势外,对其他10个性状有提高作用。各性状的变异幅度和变异系数分析结果表明,缩节胺、缩节胺与赤霉素互作分别对株系性状的变异幅度既有增加作用也有减小作用;各项指标的变异程度较大,各性状均值变异系数介于5.40 %~43.15 %,其中3个处理的顶部果枝长和皮棉产量的变异系数都较高,衣分的变异系数最低。
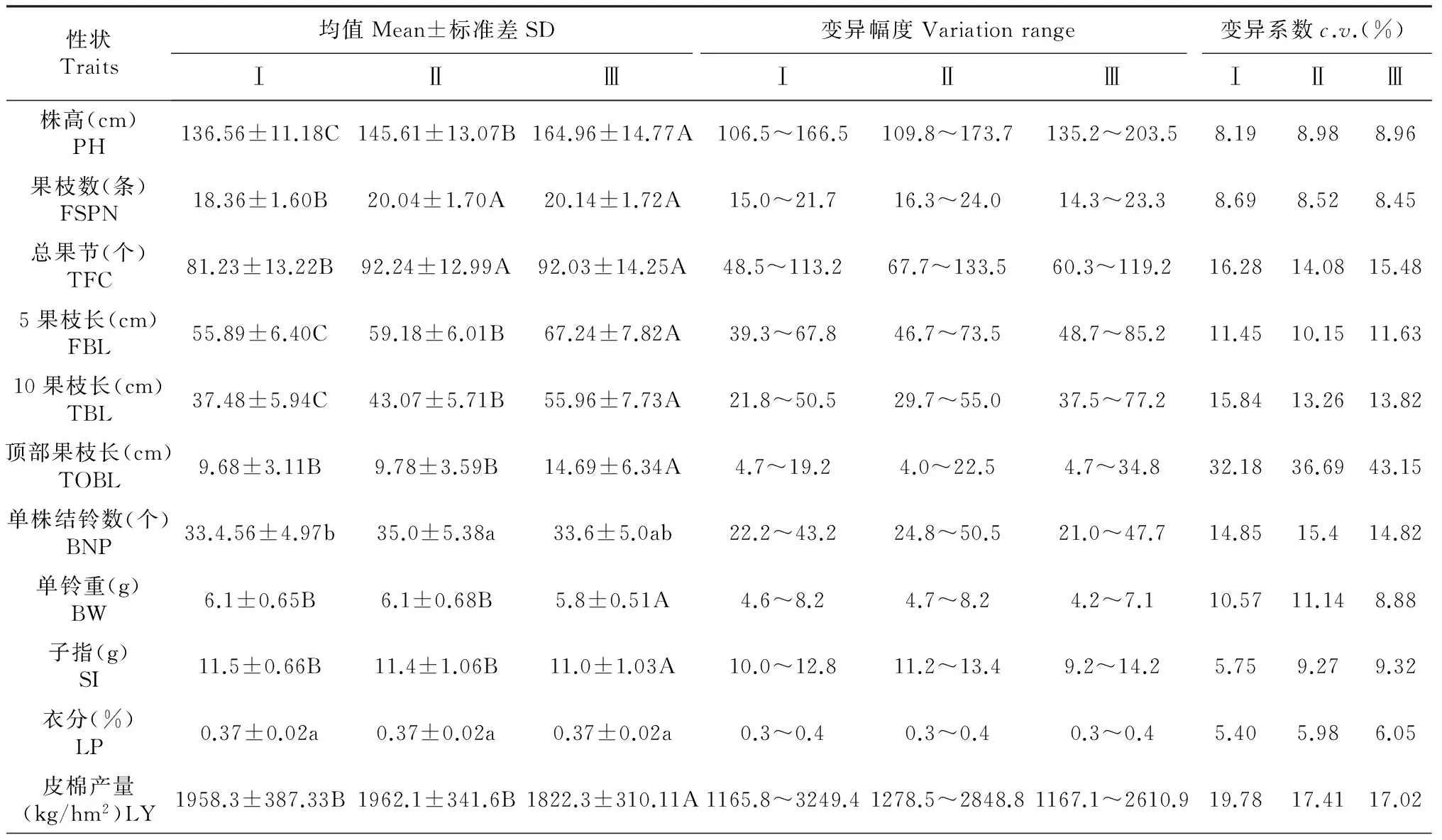
表1 棉花MAGIC群体后代株系主要性状对缩节胺与赤霉素互作的响应
注:同一行不同大写字母表示达到极显著差异(P﹤0.01);不同小写字母表示达到显著差异(P﹤0.05)。下同。 Note: Different capital letters in the same line indicate extremely significant difference at 0.01 level; The different small letters indicate significant difference at 0.05 level. The same as below. PH: Plant height; FSPN: Fruit section pitch number; TFC: Total fruit section; FBL: Fifth branch length; TBL: Tenth branch length; TOBL: Top of branch length; BNP: Boll number per plant; BW: Boll weight; SI: Seed index; LP: Lint percentage; LY: Lint yield.The same as below.
2.2 缩节胺与赤霉素互作对棉花MAGIC群体生态响应度的影响
生态响应度可以反映种群内各种基因型对生态环境的响应是否一致,也可以确定不同育种材料对不同环境的敏感性。计算各处理所考察的11个性状的生态响应度,根据响应度再进行差异显著性分析。表2结果显示,缩节胺与赤霉素互作对株高、10果枝长的响应度影响较缩节胺相比达到极显著水平,对果枝数、5果枝长的响应度影响较缩节胺相比达到显著水平,对总果节、顶部果枝长、单株结铃数、单铃重、子指、衣分、皮棉产量的响应度影响较缩节胺相比没有达到显著水平。缩节胺与赤霉素互作可以降低株高、果枝数、总果节、5果枝长、10果枝长、顶部果枝长、单株结铃数、单铃重、皮棉产量的生态响应度,可以提高子指、衣分的生态响应度。另外,不同处理各性状的生态响应度的变化幅度波动较大,缩节胺处理各性状的变化幅度较缩节胺与赤霉素互作相比较稳定。
2.3 棉花MAGIC群体后代株系主要性状生态响应度的相关分析
从表3可以看出,缩节胺处理后,皮棉产量与单株结铃数、单铃重、衣分呈极显著正相关,单铃重与子指、衣分呈正相关,单株结铃数与单铃重、衣分呈正相关,子指与衣分呈正相关,株高与果枝数、总果节、10果枝长呈极显著正相关,与5果枝长呈显著正相关,果枝数与总果节、10果枝长呈极显著正相关;株高与单株结铃数、单铃重呈负相关,与皮棉产量呈极显著负相关,总果节、5果枝长、10果枝长分别与单株结铃数、单铃重、皮棉产量呈负相关。赤霉素与缩节胺互作后,皮棉产量与总果节、单株结铃数、单铃重呈极显著正相关,衣分与子指呈极显著正相关,单株结铃数与单铃重、衣分呈正相关,单铃重与衣分呈正相关,株高与5果枝长呈显著正相关、10果枝长呈极显著正相关,株高与总果节、单株结铃数、单铃重、衣分呈负相关,与皮棉产量呈极显著负相关,果枝数与总果节、单株结铃数呈极显著正相关,总果节与单株结铃数呈极显著正相关,子指与株高、果枝数、总果节、顶部果枝长、单株结铃数、单铃重呈负相关。
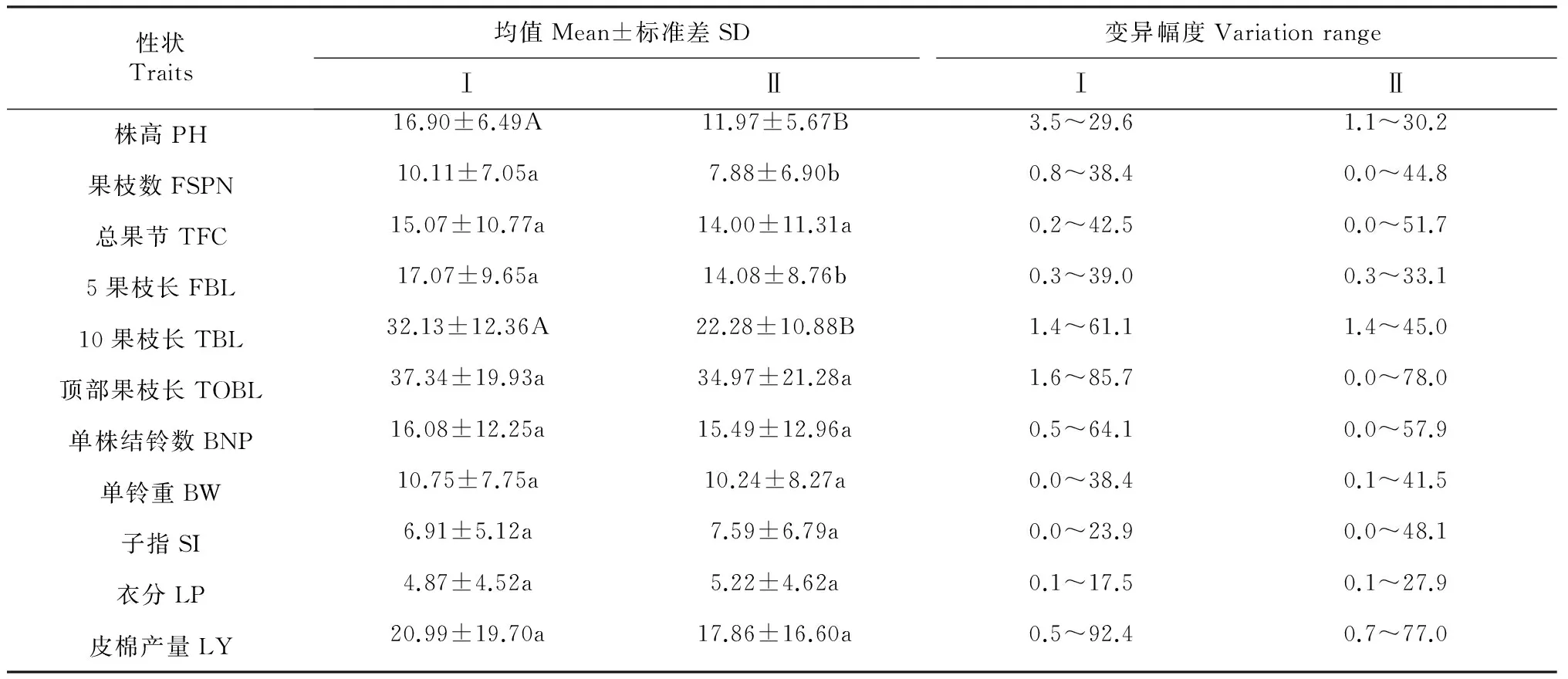
表2 棉花MAGIC群体生态响应度对缩节胺与赤霉素互作的响应
通过比较缩节胺及缩节胺与赤霉素互作两个处理的相关性,结果表明棉花MAGIC群体后代株系多数性状生态响应度间的相关性在2个处理下存在一致性,如皮棉产量与单株结铃数、单铃重、衣分呈极显著正相关,单铃重与衣分呈正相关,单株结铃数与单铃重、衣分呈正相关,株高与单株结铃数、单铃重呈负相关,与皮棉产量呈极显著负相关等;也有少数性状的相关性存在不一致性,如株高与果枝数在缩节胺处理后呈极显著正相关,而在缩节胺与赤霉素互作后呈负相关,籽指与单铃重在缩节胺处理后呈正相关,而在缩节胺与赤霉素互作后呈负相关等。
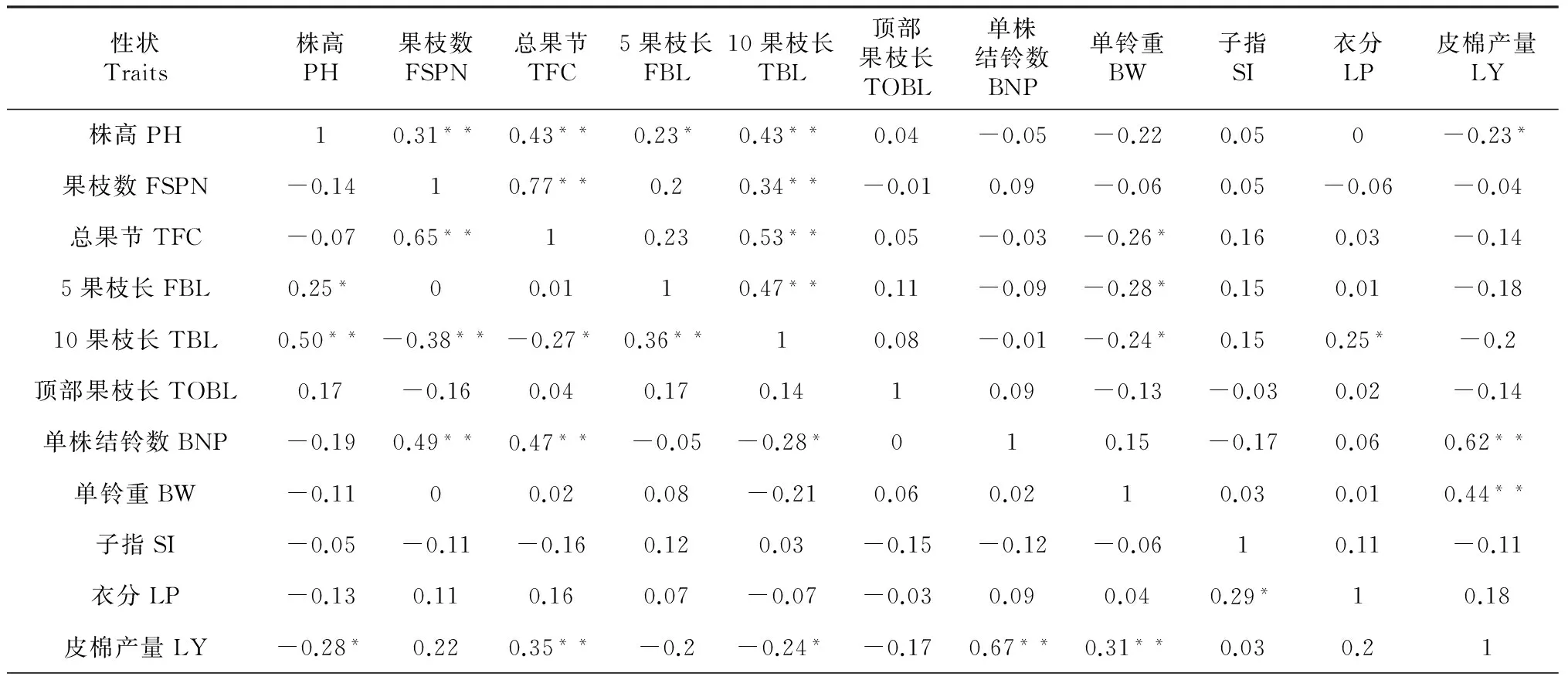
表3 棉花MAGIC群体后代株系主要性状生态响应度的相关分析
注:表中右上角为缩节胺处理,左下角为赤霉素与缩节胺处理;*和**分别表示在0.05水平和0.01水平上显著相关。 Note: Table top right corner for DPC treatment, table lower left corner for interaction of DPC and gbberelin treatment; * and ** represent significance at 0.05 and 0.01 levels.
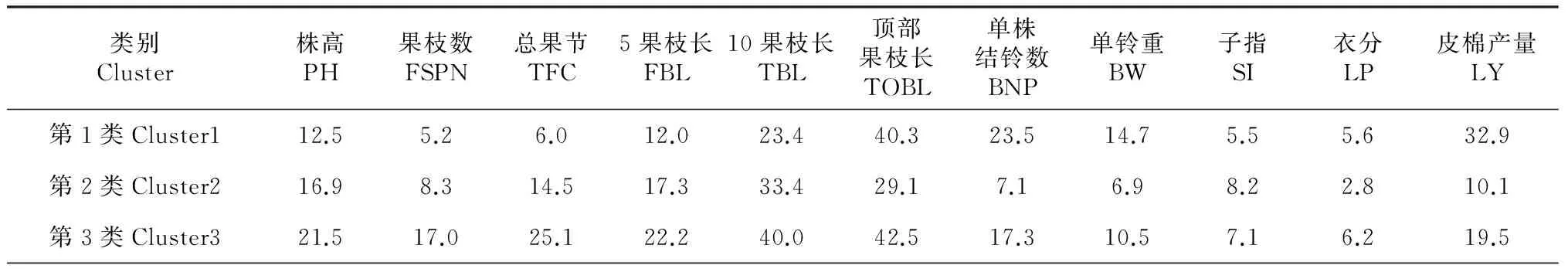
表4 3个类群棉花MAGIC群体后代株系11个性状的生态响应度的平均值
2.4 棉花MAGIC群体后代株系对缩节胺的敏感性鉴定
利用上述缩节胺处理棉花MAGIC群体后代株系的11个性状得到的生态响应度,将数据标准化转换后,依据欧氏距离采用离差平方和法,进行系统聚类分析。结果表明,当欧氏距离在14.38的水平上时可以把70份试验材料划分成3类(图1),其中3个不同类群的各个性状的响应度的平均值见表4。第1类群包括24份材料,其特点是株高、果枝数、总果节、5果枝长、10果枝长、子指对缩节胺的响应度最低,单株结铃数、单铃重、皮棉产量对缩节胺的响应度最高,顶部果枝长、衣分对缩节胺的响应度居中,综合表现为第1类群对缩节胺响应钝感,可以将该类群划分为钝感材料池。第2类群包括23份材料,其特点为株高、果枝数、总果节、5果枝长、10果枝长对缩节胺的响应度居中,顶部果枝长、单株结铃数、单铃重、衣分、皮棉产量对缩节胺的响应度最低,子指对缩节胺的响应度最高,综合表现为第2类群对缩节胺响应属于中间类型,可以将该类群划分为中间材料池。第3类群包括23份材料,其特点为株高、果枝数、总果节、5果枝长、10果枝长、顶部果枝长、衣分对缩节胺的响应度最高,单株结铃数、单铃重、子指、皮棉产量对缩节胺的响应度居中,综合表现为第3类群对缩节胺响应敏感,可以将该类群划分为敏感材料池。
3 讨 论
塑造理想株型,防止疯长,构建合理群体,是棉花高产稳产的基础。棉花的缩节胺化控技术要求很高,浓度、时期的不同,缩节胺化控对棉花的形态、产量、纤维品质等性状的影响会不同;不同棉花品种对缩节胺化控的敏感性也存在差异。缩节胺是内吸性的抑制型植物生长调节剂,通过减少赤霉素合成,抑制细胞的增加和扩大[4],而赤霉素最显著的生理效应就是促进植物的生长发育,主要是通过细胞数目的增加和细胞的伸长来完成[18],对棉花的种子发育[19]、叶片生长[3]、棉铃发育及产量[20]等影响较大。本研究结果表明,缩节胺对株高、果枝数、总果节、果枝长有抑制作用,对单铃重、子指、皮棉产量有促进作用,对衣分没有影响,这与前人研究结果一致[6,14,21-22]。但是缩节胺对单株结铃数略有降低,这与徐新霞[13]等、Gwathmey等[14]结果不一致,这可能与试验群体材料的复杂性及缩节胺喷施剂量有关系。缩节胺与赤霉素互作对株高、果枝数、5果枝长、10果枝长、顶部果枝长有抑制作用,对总果节、单株结铃数、单铃重、子指、皮棉产量有促进作用,说明棉花MAGIC群体后代株系对缩节胺及缩节胺与赤霉素互作的响应比较明显。在试验过程中,在不同时期喷施缩节胺过量时,可以用赤霉素与缩节胺配合使用,在一定程度上可以对缩节胺的调节效应起到正向的缓和作用。2个处理对各性状的变异幅度和变异系数既有增加作用也有减少作用,变化幅度较大,说明棉花MAGIC群体后代株系类型丰富,各具特点,差异明显,所考察的主要性状对缩节胺及缩节胺与赤霉素互作的响应较为敏感。另外,与缩节胺处理相比,缩节胺与赤霉素互作可以显著改变部分农艺性状(株高、果枝数、5果枝长和10果枝长)的生态响应度,而对产量性状(单株结铃数和皮棉产量等)的生态响应度无显著改变,说明在2种环境下农艺性状生态响应度的一致性没有产量性状生态响应度稳定。不同处理各性状的生态响应度的变化幅度波动较大,缩节胺处理各性状的变化幅度较缩节胺与赤霉素互作相比较稳定。
高产一直是棉花育种家们追求的目标,而影响棉花产量的性状较多,且性状间存在着相关性[23-24]。在田间性状选择时,应该注意各性状间相互制约的复杂关系,在争取改良一个性状的同时,也要防止另一个性状的劣变。通过对对棉花MAGIC群体后代株系主要性状生态响应度的相关分析,结果表明缩节胺、缩节胺与赤霉素互作对棉花MAGIC群体后代株系多数性状生态响应度间的相关性有较大影响,在2种环境下性状的相关性既存在一致性也有不一致性,说明不同环境对性状间的相关性有影响;同时获得的大量极显著(显著)相关的表型性状指标,如皮棉产量与单株结铃数、单铃重、衣分呈正相关,单株结铃数与单铃重、衣分呈正相关等,了解了性状间的相互关系,这为评价棉花MAGIC群体生态响应的一致性提供了参照。
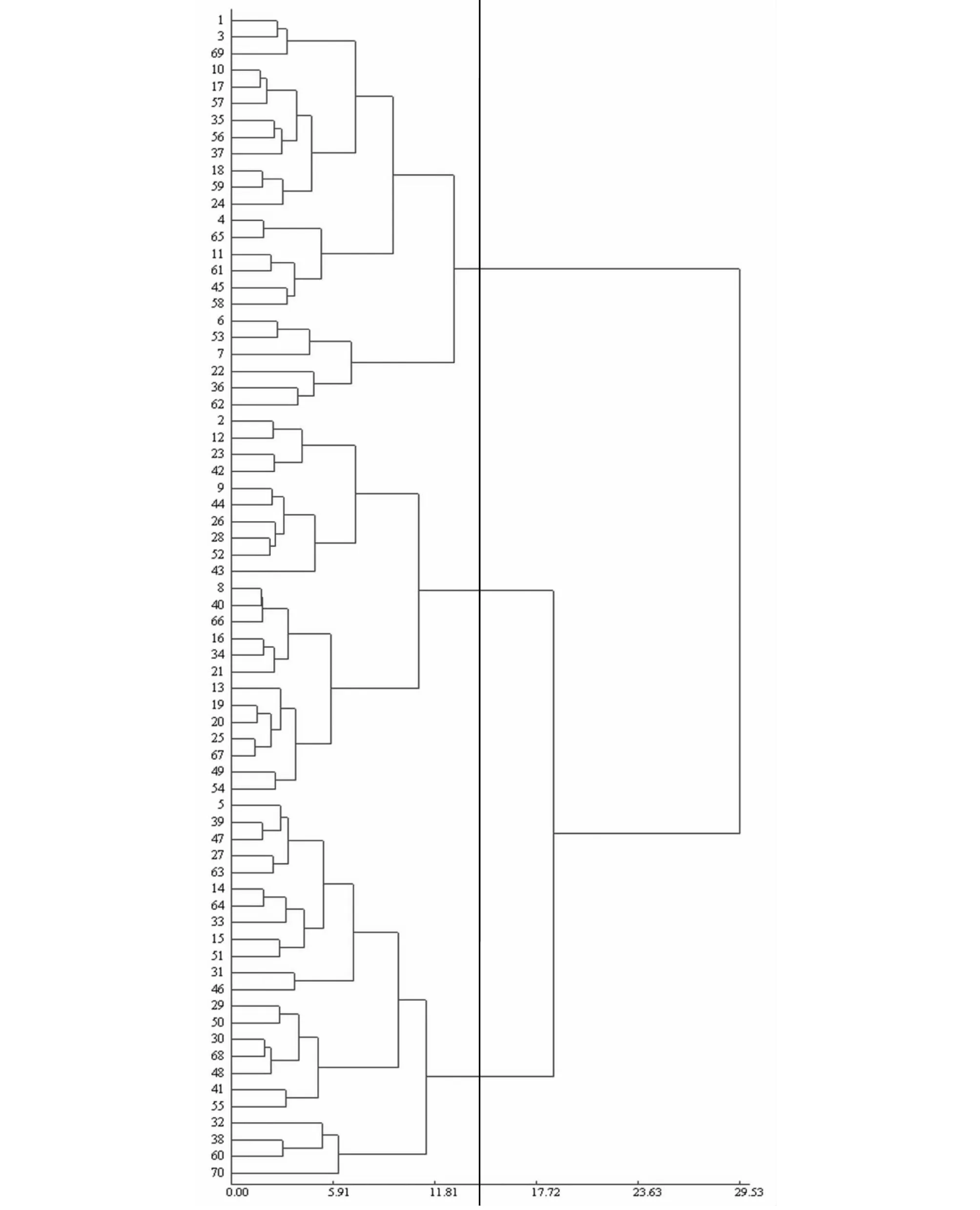
图1 棉花MAGIC群体后代株系各性状的聚类分析Fig.1 Cluster analysis of the traits for the descendant lines of cotton MAGIC population
聚类分析的依据是遗传距离,遗传距离大的品种分布在不同的类群,遗传距离小的品种分布在同一类群。聚类分析在棉花[25]、水稻[26]、小麦[27]等作物品种(系)资源的差异和分类方面,证明是比较可行的分析评价方法。本研究对经过缩节胺处理的棉花MAGIC群体后代株系11个性状的生态响应度进行聚类分析,结果表明,根据棉花MAGIC群体后代株系对缩节胺响应的敏感性,可以划分为3个材料池:钝感材料池(24份材料)、中间材料池(23份材料)、敏感材料池(23份材料)。极端材料池的构建,可以为棉花提供一些特异的育种材料。生态响应钝感性是决定品种稳产的关键因素,在育种工作中,选育钝感材料, 一定程度上与产量具有同等的重要性,甚至在产量水平已经很高的情况下,稳产和广适性的品种具有更加重要的意义。一个品种是在特定的环境下育成的,只能适应一定的生态区域和栽培条件。所以,在不同区域,缩节胺化控技术在棉花群体材料中的应用是未来棉花育种的一个重要趋势。
植物生长调节剂和植物激素对棉花生育的调节效应十分复杂,受环境条件影响很大。本文只对适宜的单一浓度进行缩节胺、赤霉素与缩节胺配合喷施,对棉花MAGIC群体后代株系性状的调控作用进行了比较研究,对于缩节胺对棉花体内的赤霉素含量影响,缩节胺与赤霉素互作机制以及棉花体内相关激素是否参入调节还有待于进一步研究。
4 结 论
在本试验条件下,棉花MAGIC群体后代株系性状对缩节胺、缩节胺与赤霉素互作产生了比较明显的响应,缩节胺与赤霉素的配合使用可以用来缓和或调节缩节胺因过量对棉花产生的不良调控效应。极端材料池的构建可以明确群体材料对环境的敏感性,为下一步进行不同生态区的棉花生态响应试验和完善生态响应机制奠定了基础。
[1]李晓方.农作物多基因型种群育种及种子生产技术体系[M]. 北京:科学出版社,2012:37-49.
[2]郑莎莎,孙传范,孙红春,等. 不同外源激素对花铃期棉花主茎叶生理特性的影响[J]. 中国农业科学,2009,42(12):4383-4389.
[3]王 丽,张明才,杜明伟,等. 赤霉素生物合成酶基因GhCPS和GhKS参与甲哌鎓对棉花幼苗叶片生长的控制[J]. 作物学报,2014,40(8):1350-1355.
[4]Rademacher W. Growth Retardants: effects on gibberellin biosynthesis and other metabolic pathways[J]. An nu. Rev. Plant Physiol, 2000, 51: 501-531.
[5]Li Wang, Chun Mu, MingweiDu,et al. The effect of mepiquat chloride on elongation of cotton(GossypiumhirsutumL.) internode is associated with low concentration of gibberellic acid[J]. Plant Science, 2014, 225: 15-23.
[6]杨长琴,张国伟,刘瑞显,等. 种植密度和缩节胺调控对麦后直播棉产量和冠层特征的影响[J]. 棉花学报,2016,28(4):331-338.
[7]Gonias E D, Oosterhuis D M, Bibi A C. Cotton radiation use efficiency response to plant growth regulators[J]. Agric Sci, 2012, 150: 595-602.
[8]杨成勋,张旺锋,徐守振,等.喷施化学打顶剂对棉花冠层结构及群体光合生产的影响[J].中国农业科学,2016,49(9):1672-1684.
[9]Fuqiang Yang, Mingwei Du, XiaoliTian, et al. Plant growth regulation enhanced potassium uptake and use efficiency in cotton[J]. Field Crops Research,2014,163: 109-118.
[10]冯国艺,罗宏海,姚炎帝,等. 新疆超高产棉花叶、铃空间分布及与群体光合生产的关系[J]. 中国农业科学,2012,45(13):2607-2617.
[11]Zhao D L, Oosterhuis D M. Pix plus and mepiquat chloride effects on physiology, growth, and yield of field-grown cotton[J]. Plant Growth Reg, 2000,19(4): 415-422.
[12]Pettigrew W T,Johnson J T. Effects of different seeding rates and plant growth regulators on early-planted cotton[J]. Journal of Cotton Science, 2005(9): 189-198.
[13]徐新霞,苏丽丽,魏 鑫,等. DPC对杂交棉生长发育调控效应研究[J]. 新疆农业科学,2015,52(7):1237-1242.
[14]Gwathmey C O, Clement J D. Alteration of cotton source-sink relations with plant population density and mepiquatchloride[J]. Field Crops Research, 2010,116: 101-107.
[15]Olszewski N, Sun T P, Gubler F. Gibberellin signaling: biosyn thesis, catabolism and response pathways[J]. Plant Cell, 2002, 14: 61-80.
[16]Ogawa M, Hanada A, Yamauchi Y, et al. Gibberellin biosynthesis and response during Arabidopsis seed germination[J]. The Plant Cell, 2003, 15: 1591-1604.
[17]赵 明,李建国,张 宾,等.论作物高产挖潜的补偿机制[J]. 作物学报,2006,32(10):1566-1573.
[18]Yamaguchi S. Gibberellin metabolism and its regulation[J]. Annu Rev Plant Biol, 2008, 59: 225-251.
[19]陈云风. 赤霉素和NaCl对棉花种子萌发的影响研究[J]. 种子,2015,34(11):97-99.
[20]何俊彦,顾静文,桂兰芳. 赤霉素对棉铃及种子发育的影响[J]. 作物学报,1983,9(3):212-214.
[21]Lili Mao, Lizhen Zhang, Xinhua Zhao, et al. Crop growth, light utilization and yield of relay intercropped cotton as affected by plant density and a growth regulator[J]. Field Crops Research, 2014, 155: 67-76.
[22]宋 妮,孙景生,陈智芳,等. 不同水分状况下化控对棉株蕾铃数和产量品质的影响[J]. 干旱地区农业研究,2015,33(2):40-49.
[23]苏桂兰,金石桥,赵淑琴,等. 中棉所49及其亲本中棉所35农艺及产量性状的通径分析[J]. 中国棉花,2014,41(12):26-29.
[24]宿俊吉,邓福军,陈 红,等. 陆地棉主要农艺性状的变异性、聚类和相关性分析[J]. 新疆农业科学,2011,48(8):1386-1391.
[25]李 武,谢德意,赵付安,等.黄河流域棉花品种农艺性状的主成分及聚类分析[J]. 江苏农业科学,2015,43(8):82-85.
[26]游书梅,曹应江,郑家奎,等. 73 份亚洲水稻恢复系农艺性状的主成分与聚类分析[J]. 植物遗传资源学报,2015,16(2):250-256.
[27]周丽艳,郭振清,马玉玲,等. 春小麦品种农艺性状的主成分分析与聚类分析[J]. 麦类作物学报,2011,31(6):1057-1062.
(责任编辑 陈 虹)
Ecological Response and Sensitive Identification of Descendant Lines of Cotton MAGIC Population to Interaction Effect of DPC and Gibberelin
LI Min1,2, SHI Zhi-peng1*, LIN Zhong-xu3, LI Xiao-fang1, LI Ding-guo1**
(1.Institute of Crop Genetic and Breeding,College of Agriculture, Yangtze University, Hubei Jingzhou 434025,China;2.College of Horticulture and Gardening,Yangtze University, Hubei Jingzhou 434025, China; 3.National Key Laboratory of Crop Genetic Improvement, College of Plant Science & Technology of Huazhong Agricultural University, Hubei Wuhan 430070,China)
【Objective】Using 70 cotton multi-parent advanced generation inter-cross (MAGIC) as tested materials, the ecological response of DPC spraying, combined of DPC and Gibberelin spraying during the flowering and boll-setting stage on cotton agronomic and yield-related traits were investigated.【Method】Two treatments with one concentration levels each, i.e. DPC 30 g/hm2and watering 225 kg/hm2, combined of DPC 30 g/hm2and Gibberelin 0.5 g/hm2, watering 225 kg/hm2were employed, and with spraying water as the control(CK). Cotton plant types were surveyed on September 15, and their single boll weight, lint percentage, seed index and lint yield were measured. 【Result】Compared with the controls, extremely significant response to DPC, combined of DPC and Gibberelin was observed in DPC-applied,combined of DPC and Gibberelin-applied cotton in terms of plant height, length of the 5th fruit branch, length of the 10th fruit branch, length of the top fruit branch, single boll weight, seed index,lint yield. Compared with DPC, extremely significant response to combined of DPC and Gibberelin was observed in combined of DPC and Gibberelin-applied cotton in terms of plant height, fruit branch number, total boll-nodes, length of the 5th fruit branch, length of the 10th fruit branch, significant response to combined of DPC and Gibberelin was observed in terms of boll number per plant. In addition to the seed index, the other 10 indicators were promoted. Significant change of ecological response degree of plant height, fruit branch number, length of the 5th fruit branch, length of the 10th fruit branch was observed by DPC and Gibberellin interaction, but the change of ecological response degree of boll number per plant, single boll weight, seed index, lint yield were not significant. Subject to the cluster analysis based on phenotype responsivity data, 70 descendant lines of cotton MAGIC population were divided into 3 categories, and three material pools were finally constructed by reference to the screening results, including the sensitive material pool, theintermediate material pooland the insensitive material pool. 【Conclusion】Under this experimental condition, the ecological response of DPC spraying, combined of DPC and Gibberelin spraying on cotton agronomic and yield-related traits, were sharped, which could be used for reducing adverse regulatory effect by DPC. The construction of extreme material pools can clear the sensitivity of the population response to the environment, and lay the foundation for improving the ecological response mechanism of cotton.
Cotton MAGIC population; Gibberelin; DPC; Ecological response; Extreme material pools
1001-4829(2017)6-1268-07
10.16213/j.cnki.scjas.2017.6.005
2016-11-10
国家公益性行业(农业)科研专项(201303008);国家自然基金(37371674)
李 敏(1992-),女,湖北广水人,硕士,主要从事棉花遗传育种研究,E-mail:cjdxlimin@163.com,Tel:13797333375,*为共同第一作者:石治鹏(1991-),男,湖北孝感人,硕士,主要从事棉花遗传育种研究,E-mail:cjdxshizhipeng@163.com,Tel:13872226051,**为通讯作者:李定国,E-mail:361113214@qq.com。
S562
A
