老龄林林分空间的点格局分析1)
2017-08-07许文秀林文树吴金卓
许文秀 林文树 吴金卓
(东北林业大学,哈尔滨,150040)
老龄林林分空间的点格局分析1)
许文秀 林文树 吴金卓
(东北林业大学,哈尔滨,150040)
采用TOPCON全站仪,在吉林省蛟河市林业实验区管理局施业区内设置1块500 m×600 m固定监测样地,并进行群落调查,在ArcGIS平台上采用Ripley’s K函数进行不同尺度的空间聚集度研究,采用空间自相关方法对研究区内的种群进行空间自相关性分析。结果表明:老龄林中数量最多的前6个种群分别是簇毛槭、白牛槭、千金榆、色木槭、裂叶榆和花楷槭;不同的起测胸径对林木的空间格局分布有重要的影响,起测胸径5 cm时,发现6个个体数量最多的种群显示出了不同的空间格局,其中白牛槭的空间聚集性最高;优势木在空间尺度较小时,生长得比较密集,在空间尺度较大时,则呈现出离散的分布特点,不同的起测胸径对研究结果也有一定的影响,因此,在研究空间格局分布时,起测胸径是首先需要考虑的因素;优势木在种群间还是不同空间距离下,优势木间均显示正向的空间自相关性,只是相关性程度略有差异,优势木的相关性受空间尺度的影响较大。
Ripley’s K;点格局分析法;空间聚集度;空间自相关性;优势木
Ripley’s K; Point pattern analysis; Spatial clustering; Spatial autocorrelation; Dominant trees
森林中最初的植物种群格局与种群结构、微生物环境、光照强度、种群竞争、单木生长状况的综合作用,决定了森林的种群空间分布格局[1]。对种群空间分布格局的研究,是为了进一步研究每个种群的分布特征以及种群与种群之间、种群与环境之间的相互作用机制。植物种群的空间分布在水平空间上,一般呈现出随机分布、均匀分布和集群分布等[2-3],而种群空间分布格局类型与空间尺度有着十分密切的联系[4]。在不同的空间尺度下,种群的空间分布格局也可能有较大的差别。例如,在小尺度下某个物种可能呈现出聚集分布,然而在更大的空间尺度下,空间分布类型可能变为随机分布或者是均匀分布。因此,在大面积固定监测样地的基础上,通过探索不同的空间尺度下的种群分布空间格局,才能更加有效地分析种群分布格局在一定尺度下的稳定性。
空间格局的分析方法一般可分为样方法、距离法和角尺度法[5-6],其中,距离法使用的Ripley’s K函数是分析种群空间分布格局最常用的方法。Ripley’s K函数是点密度距离函数,它是由著名的统计学家Ripley于1977年首次提出,并应用到点格局分析中[7-9]。由于Ripley’s K函数能够有效地避免尺度效应的难题,减少数理统计中庞大的计算量,因此,在国内外林木空间格局分析中得到了广泛的应用。例如,P. Haase[10]介绍了应用Ripley’s K函数需要掌握的获取野外数据的步骤、统计分析过程以及如何进行结果检验。Druckenbrod et al[11]应用Ripley’s K函数检验了美国弗吉尼亚山麓地带7 512株活立木和枯死木在1~25 m半径范围内的林木空间分布格局,结果表明,弗吉尼亚山麓地带的林分空间格局是由于树木间竞争造成的,包括由不同地点的土壤湿度以及光照的可获得性驱动的种群生活史。王国严等[12]利用Ripley’s K函数进行点格局分析,研究了雅鲁藏布江砂生槐树的种群空间分布类型,指出砂生槐的种子在当地恶劣的自然条件下,发芽率和幼苗成活率都较低,一旦个体成长到一定大小,则抗逆和生存能力得到较大提高,种群以中等大小个体最多。张金屯[4]利用点格局分析法,分析了美国密歇根州克林顿县栎林3个优势种的格局,结果表明,点格局分析的结果比传统的方法更接近实际,在群落结构研究中有重要意义。尤海舟等[13]将点格局分析和其他格局分析方法进行了比较,指出了点格局分析方法在格局分析中的重要地位,并对点格局分析的研究趋势进行展望。综上可知,基于Ripley’s K函数的点格局分析在研究中得到了广泛的应用,但是在大面积样地中应用的有关文献则很少。本文以吉林省蛟河市林业实验区管理局的30 hm2老龄林为研究对象,分析不同树种在不同的空间尺度下的空间格局变化情况、不同起测胸径对空间格局变化的影响以及优势木的空间分布格局,最后,对优势树种之间的空间关联性进行分析。研究结果可为群落的空间格局分布和物种共存机制研究以及森林保护与可持续经营提供理论依据和参考。
1 研究区概况
研究区位于吉林省蛟河市林业实验区管理局林场(127°35′~127°51′E,43°51′~44°5′N),海拔593~803 m。该管理局位于长白山系张广才岭西南坡余脉,东邻延边、北接龙江、西南为蛟河。辖区内总经营面积31 823 hm2,林业用地面积28 646 hm2,有林地面积28 134 hm2,森林覆盖率为88.40%。研究区属亚温带大陆性季风气候,四季分明,春季少雨干燥,夏季温热多雨,秋季凉爽多晴,冬季漫长而寒冷。年平均气温3.8 ℃,年降水量为700~800 mm,无霜期120~130 d。土壤类型以暗棕色森林土为主,平均厚度20~100 cm。
研究区植被属于长白山植物区系,林分类型有天然阔叶红松林、次生针阔叶混交林等[14]。主要乔木树种有:红松(Pinuskoraiensis)、色木槭(Acermono)、白牛槭(Acermandshuricum)、胡桃楸(Juglansmandshurica)、水曲柳(Fraxinusmandshurica)、蒙古栎(Quercusmongolica)、怀槐(Maackiaamurensis)、白桦(Betulaplatyphylla)、大青杨(Populusussuriensis)、裂叶榆(Ulmuslaciniata)、紫椴(Tiliaamurensis)等。灌木有:东北山梅花(Philadelphusschrenkii)、毛榛(Corylusmandshurica)、瘤枝卫矛(Euonymusverrucosus)、刺五加(Eleutherococcussenticosus)等。乔木分布于低山、丘陵地带,灌木多分布在草甸、沼泽、河谷地带。
2 研究方法
2.1 样地建设和群落调查
在研究区的老龄林内,采用TOPCON全站仪设置1块500 m×600 m固定监测样地。该样地平均海拔690 m,最大坡度为40°[15]。样地的林冠郁闭度大于0.80,林分密度1 694株hm2,胸高断面积31.45 m2·hm-2[16-17]。将样地划分为750个连续的20 m×20 m样方。为方便测量和记录,将每个20 m×20 m的样方又分成16个5 m×5 m的小样方。样地建设和群落调查参照CTFS(Center for Tropical Forest Science)的技术规范[18]。由于起测胸径直接影响对森林结构的合理划分、森林资源的准确计量以及对林分内部特征的正确把握[19],因此,本研究中对胸径大于1 cm的树木进行每木调查,以确定不同的起测胸径和取样尺度对林木分布格局的影响。调查内容包括树种、胸径、树高、冠幅(东西冠幅长×南北冠幅长)及枝下高,并挂牌标记。测量植株到5 m×5 m小样方4个顶点的距离,并换算为其在整个样地内的相对坐标。
2.2 点格局分析法
本研究应用Ripley’s K函数对不同种群、不同空间尺度及优势木进行空间点格局分析。Ripley’s K函数计算过程的实质就是选取不同尺度的半径,以此半径确定搜索范围得到一定数量的点数据,然后对获取的点数据,进行点格局分析。Ripley’s K函数可用如下公式来表示[20-21]:
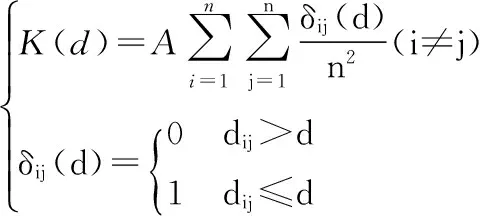
(1)
式中:A为样地面积;d为选择的尺度;dij为任意两个个体i与j之间的距离,n为样地内个体数。
结合Diggle提出的边缘校正和一项平方根转换,将L(d)定义为公式(2)[22]。L(d)将K(d)线性化,使其方差稳定并应用置信区间检验结果的真实性。当L(d)高于上包迹线时,数据点为聚集分布,在上下包迹线间为随机分布,低于下包迹线为均匀分布[3]。

(2)
本文在ArcGIS平台上采用Ripley’s K函数来分析不同空间尺度下的种群分布格局。将整个样地内的每株单木,看成平面中一个具有坐标的平面点,将测量得到的树木相对坐标换算成绝对坐标,将所有林木的坐标、树种、胸径、树高、冠幅以及枝下高作为点的属性。另外,在ArcGIS中应用Ripley’s K函数时还需要进行边缘校正[23-25]。本文选择的方法参数为最小包围矩形,选择里普利的边缘校正公式边缘校正法。分析时,取起算距离段数量为默认值10段,起算距离参数为5 m,距离增量设置为5 m,置信度为99.90%。选择老龄林中乔木种群数量前6位的树种进行10个不同尺度的空间聚集度研究,并分析不同起测胸径对空间格局变化的影响。另外,本文还对样地内的优势木的空间聚集度进行分析。根据优势木的定义,选取每公顷胸径最大的前100株树木作为优势木。由于样地面积为30 hm2,因此,一共选取3 000株优势木进行空间聚集度分析。
2.3 空间自相关分析
空间自相关是指一些变量在同一个分布区内的观测数据之间潜在的相互依赖性[26]。任何相邻的地理要素无论是竞争关系还是共存关系,都要受到生存环境的影响而存在一定的关系,而这种必然存在的关系被称为空间依赖。本文采用空间自相关方法对研究区域内的种群进行空间自相关性分析。用5个参数对空间格局进行评估,即Moran’s I指数、预期指数、方差、Z得分和P值。当Moran’s I>0时,表示种群间显示出空间正相关,而且计算结果越大,代表相关性越强,种群之间存在较强的相互利用关系;当Moran’s I<0时,表示种群间为空间负相关,数值越小代表负相关性越强,说明种群之间在竞争过程中的相互排斥作用越显著;当Moran’s I=0时,表示没有相关关系,即空间格局分布是随机的[27]。由于本文研究的是一个大区域中的众多点数据,因此,在参数选择上选取固定距离法进行计算,对优势木不同距离组和优势木种群间分别进行空间自相关分析。
3 结果与分析
3.1 群落树种组成分析
由表1可知,通过对样地的群落调查,发现老龄林的种群组成十分丰富,共监测到52种木本植物。物种数最多的科是槭树科,包括6个种,其次是蔷薇科、桦木科和忍冬科。样地内个体数量排在前20位的种群,其中乔木类16种,灌木类4种。老龄林中簇毛槭(Acerbarbinerve)、千金榆(Carpinuscordata)、白牛槭、色木槭、裂叶榆和花楷槭(Acerukurunduense)6个种群的个体数量相对较多,6个种群的总株数占整体的54.30%。先锋演替树种裂叶榆、色木槭和紫椴个体数量较多。根据优势木定义,老龄林样地中优势木共包含17个种群,其中:乔木为簇毛槭、千金榆、白牛槭、色木槭、裂叶榆、花楷槭、紫椴、青楷槭、红松、水榆花楸、黄檗、胡桃楸、水曲柳、沙松、稠李、枫桦;灌木为暴马丁香。老龄林优势树种主要由乔木组成,优势木的平均胸径普遍较大(>35 cm),平均胸径在40 cm以下的占10.50%,平均胸径40~50 cm占65.43%,平均胸径大于50 cm的占23.97%。

表1 老龄林样地内物种分布情况
根据对群落中所有样本的胸径调查情况,将所有的林木分别按照不同的起测胸径(D)(1、2、3、4、5、6、7、8 cm)进行统计,起测胸径为2 cm时,林木株数占整个调查样本的92.09%(46 800株);起测胸径为3 cm时,林木株数占整个调查样本的71.92%(36 547株);起测胸径为4 cm时,林木株数占整个调查样本的58.36%(29 657株);起测胸径为5 cm时,林木株数占整个调查样本的48.74%(24 767株);起测胸径为6 cm时,林木株数占整个调查样本的42.28%(21 486株);起测胸径为7 cm时,林木株数占整个调查样本的37.29%(18 949株);起测胸径为8 cm时,林木株数占整个调查样本的33.18%(16 863株)。
3.2 空间分布格局分析
3.2.1 不同起测胸径林木的空间分布格局
不同起测胸径林木的空间分布情况如图1所示。
当起测胸径为1 cm时,在10种不同的空间尺度(d)下,林木呈现出显著的空间聚集性,各单株木之间具有一定的相关性,随着空间尺度的增加,空间聚集性程度不断增加(图1(a))。
当D≥2 cm时,空间分布格局(图1(b))与D≥1 cm时很相似,说明无论采用1 cm还是2 cm作为起测胸径,对研究结果几乎没有影响。
当D≥3 cm时,种群也呈现出空间聚集性,但是从小尺度到大尺度的变化过程中,空间聚集性程度逐渐变低(图1(c))。
当D≥4 cm时,在不同尺度下均呈现出了由空间随机性转变为空间聚集性;空间尺度d<15 m时,空间聚集性程度逐渐增加,15 m≤d≤50 m时,空间聚集性先变大后变小,d>50 m时,空间聚集性有逐渐递减的趋势(图1(d))。
当D≥4 cm时,d<40 m时,呈现出空间聚集性,5 m≤d≤25 m及d>40 m时,呈随机性(图1(e))。
当D≥5 cm时,d<30 m呈现空间随机性,30 m≤d≤40 m呈现空间聚集性,空间聚集性成都很低(图1(f))。
当D≥7 cm时,整体呈现空间随机性,30 m≤d≤45 m时,接近空间聚集性(图1(g))。
当D≥8 cm时,整体呈现明显的空间随机性(图1(h))。
综上所述,不同的起测胸径对林木的空间格局分布有重要的影响。如果起测胸径选择过小,计算过程就会变得繁杂;反之,起测胸径选择过大,又不能代表小胸径林木的空间分布情况。考虑到我国现行一、二类调查技术,将乔木的起测胸径确定为5 cm,因此,本文选择以5 cm为起测胸径进行种群格局分析。
3.2.2 不同种群空间分布格局分析
研究区内个体数量最多的6个种群(D≥1 cm)在样地中的空间分布情况如图2所示。可以看出,簇毛槭、白牛槭、色木槭和裂叶榆在整个样地中分布得比较密集;千金榆、花楷槭分布不均匀,千金榆在样地东北部分布比较稀疏,其它部分相对密集;花楷槭在整个样地的南部分布比较稀疏,在北部分布比较密集。总体来看,6个种群的点分布是相对密集的。
如图3所示,由于本文的统计分析是基于起测胸径5 cm,因此,对不同的空间尺度下(5、10、15、20、25、30、35、40、45、50 m)的林木空间聚集度进行分析。簇毛槭在5~35 m空间尺度上呈现出了明显的空间随机性,但是从小尺度到大尺度,均匀程度上有所改变,呈现出随机性逐渐变大的趋势,d>40 m呈现出空间离散性分布(图3(a));白牛槭无论在任何尺度下其空间分布均呈现出空间随机性(图3(b));千金榆在任何尺度下表现出空间随机性,且随机性由小逐渐变大后再逐渐变小,且在更大的空间尺度下保持了显著的空间随机性(图3(c));色木槭在5~35 m呈现出空间随机性,从小尺度到大尺度空间随机性程度逐渐变小,类似于白牛槭,但是空间聚集的程度远小于白牛槭,d>35m呈现空间离散性(图3(d))。裂叶榆在d>20m尺度下表现出的空间离散性,5~20 m尺度下表现出空间随机性(图3(e))。花楷槭在任何尺度下均表现出空间离散性(图3(f))。综上可知,白牛槭是6个种群中聚集性最高的。
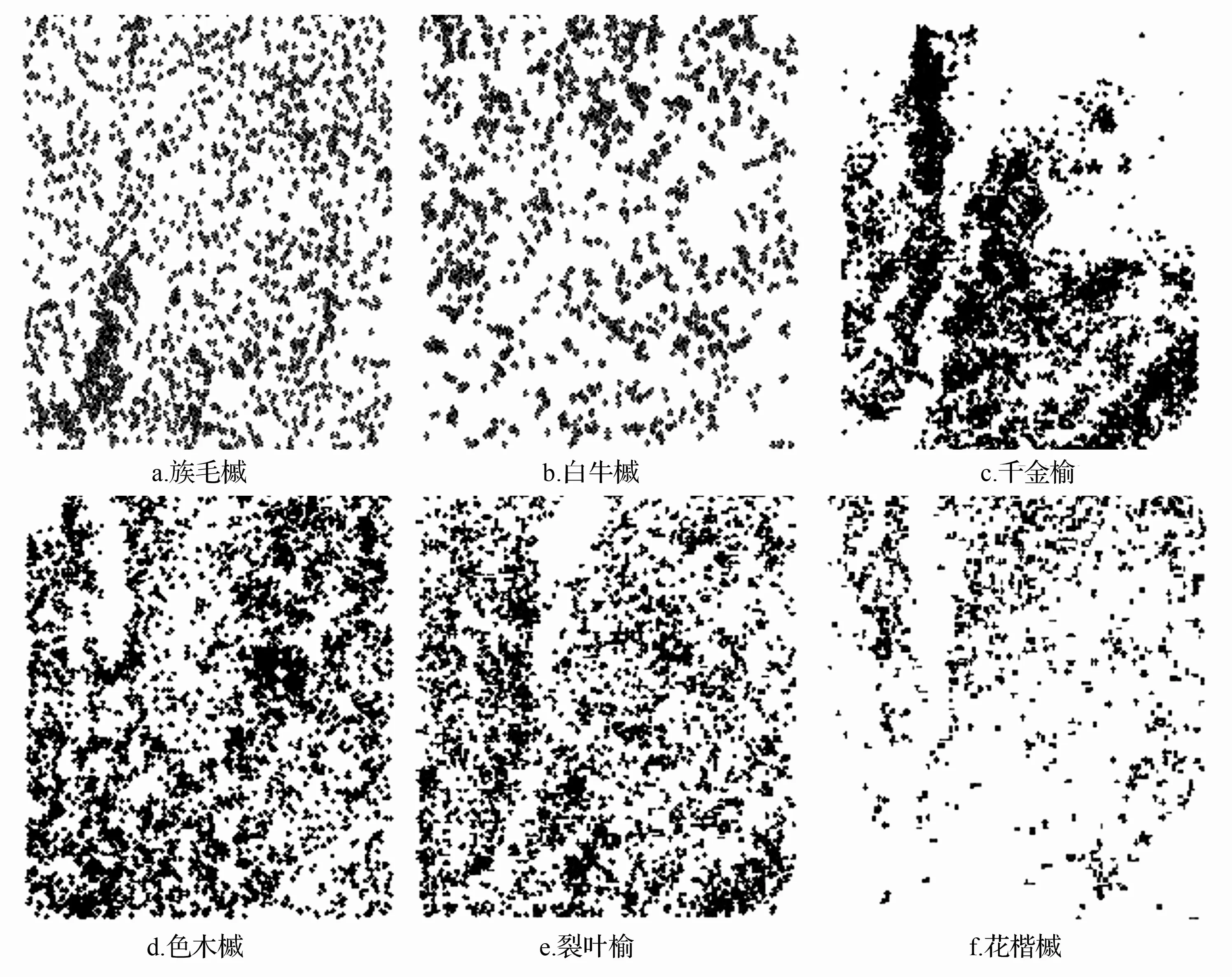
图2 6个种群点密度

图3 6个种群的空间聚集性
3.2.3 优势木的空间分布格局
在不同的空间尺度下,优势木的空间聚集度表现为:优势木在d<25 m时呈现空间聚集性,且随着空间尺度的增大,空间聚集性程度逐渐降低;在d=30 m附近呈现空间随机性分布,随着空间尺度的增加,逐渐向空间离散分布过渡。结果证明,优势木在空间尺度较小时,生长的比较密集,在空间尺度较大时,则呈现出离散的分布特点。
3.3 种群的空间自相关性
3.3.1 优势树种群间的空间自相关性
选取优势木中前4个乔木类种群,进行两两种群间的空间自相关性分析,结果如表2所示。由表2可知,簇毛槭与千金榆的相关性相对较大,与白牛槭、色木槭的空间相关性程度相似,且置信度相对大于簇毛槭与千金榆的置信度;千金榆与簇毛槭的相关性最大,其次是白牛槭和色木槭,且色木槭与千金榆空间相关性的置信度相对较大;白牛槭与色木槭的空间相关性大于白牛槭与簇毛槭、千金榆的空间相关性,且白牛槭与色木槭空间相关性的置信度也是最高的;色木槭与白牛槭的空间相关性程度也大于其它两个种群,而且置信度也较大。综上所述,4个种群相互之间都存在一定的空间相关性,相关性程度都接近0.50,所以种群之间除了竞争关系,还存在相互协调生长的关系。
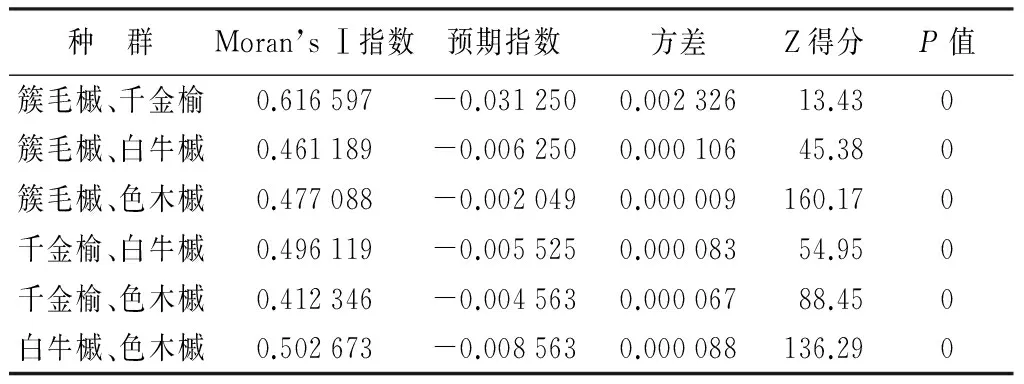
表2 优势木种群间空间自相关性
3.3.2 优势木不同空间距离空间自相关性
对优势木进行不同空间距离(50、100、150、200、250、300、350、400、450、500 m)的空间相关性研究,结果如表3所示。由表3可知,优势木在空间距离较小时空间相关性很高。例如,当空间距离为50 m时,Moran’s I指数为0.937 6,显示了较高的空间相关性;随着空间距离尺度的增大,优势木之间正相关性变弱;当研究尺度大于样方边长距离一半(250 m)时,空间相关性程度小于0.50;当空间尺度增加到200 m时,方差为0,说明结果波动范围很小,甚至不存在波动,结果落在置信区间的概率达到最大值,具有很高的置信度,这也说明本研究中的空间相关性结果具有较高的可信度;优势树种之间虽然属于不同种群,存在一定的竞争关系,但是也存在相互利用的关系;优势木的空间自相关系数为正数,即在所有尺度下均呈正关联,只是在某些尺度下关联性比较显著,而在另一些尺度下关联性不显著。

表3 优势木不同空间距离空间自相关性
综上所述,研究的空间尺度的大小,对优势木的空间相关性研究结果有很大的影响,种群间的相关性除了受到环境因素与种群类别因素的影响外,树木之间的距离也是至关重要的因素。
4 结论与讨论
林分的空间格局是森林植被的重要特征,它不仅对于植被的生长、发育以及系统的稳定性具有重要的意义,而且与这些植被相互作用的其它有机体(如食草动物、传粉者等)也有重要的影响。本文以吉林省蛟河市林业实验区管理局30 hm2老龄林为研究对象,分析老龄林的林木空间分布格局和种群空间自相关性,得出结论:研究样地内个体数量最多的前6个种群分别是簇毛槭、千金榆、白牛槭、色木槭、裂叶榆和花楷槭,总株数占整体的54.30%。先锋演替树种裂叶榆、色木槭和紫椴的胸高断面积较大,且三者个体数量较多。簇毛槭和千金榆是研究区内最主要的树种。色木槭相对于簇毛槭和千金榆竞争力较弱,它的斑块只出现在簇毛槭和千金榆的斑块间隙,因此,它与簇毛槭和千金榆的关系也十分密切,不同的起测胸径对空间聚集度有一定的影响。在实际工作中,起测胸径是以平均胸径的0.40倍来确定的,然而由于具体林分的条件差异性,不同研究中的起测胸径往往差异较大,很难直接将结果进行比较[16]。因此,研究空间格局分布时,起测胸径是首先需要考虑的因素。基于起测胸径5 cm对不同的空间尺度(5、10、15、20,、25、30、35、40、45、50 m)的林木空间聚集度进行分析,发现6个种群均呈现出空间聚集性,证明了该林场的老龄林的生长密度较高。6个种群中簇毛槭是种群数量最多的,但是却不是空间聚集度最高的,证明种群的空间分布情况不单单取决于种群的数量,还取决于种群之间的相关性、竞争能力、环境因素等;对优势木的空间相关性研究表明,无论是对优势木进行种群间的相关性研究,或优势木不同空间距离尺度研究,优势木之间具有正向的空间自相关性,只是相关性程度有所差异,优势木的相关性受到空间尺度的影响较大。
本文采取大样方数据对吉林省蛟河市林业实验区管理局的老龄林进行研究时,只分析了正方形样方的各林分空间格局参数,但是没有考虑不同的样方形状、地形、坡位、坡向等其他因素及其交互作用对点格局分析性能的影响,未来还需更进一步深入探讨。
[1] 王芳.基于ArcGIS林分空间结构的研究:以武夷山保护区为例[D].南京:南京林业大学,2013.
[2] 孔雷,杨华,亢新刚,等.林木空间分布格局研究方法综述[J].西北农林科技大学学报(自然科学版),2011,39(5):119-125.
[3] 杨洪晓,张金屯,吴波,等.毛乌素沙地油蒿种群点格局分析[J].植物生态学报,2006,30(4):563-570.
[4] 张金屯.植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349.
[5] 惠刚盈.森林空间结构量化分析方法[M].北京:中国科学技术出版社,2003.
[6] 惠刚盈,李丽,赵中华,等.林木空间分布格局分析方法[J].生态学报,2007,27(11):4717-4728.
[7] 秦丰林,杨丽.基于Ripley’s K函数的昆山市景观格局时空变化特征研究[J].中国资源综合利用,2014,32(11):51-56.
[8] 贾国栋,余新晓,邓文平,等.北京山区典型流域不同海拔椴树种群的空间点格局分析[J].生态环境学报,2011,20(6-7):996-1002.
[9] 岳永杰,余新晓,武军,等.北京山区天然次生林种群空间分布的点格局分析:以雾灵山自然保护区为例[J].中国水土保持科学,2008,6(3):59-64.
[10] HAASE P. Spatial pattern analysis in ecology based on Ripley’s K-function: Introduction and methods of edge correction[J]. Journal of Vegetation Science,1995,6(4):575-582.DOI:10.2307/3236356.
[11] DRUCKENBROD D L, SHUGART H H, DAVIES I. Spatial pattern and process in forest stands within the Virginia piedmont[J]. Journal of Vegetation Science,2009,16(1):37-48.
[12] 王国严,方江平,许新勇,等.雅鲁藏布江中游干旱沙地砂生槐种群结构与点格局分析[J].中国沙漠,2010,30(5):1092-1098.
[13] 尤海舟,贾成,樊华,等.格局分析的最新方法-点格局分析[J].四川林业科技,2009,30(6):198-235.
[14] 吴金卓,孔琳琳,王娇娇,等.吉林蛟河不同演替阶段针阔混交林凋落物持水特性研究[J].南京林业大学学报(自然科学版),2016(2):113-120.
[15] 丁胜建,张春雨,夏富才,等.老龄阔叶红松林下层木空间分布的生境关联分析[J].生态学报,2012,32(11):3334-3342.
[16] 彭萱亦.不同演替阶段针阔混交林生物多样性评价指标体系研究[D].哈尔滨:东北林业大学,2014.
[17] 吴金卓,蔡小溪,林文树,等.吉林蛟河不同演替阶段针阔混交林木本植物α多样性评价[J].林业科技开发,2015,29(3):128-133.DOI:10.13360/j.issn.1000-8101.2015.03.031.
[18] 吴金卓,蔡小溪,林文树.吉林蛟河不同演替阶段针阔混交林土壤健康评价[J].东北林业大学学报,2015,43(6):78-82.
[19] 牛丽丽,余新晓,刘彦,等.不同起测胸径对判定油松分布格局的影响[J].北京林业大学学报,2008,30(S2):12-16.
[20] 刘妍妍,金光泽.小兴安岭阔叶红松林粗木质残体空间分布的点格局分析[J].生态学报,2010,30(22):6072-6081.
[21] 郭垚鑫,康冰,李刚,等.小陇山红桦次生林物种组成与立木的点格局分析[J].应用生态学报,2011,22(10):2574-2580.
[22] 杨云方,丁晖,徐海根,等.武夷山典型常绿阔叶林4个主要植物种群点格局分析[J].生态与农村环境学报,2013,29(2):184-190.
[23] 汤孟平,唐守正,雷相东,等.Ripley’s K(d)函数分析种群空间分布格局的边缘校正[J].生态学报,2003,23(8):1533-1538.
[24] 欧阳芳,戈峰,袁哲明,等.点格局分析函数的边缘校正及其在昆虫种群格局分析中的应用[J].生态学报,2009,29(4):1804-1813.
[25] 时培建,戈峰,杨清培,等.点格局分析中边缘校正的一种新算法及其应用[J].生态学报,2009,29(2):804-809.
[26] 朱子明,祁新华.基于Moran’I的闽南三角洲空间发展研究[J].经济地理,2009,29(12):1977-1980.
[27] 王芳,夏丽华,张太煜.基于GIS的珠江三角洲城市群结构与空间关联研究[J].广州大学学报(自然科学版),2010,9(1):47-53.
1)中央高校基本科研业务费专项资金项目(2572015CA01);“十二五”国家科技支撑计划专题(2012BAC01B03-3);国家自然科学基金项目(31400539)。
许文秀,女,1990年10月生,东北林业大学工程技术学院,硕士研究生。E-mail:511741156@qq.com。
吴金卓,东北林业大学工程技术学院,副教授。E-mail:wujinzhuo1980@163.com。
2016年12月13日。
S714
责任编辑:王广建。
Spatial Point Pattern of An Old Growth Forest//Xu Wenxiu, Lin Wenshu, Wu Jinzhuo(Northeast Forestry University, Harbin 150040, P. R. China)//Journal of Northeast Forestry University,2017,45(3):17-22,28.
TOPCON total station was used in an old growth forest in Jilin Province Jiaohe Forestry Administration of Experimental Area to set up a 500 m×600 m fixed monitoring plot and community survey was conducted. The Ripley’s K function on ArcGIS platform was used to study the spatial aggregation of tree species in different scales in the sample plot. The spatial autocorrelation method was applied to study the spatial dependence of different species in the study plot. The top six tree species in terms of population size wereAcerbarbinerve,A.mandshuricum,Carpinuscordata,A.mono,Ulmuslaciniata, andA.ukurunduense. The minimum measured diameter had important impact on the spatial pattern of stands. When the minimum measured diameter was 5 cm, the top six largest populations all showed spatial clustering in varying degrees and the spatial aggregation ofA.mandshuricumwas the largest among them. The dominant tree species in the old growth forest were determined and the spatial aggregation of dominant trees was analyzed. The dominant trees showed dense distribution in smaller spatial scales and showed scattered distribution in larger spatial scales. Different minimum measured diameter also had certain impact on the study results, therefore, this factor should be considered firstly while studying the spatial pattern of individual trees. Among different tree species and under different spatial scales, the dominant tree species all showed positive spatial autocorrelation with varying correlation degrees, and the spatial autocorrelation was greatly affected by the spatial scales.
