南京城郊毛竹林内外气温及土壤温度特征1)
2017-08-07杨丹杨静张金池庄家尧薛雪成举荣
杨丹 杨静 张金池 庄家尧 薛雪 成举荣
(南京林业大学,南京,210037) (科学出版社) (南京林业大学)
南京城郊毛竹林内外气温及土壤温度特征1)
杨丹 杨静 张金池 庄家尧 薛雪 成举荣
(南京林业大学,南京,210037) (科学出版社) (南京林业大学)
为揭示毛竹(Phyllostachysedulis)林调节林内气温及土壤温度的作用。使用Decagon微气象监测系统对南京城郊毛竹林内太阳辐射(RAR)、气温(Tr)及0~40 cm土层土壤温度(Ts)等因子进行长期监测,以裸地的试验观测作为对照,分析了南京城郊毛竹林内Tr及Ts变化特征规律。结果表明:林内RAR呈多峰曲线变化,毛竹林能有效削减林内RAR,夏秋季尤其明显;裸地与林内Tr变化趋势一致,均呈单峰曲线变化,毛竹林具有在夜间提高林内Tr在白天降低林内Tr的作用,春夏秋冬季毛竹林均能降低林内月均Tr(约0.5 ℃);浅层Ts日变化呈正弦函数图像,并随着土壤深度的增加,Ts日变化幅度越小,春夏秋季,同一土层,裸地Ts高于林内Ts,并随土壤深度的增加,两者差异越不显著,Ts季节变化曲线与林内Tr季节变化曲线一致,毛竹林在夏季具有降低Ts(约为1 ℃)冬季具有升高Ts的作用(2~4 ℃);毛竹林可以滞后Ts最值出现时刻30~90 min;裸地与林内RAR、Tr及各土层Ts均呈极显著的相关性(P<0.01)。毛竹林在降低林内Tr、缓和林内Ts变化和调节Ts方面具有极显著作用。
林内气温;土壤温度;太阳辐射;毛竹林
Forest air temperature; Soil temperature; Solar radiation;Phyllostachysedulisforest
温度是植被生长发育的主要环境条件,直接影响森林土壤微生物活性、枯落物分解率及土壤呼吸[1-4]。森林温度效应的研究对森林生态系统的功能研究具有重要的意义[5]。伴随着气候变暖,森林温度上升,将不利于森林植被的生长发育。Janík[6]的研究表明,森林土壤温度能直接影响植物的生长;Luxhøi et al.[7]认为,土壤温度对土壤微生物活性有直接影响;王俊峰[8]也认为,气温温度上升对生态系统平衡有重要影响。而毛竹(Phyllostachysedulis)具有生长快、成材早、经济价值高的特点,且具有较强的固碳能力,对气候变化具有不可低估的影响[9-11]。
目前,相关学者对土壤温度及林内外气温变化特征研究取得了一系列的研究成果,如Brian et al.[12]通过对澳大利亚109个气象站的气温、降雨及土壤温度数据处理分析,建立了澳大利亚土壤温度预测模型;孙金伟等[13]通过对长白山红松(Pinuskoraiensis)林及空旷地空气温湿度及土壤温度的长期对比研究,探讨了森林调节林内小气候的效应;陈继康等[14]通过对冬小麦生育期土壤温度及气温的长期监测,研究了华北平原不同耕作方式麦田土壤温度及其气温的响应特征;侯建花等[15]通过分析安吉县土壤温度及气温的变化特征建立了土壤温度预测模型,并探索了土壤温度的滞后效应。国内外对土壤温度及气温的研究大多分为2种:一为利用气温进行土壤温度的预测;二为探讨森林调节林内小气候的效应。在森林调节林内小气候效应方面大多探讨的也是森林调节林内气温及土壤温度的作用,在森林滞后林内气温及土壤温度方面的研究还较少。
笔者根据南京市铜山土壤温度及气象系统资料,以日、季节、年等不同尺度,对裸地与毛竹林内各土层(0、10、15、40 cm)土壤温度、林内气温及林内太阳辐射进行统计分析与比较,探讨了毛竹林调节林内外气温及土壤温度的作用,并研究了毛竹林对林内气温及土壤温度滞后效应,旨在揭示南京城郊毛竹林小气候效应,加强对森林生态系统功能的认识,为林内生态环境保护、森林经营、保护提供科学依据。
1 研究地区概况
试验区位于长江三角区西部南京市国营东善桥林场铜山分场(31°35′~31°39′N,118°50′~118°52′E),属北亚热带季风气候区,年平均气温15.1 ℃,无霜期229 d,年日照时间199 h,区内气候温和湿润,四季分明,水热资源比较丰富,生长季长。年平均降水量1 100 mm,地形为苏南丘陵,土壤类型为黄棕壤。试验地点选择不受林缘和林窗影响的毛竹林地与裸地,毛竹林郁闭度0.8以上,毛竹林地与裸地坡向均为东北向,立地条件一致,土壤厚度为60 cm左右,60 cm以下为砂岩风化母质层,海拔26.0 m,地下水位位于10.0 m以下。
2 研究方法
在土壤与光照条件一致的毛竹林与裸地内各设2个试验点。试验点设在数株毛竹组成的多边形的中心,附近10 cm内无直径0.5 cm以上树根分布,并保证2试验点在同一水平方向上。在试验点地表覆盖2 cm厚的毛竹凋落物,设置Decagon微气象监测系统,于2012年6月至2013年5月连续监测毛竹林内与裸地气温(Tr)、太阳辐射(RAR)及不同深度(0、10、15、40 cm)的土壤温度(Ts)。每30 min记录一次数据,取每个样地的2组气象数据的均值进行数据统计与分析。四季的划分采用气候学统计法以公历3至5月为春季,6至8月为夏季,9至11月为秋季,12至次年2月为冬季。取3月12日、6月14日、9月15日、12月15日(通过查南京历史天气,均为晴天)数据分别代表春、夏、秋、冬各季RAR、Tr及Ts的日数据;取2012至6月至2013年5月每月瞬时数据的平均值作为该月的温度值,进行RAR、Tr及Ts季节变化分析;并对林内外Tr与各土层Ts每日出现最值时刻的频率进行年统计分析。
利用Microsoft Excel 2007软件进行数据处理,Origin 8.5绘图软件进行数据分析及绘图,采用SPSS 19.0进行相关的统计与分析,采用配对样本T检验的方法比较裸地与毛竹林不同数据组间的差异性,显著性水平设定α=0.05,用Pearson相关系数评价不同因子间的相关性。
3 结果与分析
3.1 日变化
3.1.1 太阳辐射
由图1可见,裸地RAR变化趋势为单峰变化趋势,林内RAR呈多峰变化趋势,裸地RAR变化幅度较毛竹林大。裸地与毛竹林内RAR出现、上升与消失的时间一致,裸地日太阳辐射始终高于林内,夏秋季,两者差异显著(P<0.05),冬春季差异不显著(P>0.05)。可知,毛竹具有显著削减林内RAR的作用。经分析,可能是由于毛竹林冠层对RAR的反射、过滤等原因,使林内RAR始终小于裸地。
3.1.2 空气温度
由图2所示,Tr呈单峰曲线变化,与RAR日变化趋势一致。00:00至08:00林内Tr略高于裸地,08:00至16:00裸地Tr高于林内,16:00至24:00林内Tr略高于裸地。除春季外,林内Tr最高温总低于裸地,最低温总高于裸地。表明,毛竹林在夜间具有一定保温能力,在白天具有降温能力。Tr主要受RAR的影响:白天,毛竹林冠层削减了林内RAR,林内RAR小于裸地,林内升温比裸地慢,使毛竹林内Tr小于裸地;夜晚由于毛竹林冠层的遮挡作用,减少气流流动,使林内热量不易扩散,降温缓慢,从而比裸地Tr高。
3.1.3 土壤温度
由图3所示,浅层(0~15 cm)Ts呈正弦函数曲线变化,并随着土壤深度的增加,Ts日变化幅度越小。Ts主要受RAR及大气Tr的影响,00:00至08:00,RAR未出现,Tr逐渐降低,使Ts也随之降低,08:00过后,RAR开始缓慢上升,Tr逐渐升高,使Ts也随之升高,并在RAR下降后下降。随着土壤深度的增加,Ts受RAR及Tr的影响变小,导致Ts变化幅度变小[16]。春秋季时,00:00至08:00,林内表层Ts高于裸地,08:00过后,裸地表层Ts逐渐高于林内,并在12:00至14:00两者差异最显著;冬季,毛竹林与裸地表层Ts差异不显著(P>0.05),这与Tr有关,冬季温度过低,毛竹林与裸地表层Ts差异不显著。表明毛竹林在夜间对土壤具有一定保温能力。

图1 不同季节毛竹林与裸地RAR日变化

图2 不同季节毛竹林与裸地气温日变化
3.2 季节变化
3.2.1 太阳辐射
由表1所示,裸地月均RAR显著大于林内。9月裸地月均RAR与林内差异最大,为88.76 W·m-2,5月裸地月均RAR与林内差异最小,为15.57 W·m-2,林内RAR较裸地年均降低53.68 W·m-2。表明毛竹林能显著削减林内RAR,夏季尤其显著。可能是由于夏季为树木的生长旺季,郁闭度和叶面积指数等均较其他季节更高[17],对RAR的影响更为强烈,削减RAR能力加大。
3.2.2 空气温度
由表1所示,裸地与林内Tr季节变化均呈单峰变化,春季Tr缓慢上升,于7月份达到最大值,秋季温度缓慢下降,于1月份达到最小值。裸地Tr始终高于林内,每月均高0.5 ℃左右。表明,毛竹林能降低林内月均Tr(约0.5 ℃)。可能是由于林冠层削弱了林内的RAR、降低了地面长波辐射,并且使进入冠层的平流、乱流涡旋体受到枝叶的阻截和磨擦作用,使空气热量交换强度被削弱,从而导致林内Tr比裸地低[18]。
3.2.3 土壤温度
由表1所示,各层Ts季节变化趋势为单峰型,与林内Tr季节变化趋势一致,Ts在7、8月份达到峰值,在12、1月达到温度最小值,0 cm土层Ts最大值与最小值差异最大,并随土壤深度的增加,土层Ts最大值与最小值差异逐渐减小,这是由于表层土壤受RAR及Tr影响较大,深层土壤影响较小,表明随着土壤深度的增加,Ts变化幅度越小;1—2月,同一土层Ts,林内高于裸地,3月至11月,林内低于裸地,表明在冬季,毛竹林能提高月均Ts 1 ℃左右,在春、夏、秋季,毛竹林能降低Ts 2~4 ℃。分析认为,这是由于冬季毛竹林地具有大量凋落物,对土壤有一定遮挡作用,缓解了Ts的扩散,从而有一定保温能力,且毛竹林内土壤微生物含量较多,土壤呼吸释放大量热能。
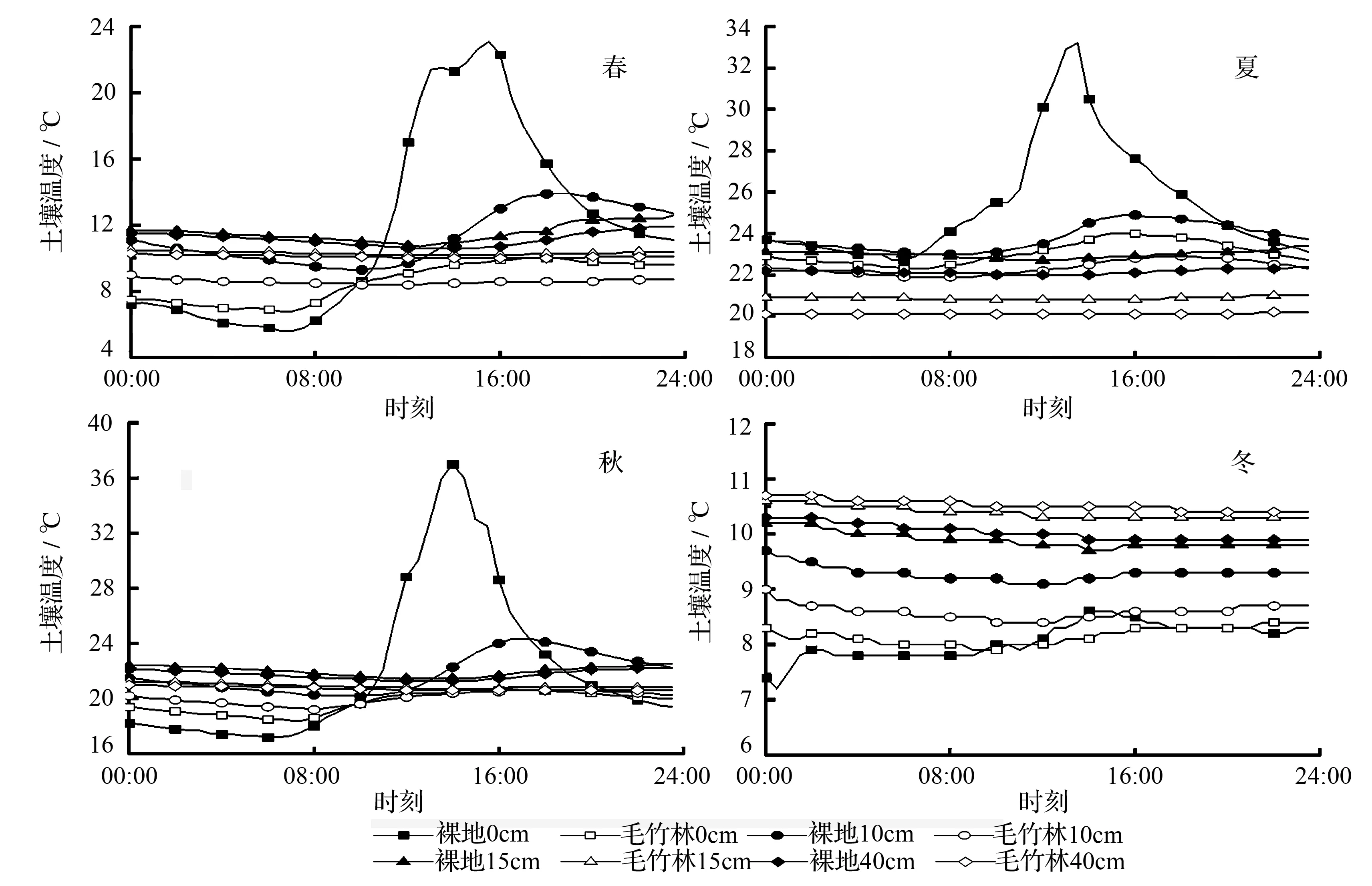
图3 不同季节毛竹林与裸地土壤温度日变化
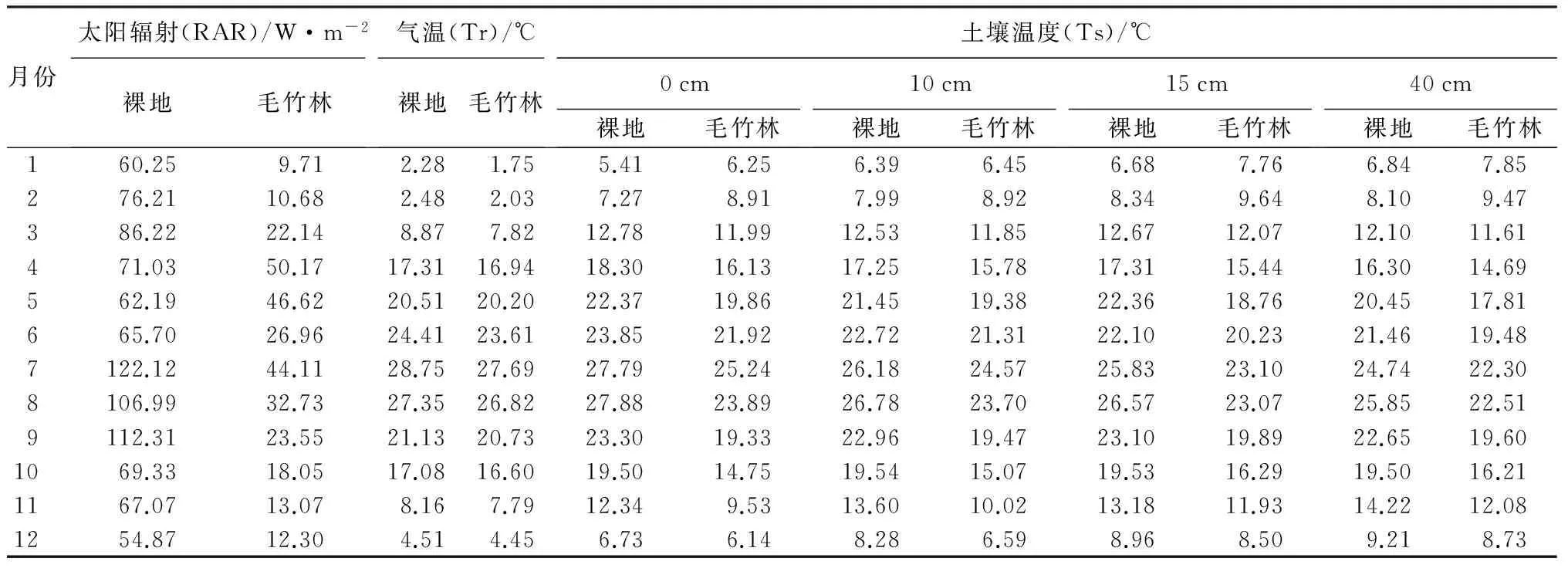
月份太阳辐射(RAR)/W·m-2裸地毛竹林气温(Tr)/℃裸地毛竹林土壤温度(Ts)/℃0cm裸地毛竹林10cm裸地毛竹林15cm裸地毛竹林40cm裸地毛竹林160.259.712.281.755.416.256.396.456.687.766.847.85276.2110.682.482.037.278.917.998.928.349.648.109.47386.2222.148.877.8212.7811.9912.5311.8512.6712.0712.1011.61471.0350.1717.3116.9418.3016.1317.2515.7817.3115.4416.3014.69562.1946.6220.5120.2022.3719.8621.4519.3822.3618.7620.4517.81665.7026.9624.4123.6123.8521.9222.7221.3122.1020.2321.4619.487122.1244.1128.7527.6927.7925.2426.1824.5725.8323.1024.7422.308106.9932.7327.3526.8227.8823.8926.7823.7026.5723.0725.8522.519112.3123.5521.1320.7323.3019.3322.9619.4723.1019.8922.6519.601069.3318.0517.0816.6019.5014.7519.5415.0719.5316.2919.5016.211167.0713.078.167.7912.349.5313.6010.0213.1811.9314.2212.081254.8712.304.514.456.736.148.286.598.968.509.218.73
3.3 年统计特征
3.3.1 滞后分析
将裸地与林内Tr及各土层Ts每日出现最大值与最小值的时刻进行频率统计,结果见图4,裸地与毛竹林Tr出现最大值的时刻均为13:30,出现最小值的时刻均为23:30,且裸地与毛竹林出现最值时刻的频率分布差异不大;毛竹林各土层Ts出现极值时刻普遍比裸地晚30~90 min,且频率分布较裸地分散。表明,毛竹林具有缓温的能力,可以滞后Ts出现最值的时间,滞后30~90 min。
3.3.2 极值分析
对裸地与毛竹林月Ts最高温与最低温之差进行统计分析(表2),裸地与林内Tr最高温差值均为正数,春夏秋季,裸地与林内Tr最低温差值为正数,冬季为负数,表明,毛竹林全年具有降低Tr极大值的能力,且冬季具有升高Tr极小值的能力。表层土壤最高温差值均为正数,除2月,10、15、40 cm土层土壤最高温差值为负数之外,其他时间最高温差值均为正数;各层土壤Ts最低温差值变化趋势一致,均为8至10月最大,1至4月为负值。表明毛竹林能降低Ts最高值,这种降温作用在夏季尤其明显。其原因可能是因为夏季毛竹林枝叶繁茂,削减RAR能力强,使得林内Ts较裸地低。

图4 裸地与毛竹林气温与土温极值出现时刻频率分布

月份气温(Tr)/℃最高温最低温土壤温度(Ts)/℃0cm最高温最低温10cm最高温最低温15cm最高温最低温40cm最高温最低温115.7-5.49.3-4.80.50.50.5-1.5-0.5-1.128.9-3.14.4-3.1-0.7-0.4-0.6-2.0-0.1-1.439.40.14.90.52.11.01.0-0.61.20410.71.95.51.02.51.42.10.41.52.7510.24.05.62.63.52.72.02.42.73.3620.10.510.4-0.10.91.01.10.72.01.4726.20.718.1-0.22.40.01.30.92.71.8823.14.816.93.53.34.82.33.92.94.2920.12.114.51.22.84.32.22.82.63.51026.96.122.54.34.17.72.85.42.05.51110.44.84.13.61.55.70.23.60.83.9128.3-3.13.9-1.31.01.20.7-0.80-0.4
3.4 相关性分析
对毛竹林与裸地RAR、Tr及各土层Ts进行相关性分析,结果见表3。由表3可见,各因子间相关系数均较高,均呈极显著相关,林内Tr与裸地Tr相关性最强,相关系数达到1。各土层Ts之间相关系数均在0.950以上,呈极显著相关。表明Ts与RAR及林内Tr密切相关,相互影响。
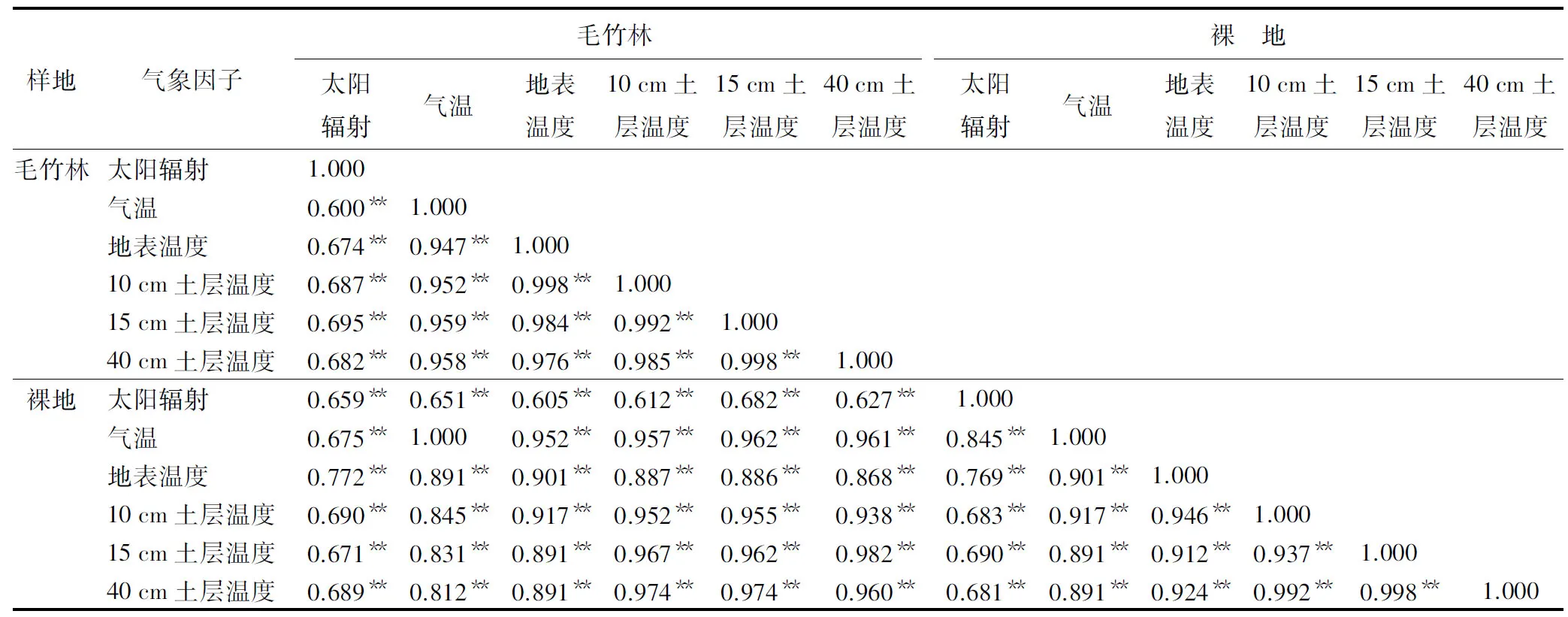
表3 毛竹林与裸地气象因子相关性
注:** 表示在0.01水平(双侧)上显著相关。
4 结论与讨论
4.1 裸地与毛竹林内太阳辐射变化特征
裸地RAR日变化呈单峰曲线,林内RAR为多峰曲线变化,这与徐云蕾等[19]研究结果一致,经分析,可能是因为林内RAR受林窗影响,林冠不断透光,加上太阳高度角的不断变化,使林内RAR呈多峰变化趋势。裸地RAR显著高于林内,夏秋季,林内RAR与裸地RAR差异最为显著[19],表明毛竹林对RAR有削减作用,夏秋季作用尤其明显。林内RAR受多重因素的影响,RAR通过冠层到林内,毛竹林冠层结构、叶的疏密程度、叶的吸收和反射特性都将影响RAR的变化特征,这还需进一步研究。
4.2 裸地与毛竹林内气温变化特征
Tr日变化趋势为单峰变化。白天,裸地Tr高于林内,且在11:00至13:00差异最为显著;夜晚,林内Tr略高于裸地,这与张一平等[20]研究结果一致,表明,毛竹林在夜间具有一定保温能力,在白天具有降温能力。除春季外,林内Tr最大值总低于裸地,最小值高于裸地。裸地与林内Tr季节变化趋势一致,呈单峰型变化趋势,这与马鸿儒等[21]研究结果一致。林内Tr始终低于裸地[22]。本研究结果表明,毛竹能降低林内月均Tr 0.5 ℃。
4.3 裸地与毛竹林内土壤温度变化特征
浅层Ts日变化呈正弦函数图像,并随着土壤深度的增加,Ts日变化幅度越小,这与李沙等[16]研究结果一致。除冬季外,同一土层,裸地Ts均高于林内,并随土壤深度的增加,两者差异越不显著,因为毛竹林冠层对RAR的阻挡作用,使林内Ts高于裸地,且随土壤深度的增加,受RAR及大气Tr的影响较小,使裸地与林内Ts差异变小。
Ts季节变化趋势为单峰型变化趋势,与林内Tr季节变化趋势一致[23];森林具有夏天降低Ts,冬天升高Ts的作用[18]。本研究结果为,除1至2月份之外,毛竹林月均Ts均小于裸地,毛竹林在夏天降低Ts 2~4 ℃,冬季升高Ts约1 ℃,这与马鸿儒等[21]研究结果有所不同,可能与研究地域不同有关。Ts主要受Tr影响。本研究结果表明,冬季,毛竹林内Tr低于裸地,但毛竹林内Ts却高于裸地。土壤温度变化受许多因子影响[24]。枯落物覆盖、土壤湿度、微生物含量等均能对其产生影响。林内土壤湿度越高,土壤微生物含量就越高,而土壤微生物呼吸活动能释放大量热量,枯枝落叶物分解的同时也能释放大量热量,加上枯枝落叶物对土壤的遮挡,阻挡了土壤与空气的热量传递,虽然说气温是土壤温度的主要影响因子,但是在土壤微生物、土壤湿度及枯枝落叶物的共同作用下,气温是否依然占据主导地位,还有待研究。
同一土层,林内Ts出现最值的时刻比裸地晚,毛竹林Ts最大值始终比裸地小,最小值始终比裸地大,表明,毛竹林具有缓温的能力,可以滞后Ts最值的时间30~90 min,这与林兴军[25]研究结果一致,林内由于冠层的阻挡作用,使林内Ts变化较为平缓,使Ts增温慢,散热也慢。
[1] 王淼,韩士杰,王跃思.影响阔叶红松林土壤CO2排放的主要因素[J].生态学杂志,2004,23(5):24-29.
[2] GUAN D X, WU J B, YU G R, et al. Meteorological control on CO2flux above broad-leaved Korean pine mixed forest in Changbai Mountains[J]. Science in China (Series D Earth Sciences),2005,48(1):116-122.
[3] GUAN D X, WU J B, ZHAO X S, et al. CO2fluxes over an old, temperate mixed forest in northeastern China[J]. Agricultural and Forest Meteorology,2006,137(3):138-149.
[4] WU J, GUAN D, WANG M, et al. Year-round soil and ecosystem respiration in a temperate broad-leaved Korean Pine forest[J]. Forest Ecology and Management,2006,223(1):35-44.
[5] 杜颖,关德新,殷红,等.长白山阔叶红松林的温度效应[J].生态学杂志,2007,26(6):787-792.
[7] LUXHØI J, FILLERY I R P, RECOUS S, et al. Carbon and N turnover in moist sandy soil following short exposure to a range of high soil temperature regimes[J]. Australian Journal of Soil Research,2008,46(8):710-718.
[8] 王俊峰.长江源区沼泽与高寒草甸生态系统变化及其碳平衡对全球气候变化的响应[D].兰州:兰州大学,2008.
[9] 周国模,姜培坤.毛竹林的碳密度和碳贮量及其空间分布[J].林业科学,2004,40(6):20-24.
[10] 漆良华,张旭东,周金星,等.湘西北小流域不同植被恢复区土壤微生物数量、生物量碳氮及其分形特征[J].林业科学,2009,45(8):14-20.
[11] 肖复明,范少辉,汪思龙,等.毛竹、杉木人工林生态系统碳平衡估算[J].林业科学,2010,46(11):59-65.
[12] HORTON B, CORKREY R. A weighted coefficient model for estimation of Australian daily soil temperature at depths of 5cm to 100cm based on air temperature and rainfall[J]. Soil Research,2011,49(4):305.DOI:10.1071/sr10151.
[13] 孙金伟,吴家兵,关德新,等.森林与空旷地空气温湿度及土壤温度的长期对比研究[J].生态学杂志,2011,30(12):2685-2691.
[14] 陈继康,李素娟,张宇,等.不同耕作方式麦田土壤温度及其对气温的响应特征:土壤温度日变化及其对气温的响应[J].中国农业科学,2009,42(7):2592-2600.
[15] 侯建花,李桥,徐小军,等.利用气温进行统计回归预测土温[J].浙江林业科技,2015,35(5):55-59.
[16] 李沙,刘冬梅,孙辉.西南亚高山森林土壤表层温度变化特征及拟合模型[J].西北林学院学报,2013,28(3):34-39.
[17] 曾小平,赵平,饶兴权,等.鹤山丘陵3种人工林叶面积指数的测定及其季节变化[J].北京林业大学学报,2008,30(5):33-38.
[18] 陈宏志,胡庭兴,龚伟,等.我国森林小气候的研究现状[J].四川林业科技,2007,28(2):29-32,28.
[19] 徐云蕾,曾馥平,宋同清,等.喀斯特峰丛洼地次生林小气候特征研究[J].农业现代化研究,2012,33(2):239-244.
[20] 张一平,何云玲,钟水新,等.岷江上游山地森林生态气候效应[J].山地学报,2005,23(3):300-307.
[21] 马鸿儒,吉春容,李新建,等.天山中部白杨沟天然林区森林小气候观测与分析[J].干旱区研究,2011,28(2):251-254.
[22] 贾傅.南昌梅岭木荷生物防火林带小气候及土壤养分研究[D].南昌:江西农业大学,2014.
[23] 张慧智,史学正,于东升,等.中国土壤温度的季节性变化及其区域分异研究[J].土壤学报,2009,46(2):227-234.
[24] 杨雅红.土壤温度的日变化及影响因子分析[J].农业与技术,2014,34(4):19-20.
[25] 林兴军.不同水肥对日光温室番茄品质和抗氧化系统及土壤环境的影响[D].北京:中国科学院研究生院,2011.
杨丹,女,1994年11月生,南京林业大学林学院,硕士研究生。E-mail:332090694@qq.com。
庄家尧,南京林业大学林学院,副教授。E-mail:nlzjiayao@njfu.edu.cn。
2016年12月5日。
S795.9;S716.2
1)江苏省高校自然科学研究重大项目(16KJA220003)。
责任编辑:程 红。
Regulating Air and Soil Temperature Characteristics of inside and outsidePhyllostachysedulisForest Stands in Nanjing Suburbs//Yang Dan(Nanjing Forestry University, Nanjing 210037, P. R. China); Yang Jing(Science Press); Zhang Jinchi, Zhuang Jiayao, Xue Xue, Cheng Jurong(Nanjing Forestry University)//Journal of Northeast Forestry University,2017,45(3):68-73.
The experiment was conducted to study the changes of air temperature and soil temperature inside and outside thePhyllostachysedulisforest and the effect ofP.edulisforest on the air temperature and soil temperature. Decagon micro-meteorological monitoring system was used to conduct long-term monitoring on solar radiation, air temperature, soil temperature during the depth from 0 cm to 40 cm and other factors ofP.edulisforest in Nanjing suburbs. With the air and soil temperature characters inside theP.edulisforest and the effect ofP.edulisforest in Nanjing suburbs, air and soil temperature regulation were analyzed. The solar radiation had a multi-peak curve, and theP.edulisforest had the effect of decreasing the solar radiation, especially in summer and autumn. The trend of air temperature of the nudation was the same as that of theP.edulisforest, fluctuated along a single-peak curve, and theP.edulisforest had the effect of increasing the air temperature in the day time and decreasing in the evening, theP.edulisforest had the effect of decreasing the air temperature (by 0.5 ℃) all year round. The fleet soil temperature diurnal curve as Trigonometric function image, and with the increase of soil depth, the diurnal variation of soil temperature was smaller, in spring, summer and winter. The soil temperature in the same soil layer and the nudation soil were higher than those inP.edulisforest, and the difference was not significant with the increase of the soil depth. The seasonal variation curve of soil temperature was consistent with the seasonal variation curve of forest temperature.P.edulisforest could reduce the soil temperature (by 1 ℃) in summer and increase the soil temperature (by 2-4 ℃) in winter. The time of the maximum and minimum temperature of soil was lagged 30-90 min byP.edulisforest. The solar radiation, air temperature and soil temperature in the nudation land andP.edulisforest were extremely significant (P<0.01).P.edulisforest has a very significant role in reducing solar radiation and air temperature, the remission of soil temperature and soil temperature regulation.
