鲤春病毒血症病毒Shlj1株糖蛋白空间结构及其B细胞抗原表位的预测
2017-07-24纪锋徐黎明赵景壮刘淼卢彤岩贺文斌尹家胜
纪锋,徐黎明,赵景壮,刘淼,卢彤岩,贺文斌,尹家胜
(1.中国水产科学研究院黑龙江水产研究所,哈尔滨150070;2.上海海洋大学水产与生命学院,上海201306)
鲤春病毒血症病毒Shlj1株糖蛋白空间结构及其B细胞抗原表位的预测
纪锋1、2,徐黎明1,赵景壮1,刘淼1,卢彤岩1,贺文斌2,尹家胜1
(1.中国水产科学研究院黑龙江水产研究所,哈尔滨150070;2.上海海洋大学水产与生命学院,上海201306)
为预测鲤春病毒血症病毒(Spring Viremia of Carp Virus,SVCV)Shlj1株糖蛋白(Glycoprotein,G)的空间结构及B细胞抗原表位,采用RT-PCR法从接种SVCV的鲤上皮细胞悬液提取的病毒总RNA中扩增糖蛋白基因,并通过序列测定获得Shlj1株糖蛋白基因的核苷酸序列及氨基酸序列,利用Phyre2对Shlj1分离株的糖蛋白空间结构进行预测,使用DNAStar软件综合分析糖蛋白的柔性区域、亲水性、表面可能性和抗原指数参数。结果表明:获得的SVCV Shlj1株糖蛋白核苷酸序列长1527 bp,推导出其氨基酸序列为509 aa;空间结构预测显示,SVCV Shlj1株糖蛋白存在一定数量的α-螺旋、β-折叠、β-转角及大量无规则卷曲结构,空间构象较规则;抗原指数及抗原表位指数分析显示,糖蛋白中存在许多抗原指数较高的区域,该区域平均抗原指数为1.025,最大值达1.250,其中5个区域(4~26、145~157、293~302、315~327、407~491氨基酸区段)可能是B细胞抗原表位的主要分布区域。本研究结果为SVCV糖蛋白表位疫苗的设计及单克隆抗体的制备奠定了理论基础。
鲤春病毒血症病毒;糖蛋白;空间结构;B细胞抗原表位
鲤春病毒血症(Spring siremia of carp,SVC)是由鲤春病毒血症病毒(Spring Viremia of Carp Virus,SVCV)引起的流行于鲤科鱼类间的一种急性高度传染性疾病,一旦感染可以引起鲤Cyprinus carpio大规模暴发疫情,具有高度的传染性,感染SVCV的仔鱼死亡率最高可达90%[1],该病曾广泛传播于欧洲和亚洲的养殖鲤中,并给当地鲤养殖业造成了重大损失。中国自2001年在全国范围开展SVCV监测工作以来,陆续出现了一些关于SVCV分离鉴定的研究报道[2-3]。本课题组在2015年对黑龙江省40个养殖场送检的鲤进行病原监测过程中,分离获得一株SVCV,命名为Shlj1(No. KX911973)。SVCV首先由Fijan[4]于20世纪70年代分离出来,该病毒属于水泡性病毒属,是一种弹状病毒,病毒粒子一端为圆弧形,另一端较平整,病毒粒子长为(135±45)nm,宽为(75±15) nm[5-6],基因组由11 019个核苷酸残基组成[7],与核蛋白(N)螺旋对称结合,形成直径约为50 nm的核衣壳,少量的聚合酶(L)和磷蛋白(P)结合在核衣壳上;由脂质和蛋白质组成的囊膜外表面有突起的糖蛋白(G),内表面为基质蛋白(M)[5]。
糖蛋白是一种包含509个氨基酸残基的典型跨膜蛋白,是SVCV病毒分子非常重要的免疫原性蛋白,参与鲤春病毒血症病毒的入侵和免疫应答,决定SVC病毒的血清学特征,是诊断试剂开发的主要靶标[8-9]。目前,SVCV糖蛋白在实验室条件下还不能做到大批量表达[2,10],而小分子蛋白相比于大分子蛋白更易于做到大量表达[11]。为此,本研究中克隆了SVCV Shlj1分离株的糖蛋白基因,获得了糖蛋白的编码基因序列及氨基酸序列,并应用生物信息学软件对糖蛋白序列进行了三维结构及B细胞抗原表位预测,旨在为SVCV Shlj1株糖蛋白单克隆抗体的制备、基因工程疫苗的设计和免疫诊断技术研发提供理论依据。
1 材料与方法
1.1 材料
鲤上皮瘤细胞(Epithelioma papulosum cyprini, EPC)由中国水产科学研究院长江水产研究所鱼类病害教研室曾令兵教授惠赠。SVCV Shlj1为本课题组对黑龙江地区养殖鲤进行流行病学调查过程中利用EPC从送检的鲤体内分离获得,经测定病毒滴度为106.28TCID50/mL。
RNA提取试剂盒SV Total RNA Isolation system为Promega生物公司产品;一步法基因扩增试剂盒、DNA聚合酶rTaq premix、DNA Marker和PCR纯化试剂盒均为TaKaRa公司产品。
1.2 方法
1.2.1 引物设计及合成 用于SVCV糖蛋白基因序列克隆的引物为Gf:5′GAAAAAAACTAACAGACATCATGTCTAT 3′,Gr:5′CTCAAACGAAGGACCGCATTTCGTG 3′,由哈尔滨博仕生物公司合成。
1.2.2 SVCV糖蛋白基因的扩增及序列测定 取保存于冰箱(-80℃)中的SVCV病毒悬液200 μL,利用SV Total RNA Isolation system试剂盒提取病毒总RNA。以提取的SVCV总RNA为模板,Gf/ Gr为引物,进行目的蛋白基因的一步法RT-PCR扩增[12],以未经SVCV感染的EPC细胞提取的RNA为阴性对照组。PCR反应程序为:50℃下反应30 min;94℃下预变性5 min;94℃下变性30 s;55℃下退火30 s,72℃下延伸30 s,共进行30个循环;最后在72℃下再延伸10 min。扩增产物在110 V电压下用10 g/L琼脂糖凝胶电泳35 min,并对目的条带进行胶回收,将胶回收产物送至上海生工生物公司进行测序。分析测序结果,利用DNAStar软件中的EditSeq程序推导糖蛋白的氨基酸序列。
1.2.3 SVCV Shlj1株糖蛋白三维结构的预测 应用在线工具Phyre2(http://www.sbg.bio.ic.ac. uk/phyre2/html/page.cgi?id=index)对SVCV Shlj1株糖蛋白的三维结构进行预测,并用Chimera蛋白分析软件进行模拟分析。
1.2.4 SVCV Shlj1株糖蛋白的二级结构以及抗原表位分析 应用DNAStar软件中的Protean模块对SVCV Shlj1株糖蛋白的B细胞抗原表位进行分析:首先用模块预测其二级结构(Chou-Fasman法和Gamier-Robson法)、柔性区段(Karplus-Schulz法)、表面可能性(Emini法);再用Jameson-Wolf法和Kolaskar-Tongaonkar法(http://tools.immuneepitope.org/tools/bcell/iedb_input)分别预测其抗原指数和抗原表位指数,用Kyte-Doolittle法分析其亲水性;最后,结合糖蛋白的三维结构预测其B细胞抗原表位。
2 结果与分析
2.1 病毒糖蛋白基因片段的扩增
扩增试验中以SVCV病毒悬液提取的RNA为模板,Gf/Gr为引物,进行一步法RT-PCR基因扩增,将扩增产物进行琼脂糖凝胶电泳,结果出现与目的条带大小一致的特异性条带(图1)。

图1 SVCV Shlj1株糖蛋白基因PCR扩增产物的凝胶电泳分析Fig.1 Gel electrophoresis of PCR product of glycoprotein gene in SVCV Shlj1 strain
2.2 糖蛋白空间三维结构预测
利用在线工具Phyre2对糖蛋白的三维结构进行预测,结果显示:其结构中存在一定数量的α-螺旋、β-折叠、β-转角和大量无规则卷曲结构,其中,分布于蛋白内部的α-螺旋区域包含大量疏水氨基酸;β-折叠形成的桶状结构处于蛋白的中心,其内部氨基酸含有大量的疏水性残基,与α-螺旋共同构成一个疏水核心区域;连接着α-螺旋与β-折叠的β-转角和无规则卷曲一并位于蛋白的外部(图2)。
2.3 SVCV Shlj1株糖蛋白的二级结构
糖蛋白的二级结构分析预测结果发现,应用Garnier-Robson法和Chou-Fasman法两种方法预测的α-螺旋、β-折叠和β-转角存在较多的重叠区域。Gamier-Robson法预测的α-螺旋呈弥散状分布,数量较少,其中192~203区段出现跨区最大的α-螺旋(12 aa),另外2个α-螺旋包含的氨基酸数目相对较少,分别只有7个氨基酸;β-折叠数目较多,大部分由多个氨基酸组成且分布均匀,其中跨区最大的β-折叠出现在213~230和465~ 482区段,分别由18个氨基酸组成;β-转角区域集中在氨基酸序列的中前段,其中81~91区段最大,由11个氨基酸组成,也偶见有单个氨基酸出现。Chou-Fasman法预测的α-螺旋数量相对较多,分布相对均匀,其中186~203区段出现最大的α-螺旋(18 aa),而且没有单个氨基酸构成α-螺旋的情况出现,其中与Garnier-Robson法预测的有部分区域重叠;β-折叠分布相对比较均匀,与Gamier-Robson法预测的结果有比较多的重叠区域,但组成β-折叠的氨基酸数量比Gamier-Robson法预测的少(图3、表1和表2)。Karplus-Schulz法预测的SVCV Shlj1株糖蛋白柔性区域位置为:14~17、24~29、47~51、56~66、68~73、83~85、104~111、118~125、127~131、138~144、156~158、167~174、179~181、188~190、201~202、209~214、233~235、243~261、272~283、303~317、325~330、337~347、353~371、380~383、392~398、408~409、413、430~438、442~451、460~465、469~471、493~503,可见SVCV Shlj1株糖蛋白有较多个柔性区域,因此,有较多个潜在的抗原表位存在(图4)。
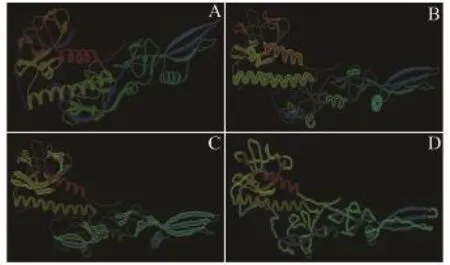
图2 SVCV Shlj1株糖蛋白的结构预测Fig.2 Protein structure prediction of glycoprotein in SVCV Shlj1 strain

图3 不同方法预测SVCV Shlj1株糖蛋白的二级结构Fig.3 Secondary structure of glycoprotein in SVCV Shlj1 strain predicted by various methods

图4 Karplus-Schulz法预测SVCV Shlj1株糖蛋白的柔性区段Fig.4 Flexible regions of glycoprotein in SVCV Shlj1 strain predicted by Karplus-Schulz method
2.4 SVCV Shlj1株糖蛋白的亲水性和表面可能性
氨基酸亲水性预测(Kyke-Doolittle法)结果显示,糖蛋白亲水性较强,亲水性区段占有较大比例且整体分布比较均匀,其中靠近羧基端的结构蛋白所跨区段最大、具有最高的亲水性指数(图5),指示该区段很有可能位于蛋白表面,极有可能作为病毒的抗原表位。SVCV Shlj1株糖蛋白的表面可能性(Emini法)分析结果显示,140~146、301~314、336~347、361~370、392~397、413~415、 426~437、443~449、491~505区域的表面可能性较大,因此,这些极有可能位于蛋白表面区域,可作为B细胞的优势区域(图6)。
2.5 SVCV Shlj1株糖蛋白的抗原指数
抗原指数(Jameson-Wolf法)预测结果显示, SVCV Shlj1株糖蛋白中存在大量抗原指数较高的区域,如68~73、82~89、106~111、119~126、166~181、209~214、270~284、303~310、337~346、362~370、377~383、392~398、426~453、491~507形成抗原表位的可能性也较大,抗原指数最高的区段特别分布在近羧基端区域,其中231~259区段所跨区域最大,因此,该区段极有可能存在优势抗原表位(图7)。
2.6 SVCV Shlj1株糖蛋白的抗原表位指数
糖蛋白抗原表位(Kolaskar-Tongaonkar法[13])预测结果显示,SVCV Shlj1株糖蛋白的平均抗原指数为1.025,最大值为1.250,最小值为0.834 (图8)。
2.7 SVCV Shlj1株糖蛋白的B细胞表位预测结果综合分析
以SVCV Shlj1株糖蛋白序列的抗原指数、亲水性指数、柔性区域、蛋白质表面可能性指数等抗原表位参数以及抗原表位指数预测结果,如果抗原指数≥1.025,表面可能性指数≥1,亲水性指数为非负数值,同时其内部结构或附近又包含有柔性区域,包含有不少于7个氨基酸残基,则这一区段极有可能作为表位区域。从符合这些条件的区域中共筛选出5个优势抗原表位区段,其氨基酸序列如表3所示。

表1 Gamier-Robson法预测的SVCV Shlj1株糖蛋白的二级结构Tab.1 Secondary structure of glycoprotein in SVCV Shlj1 strain predicted by Gamier-Robson method

表2 Chou-Fasman法预测的SVCV Shlj1株糖蛋白的二级结构Tab.2 Secondary structure of glycoprotein in SVCV Shlj1 strain predicted by Chou-Fasman method
3 讨论
3.1 抗原表位的作用
整个蛋白质抗原分子很难被免疫细胞表面受体所识别,而仅能够被位于抗原分子表面的决定抗原特异性的数个氨基酸残基组成的特殊序列所识别,这段特殊序列即为抗原表位(Epitope),也被称为抗原决定簇(Antigenic determinant),是抗原肽分子上具有一定空间结构的某一特定区段。严格说,抗体的特异性是针对大分子表面的抗原表位,而不是整个抗原分子[14]。表位一般由5~7个氨基酸和单糖残基组成,氨基酸残基的个数最多不会超过30个,且多位于蛋白表面的β-转角和无规则卷曲等比较松散、易扭曲的区段。想要了解抗原的结构与功能、抗原抗体反应等有关免疫反应的相关信息,以及研发诊断试剂、合成药物和开发疫苗等,就必须要做到正确分析和确定抗原表位[15]。有研究证实,蛋白质的序列或结构特征在一定程度上影响着抗原表位生物学特性[8,16]。本研究中获得了SVCV Shlj1株糖蛋白的核苷酸序列(1527 bp)并推导出其氨基酸序列(509 aa);空间结构预测结果显示,SVCV Shlj1株糖蛋白存在一定数量的α-螺旋、β-折叠、β-转角及大量无规则卷曲结构,空间构象较规则。鉴于糖蛋白在该病毒中起到主要免疫原作用,所以对SVCV Shlj1株糖蛋白的研究为后期工作的开展具有更好地指导作用。

图5 SVCV Shlj1株糖蛋白的亲水性分析Fig.5 Hydrophilicity of glycoprotein in SVCV Shlj1 strain
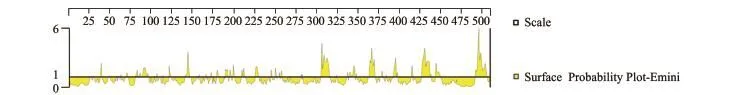
图6 SVCV Shlj1株糖蛋白的表面可能性分析Fig.6 Surface probability of glycoprotein in SVCV Shlj1 strain
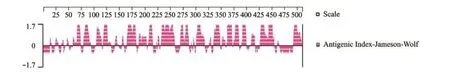
图7 SVCV Shlj1株糖蛋白的抗原指数分析Fig.7 Antigenic index of glycoprotein in SVCV Shlj1 strain
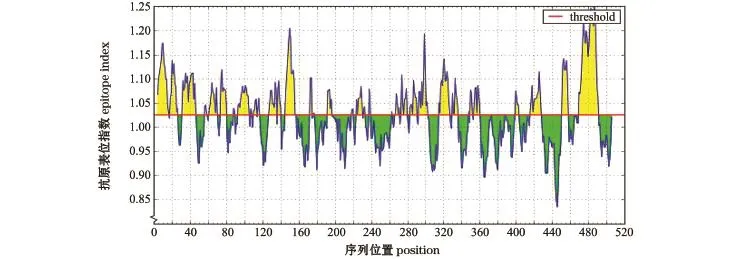
图8 SVCV Shlj1株糖蛋白的抗原表位指数分析Fig.8 Antigenic epitope index of glycoprotein in SVCV Shlj1 strain
3.2 准确预测SVCV糖蛋白抗原表位的重要性
SVCV粒子呈子弹状,编码5种主要的蛋白,其中跨膜糖蛋白G具有主要的免疫原性,决定了病毒的血清学特征,对病毒的入侵和参与免疫应答等特性起到关键性作用,是诊断试剂开发的重要目标蛋白[8]。目前,SVCV糖蛋白在实验室条件下还不能做到大批量表达,真核表达效率也非常低,但有原核表达系统生产抗原且获得成功的相关报道[17-19]。在原核表达系统中,翻译后加工修饰体系与真核表达系统相比并不是很完善,可能不存在翻译后的修饰,但表达产物却不失免疫原性[11,20],可能是抗原表位起到了决定作用。作为疫苗,小分子蛋白相比于大分子蛋白易于做到大量表达,相比于减毒活疫苗又能做到安全可靠。所以,准确地预测抗原表位,通过原核表达抗原表位基因的方法来大量获取具有免疫原性的目的蛋白,也同样促进了多肽疫苗的合成、诊断试剂的制备和单克隆抗体的筛选。用多种方法和非单一参数的综合预测抗原表位具有更高的可信度,提高了预测结果的准确性。本研究中,抗原指数以及抗原表位指数等参数分析结果显示,糖蛋白中存在许多抗原指数较高的区域,这些区域平均抗原指数为1.025,最大值达1.250,其中4~26、145~157、293~302、315~327、407~491氨基酸区段可能是B细胞抗原表位的主要分布区域。
3.3 抗原表位与多种因素之间的关系
早在20世纪末,Hopp等[21]就已经提出了通过氨基酸残基亲水性来预测抗原表位,至此,运用单一参数和多参数预测模型均有成功做到预测某种类型蛋白抗原表位的案例[22-24]。虽然亲水性和表面可能性都是形成表位很重要的条件之一,但是为了逃避宿主免疫检测机制导致抗原表位往往是高变的特质,所以仅仅满足其中任何一种条件并不能表示一定具有抗原性,因此,单参数预测有其局限性。多种因素相互影响决定表位抗原性,多种因素综合作用是表位形成的重要条件,很大程度上抗原表位生物学特性会受到蛋白质二级结构的影响。正确剖析和确定表位不仅能够对抗原的结构与功能、抗原抗体反应等信息有更进一步地了解,而且对诊断试剂的研发制备、药物的筛选合成和多肽疫苗设计生产等也具有重要的理论指导意义。
本研究中成功地对鲤春病毒血症病毒Shlj1株糖蛋白基因进行了克隆及序列测定,并分析了糖蛋白的二级结构、亲水性、表面可能性、柔性区段与抗原性指数等多种参数;并以多个单因素为基础,对糖蛋白的B细胞抗原表位进行了综合性分析,再利用二级结构预测结果筛选抗原表位,最后确定了SVCV Shlj1株糖蛋白优势表位。本研究结果可为SVCV Shlj1株糖蛋白单克隆抗体的制备、表位疫苗的设计和研究SVCV机制奠定了理论基础。
[1] Baudouy A M,Danton M,Merle G.SVCV infection of carp (author's transl)[J].Annales de Recherches Veterinaires, 1980,11(3):245-249.
[2] 刘荭,付峰,黄倢,等.鲤春血症病毒中国分离株糖蛋白基因和氨基酸序列的初步解析[J].中国病毒学,2005,20(6):647-651.
[3] Liu H,Gao L,Shi X,et al.Isolation of spring viraemia of carp virus (SVCV)from cultured koi(Cyprinus carpio koi)and common carp(Cyprinus carpio carpio)in PR China[J].Bulletin of the European Association of Fish Pathologists,2004,24(4):194-202.
[4] Fijan N.Spring viraemia of carp and other viral diseases and agents of warm-water fish[M]//Woo P T K,Bruno D W.Fish Diseases and Disorders 3:Viral,Bacterial and Fungal Infections.Wallingford:CAB International,1999:177-244.
[5] Ahne W,Bjorklund H V,Essbauer S,et al.Spring Viremia of Carp (SVC)[J].Diseases of Aquatic Organisms,2002,52(3):261-272.
[6] Wolf K.Fish Viruses and Fish Viral Diseases[M].New York:Cornell University Press,1988.
[7] Hoffmann B,Schütze H,Mettenleiter T C.Determination of the complete genomic sequence and analysis of the gene products of the virus of Spring Viremia of Carp,a fish rhabdovirus[J].Virus Research,2002,84(1-2):89-100.
[8] Björklund H V,Higman K H,Kurath G.The glycoprotein genes and gene junctions of the fish rhabdoviruses Spring Viremia of Carp Virus and hirame rhabdovirus:analysis of relationships with other rhabdoviruses[J].Virus Research,1996,42(1-2):62-80.
[9] 罗培骁,张琪,王敏,等.鲤春病毒血症病毒G蛋白的原核表达及多克隆抗体的制备[J].水生态学杂志,2014,35(4):81-86.
[10] 张家林,李强,叶仕根,等.鲤春病毒血症病毒糖蛋白的原核表达及单克隆抗体的制备[J].大连海洋大学学报,2014,29 (5):454-458.
[11] 徐进,周勇,陈倩,等.鲤春病毒血症病毒糖蛋白的截短表达及其免疫原性分析[J].中国水产科学,2016,23(2):352-358.
[12] Koutná M,Vesely T,Psikal I,et al.Identification of spring viraemia of carp virus(SVCV)by combined RT-PCR and nested PCR[J].Diseases of Aquatic Organisms,2003,55(3):229-235.
[13] Kolaskar A S,Tongaonkar P C.A semi-empirical method for prediction of antigenic determinants on protein antigens[J].FEBS Letters,1990,276(1-2):172-174.
[14] 李海侠,毛旭虎.蛋白质抗原表位研究进展[J].微生物学免疫学进展,2007,35(1):54-58.
[15] Feltkamp M C W,Smits H L,Vierboom M P M,et al.Vaccination with cytotoxic T lymphocyte epitope-containing peptide protects against a tumor induced by human papillomavirus type 16-transformed cells[J].European Journal of Immunology,1993,23(9):2242-2249.
[16] Duan L,Lei P,Yumei X,et al.Prediction and identificationbased prediction of Chinese hepatitis C viral-specific cytotoxic T lymphocyte epitopes[J].J Med Virol,2011,83(8):1315-1320.
[17] 张家林.鲤春病毒血症病毒G蛋白的原核表达及其单克隆抗体的制备[D].大连:大连海洋大学,2014.
[18] 付峰,刘荭,范万红,等.鲤春病毒血症病毒糖蛋白基因的克隆及其在毕赤酵母中的初步表达[J].海洋水产研究,2007, 28(4):72-76.
[19] 张琳,丁雅苓,陈建民,等.中国鲤鱼春季病毒血症毒株糖蛋白基因的亚克隆表达与纯化[J].中国兽医杂志,2011,47 (7):10-12.
[20] 兰文升,刘荭,高隆英,等.鲤春病毒血症病毒糖蛋白的高效表达和纯化[J].中国动物传染病学报,2010,18(2):18-22.
[21] Hopp T P,Woods K R.Prediction of protein antigenic determinants from amino acid sequences[J].Proceedings of the National Academy of Sciences of the United States of America,1981,78 (6):3824-3828.
[22] Kosmopoulou A,Vlassi M,Stavrakoudis A,et al.T-cell epitopes of the La/SSB autoantigen:prediction based on the homology modeling of HLA-DQ2/DQ7 with the insulin-B peptide/HLADQ8 complex[J].Journal of Computational Chemistry,2006,27 (9):1033-1044.
[23] 闫红,徐发良,顾长国,等.MD-2的B细胞抗原表位预测[J].免疫学杂志,2006,22(1):17-19.
[24] Sekhavati M H,Heravi R M,Tahmoorespur M,et al.Cloning,molecular analysis and epitopics prediction of a new chaperone Gro-EL Brucella melitensis antigen[J].Iranian Journal of Basic Medical Sciences,2015,18(5):499-505.
Predication of spatial structure and B cell antigenic epitope of glycoprotein in Shlj1 strain of Spring Viremia of Carp Virus
JI Feng1,2,XU Li-ming1,ZHAO Jing-zhuang1,LIU Miao1,LU Tong-yan1,HE Wen-bin2,YIN Jia-sheng1
(1.Heilongjiang River Fisheries Research Institute,Chinese Academy of Fishery Sciences,Harbin 150070,China;2.College of Fisheries and Life Science,Shanghai Ocean University,Shanghai 201306,China)
Glycoprotein gene was cloned from the total RNA extracted from the suspension of Shlj1 strain with Spring Viremia of Carp Virus(SVCV)using RT-PCR,nucleotide sequence of the glycoprotein was sequenced, and amino acid was deduced to predict the spatial structure and B cell antigenic epitope of SVCV glycoprotein.A three dimensional model of the glycoprotein was established based on the amino acid sequence,and by Phyre2,and flexible region,hydrophilicity,surface probability and antigenic index of the glycoprotein was analyzed by the Protean module in DNAStar software.It was found that the glycoprotein of the SVCV Shlj1 strain had nucleotide sequences of 1527 bp coding 509 amino acids.Space structure prediction revealed that there were a certain number of glycoprotein α-helix,β-pleated sheet,β-turn and a large number of random coil structure,with relatively regular spatial confirmation,in the SVCV Shlj1 strain.The analysis of parameters such as antigen index and antigen epitope index indicated that there were many fragments with high antigen index located in the glycoprotein,with average antigen index of 1.025,and the maximum value of 1.250.The potential B cell antigenic epitope regions were more likely located in 4-26,145-157,293-302,315-327,and 407-491 amino acid sites,indicating that the SVCV Shlj1 strain appeared to have five antigen epitope regions located in the glycoprotein.The findings provide a theoretical basis for preparation of monoclonal antibody and design of epitope vaccine against SVCV.
Spring Viremia of Carp Virus(SVCV);glycoprotein structural protein;spatial structure;B cell antigenic epitope
S941.41
A
10.16535/j.cnki.dlhyxb.2017.04.011
2095-1388(2017)04-0440-07
2016-10-10
黑龙江省应用技术研究与开发计划项目(GA13B401)
纪锋(1990—),男,硕士研究生。E-mail:fengjii@foxmail.com
尹家胜(1960—),男,研究员。E-mail:yinjiasheng@hrfri.ac.cn
