独角雪冰鱼和伯氏肩孔南极鱼骨骼中的元素分析
2017-07-24孙建飞刘洋许强华
孙建飞,刘洋、2,许强华、2、3、4
(1.上海海洋大学海洋科学学院,上海201306;2.大洋渔业资源可持续开发省部共建教育部重点实验室,上海201306;3.国家远洋渔业工程技术研究中心,上海201306;4.远洋渔业协同创新中心,上海201306)
独角雪冰鱼和伯氏肩孔南极鱼骨骼中的元素分析
孙建飞1,刘洋1、2,许强华1、2、3、4
(1.上海海洋大学海洋科学学院,上海201306;2.大洋渔业资源可持续开发省部共建教育部重点实验室,上海201306;3.国家远洋渔业工程技术研究中心,上海201306;4.远洋渔业协同创新中心,上海201306)
为探究缺失血红蛋白的南极独角雪冰鱼Chionodraco hamatus(CH)和具有血红蛋白的伯氏肩孔南极鱼Trematomus bernacchii(TB)骨骼中元素含量的差异,采用电感耦合等离子体质谱法对两种南极鱼的脊椎骨与头盖骨中Na、Mg、Al、K、Ca、56Fe、Zn等常量元素,以及V、Cr、Mn、As、Se、206Pb等微量元素含量进行了测定与分析。结果表明:两种南极鱼脊椎骨中Al、K、56Fe、Mn、Se含量有显著性差异(P<0.05),头盖骨中Mg、K、Ca、56Fe、V、Mn等元素含量有显著性差异(P<0.05),推测此差异可能与南极冰鱼缺失功能性血红细胞有关;将两种南极鱼脊椎骨骨骼中的元素含量与7种非南极海水鱼和3种淡水鱼相比,发现两种南极鱼骨骼中K、Na、Mg、56Fe、Zn、Ca等常量元素含量较少,且Mn、V、Cr、206Pb等微量元素含量明显低于非南极鱼,这可能与南极海域存在较低污染相关。本研究中首次测定了两种南极鱼骨骼中的元素含量,为后续拟通过体内组织元素分析探索南极鱼类的生理差异提供了科学依据。
独角雪冰鱼;伯氏肩孔南极鱼;常量元素;微量元素;骨骼
对抗冻蛋白AFPs(Antifreeze proteins)的研究,是在对南极鱼研究过程中取得的一项较重要的成果[1],几乎所有的南极鱼均存在着抗冻蛋白。在南极冰冻环境中的众多南极鱼类中,存在着一种极具研究价值的鱼类——鳄冰鱼科Channichthyidae鱼类[2],鳄冰鱼科又称南极冰鱼,隶属于南极鱼亚目Notothenioidei,其广泛分布于南极海域,在长期的适应性进化中逐渐丢失血红蛋白,并因无血红蛋白而成为硬骨鱼类中的独特类群,且是目前已知的唯一缺乏具有功能性血红细胞的脊椎动物,南极冰鱼主要依靠皮肤和鳃吸收溶解在水中的氧气[3]。
独角雪冰鱼Chionodraco hamatus(CH)是生活在南极深海零度以下无污染水域的底栖性鱼类。作为已知脊椎动物中唯一不含血红蛋白的类群,冰鱼的血液循环系统发生了巨大的补偿性生理变化[4]。伯氏肩孔南极鱼Trematomus bernacchii(TB)隶属于鲈形目、南极鱼亚目,其生存环境和南极冰鱼相似,且伯氏肩孔南极鱼和南极冰鱼具有较近的亲缘关系,但其体内存在正常表达的血红蛋白和红血球[5]。目前,南极冰鱼独特的形态学和生理学特征,成为研究低温环境中生物的功能基因进化、特别是低温下心血管系统发育的重要模型[6]。
各种常量元素和微量元素可直接或间接地参与机体几乎所有的生理和生化过程,是动物维持生命必不可少的营养素。如Zn、Fe、Mn等元素可在动物体内参与非常重要的生理功能。其中,Zn对蛋白质的合成具有重要意义,缺Zn会降低动物对氮的利用效率[7];Fe是红细胞中血红蛋白的组成成分,能够保证氧在机体内的正常运输,且在动物机体内还作为多种酶的组成成分影响能量代谢、蛋白质合成和免疫机能[8];Mn参与许多酶的合成与激活,可催化造血机能、调节内分泌、提高机体免疫功能[9]。由此可见,研究生物体内元素含量组成对了解生物体基本生理特征至关重要。
迄今为止,尚未见有关南极鱼体内元素含量测定的报道。为了解南极鱼适应环境的特殊生理机制,本研究中将首次测定缺失血红细胞的独角雪冰鱼和具有血红细胞的伯氏肩孔南极鱼脊椎骨和头盖骨中的元素含量,比较两种南极鱼骨骼中元素含量的差异及其与其他非南极鱼骨骼中各元素含量的差异,旨在明确南极鱼体内元素含量的特征,探索南极环境对南极鱼身体结构所产生的影响。
1 材料与方法
1.1 材料
试验用独角雪冰鱼和伯氏肩孔南极鱼由本实验室成员乘坐雪龙号从南极获取,在船上于超低温冰箱(-80℃)中保存,带回实验室后置于冰箱(-80℃)中保存。
试验用仪器主要有电感耦合等离子体质谱仪(ICP-MS)、电子天平、移液器、超纯水系统、硝解罐、鼓风干燥箱、移液枪等。试验用试剂主要有硝酸、双氧水、超纯水、高氯酸、四氟乙烯。
1.2 方法
1.2.1 试验设计 试验用样本分为独角雪冰鱼和伯氏肩孔南极鱼两组,每组3个样本,均为成年雄鱼。3只独角雪冰鱼的体长为(30±5)cm,体质量为(200±20)g;3只伯氏肩孔南极鱼体长为(15±2)cm,体质量为(80±5)g。分别将每组鱼的脊椎骨、头盖骨组织混合待测。
1.2.2 样品的消化 将鱼体浸置于开水中烫3 min后取出剥制,将分离得到的骨骼用双氧水漂白6 h左右,保存于5%的福尔马林溶液中或自封袋内[10]。准确称取骨骼0.5 g,置于容量为30 mL的瓷坩埚中,先放在电炉上碳化到不冒烟为止,然后迅速将其转移到550℃的消化炉中灰化6 h(灰化后的样品为灰白色)。取出后冷却至室温,再加入硝酸与高氯酸的混合酸(4∶1)10 mL,放置至少5 h(最好过夜);然后在电炉上用小火加热,蒸煮0.5 h,使灰化样品充分溶解直到黑色碳粒消失,但不能使溶液蒸干,必要时可以再滴加混合酸,直到见不到碳化颗粒。为除去样品液中的硝酸,可以再加10~20 mL的蒸馏水继续蒸煮15 min。冷却后用25 mL的容量瓶定容,待测。不加样品,用同样的方法获取的试液作空白对照[11]。
1.2.3 测定方法 本试验中,采用由国家海洋局所规定的电感耦合等离子体质谱法测定微量元素。其原理如下:以等离子体作为质谱离子源,样品雾化后以气溶胶的形式进入等离子体区域,经过蒸发、解离、原子化、电离等过程,被导入高真空的质谱部分,待测离子经质量分析器按质荷比的大小过滤分离后进入离子检测器,根据离子强度的大小计算得到样品中待测元素的浓度。
1.3 数据处理
试验数据采用SPSS 10.0软件进行统计处理,采用One-way ANOVA检验法对不同组织内元素含量的差异分别进行显著性分析。试验结果用平均值±标准差(mean±S.D.)表示,显著性水平设为0.05,极显著性水平设为0.01[12]。
2 结果与分析
试验所测定的元素种类包含:Be、Na、Mg、Al、K、Ca、V、Cr、Mn、56Fe、57Fe、Co、Ni、Cu、Zn、As、Se、Sr、Mo、Ag、Cd、Sb、Ba、Tl、206Pb、207Pb、208Pb、Th、U。由于部分元素不存在或并不具有独特性,或对于研究目标无意义,故可以舍去,仅保留Na、Mg、K、Ca、Al、56Fe、V、Cr、Mn、As、Se、206Pb用以后续分析。
2.1 两种南极鱼脊椎骨中的元素含量
分别取CH脊椎骨组织混合样品0.336、0.297、0.407 g,TB脊椎骨组织混合样品0.126、0.067、0.066 g,测定其中常量元素与微量元素含量。从表1可见:CH脊椎骨中常量元素含量依次为Ca>Na>K>Mg>Zn>56Fe>Al,TB脊椎骨中微量元素含量依次为Ca>K>Na>Mg>Zn>Al>56Fe;CH与TB脊椎骨中,Na、Mg、Ca、Zn含量无显著性差异(P>0.05),而56Fe含量有显著性差异(P<0.05),Al、K含量有极显著性差异(P<0.01)。
从表2可见:CH脊椎骨中微量元素含量依次为Mn>As>Se>206Pb>Cr>V,TB脊椎骨中微量元素含量依次为Mn>As>V>Cr,TB脊椎骨中Se和206Pb含量均为0;CH与TB脊椎骨中,Se含量有显著性差异(P<0.05),Mn含量有极显著性差异(P<0.01)。
2.2 两种南极鱼头盖骨中的元素含量
分别取CH头盖骨组织0.145、0.121、0.037 g,TB头盖骨组织0.123、0.048、0.135 g,测定其中常量元素与微量元素含量。从表3可见:CH头盖骨中常量元素含量依次为Ca>Na>K>Mg>Zn>56Fe>Al,TB头盖骨中常量元素含量依次为Ca>Na>K>Mg>Zn>56Fe=Al;CH与TB头盖骨中,仅Mg、K、Ca、56Fe含量有显著性差异(P<0.05)。
从表4可见:CH头盖骨中微量元素含量依次为Mn>As>206Pb>Se>Cr>V,TB头盖骨中微量元素含量依次为Mn>V>206Pb>As>Cr>Se;CH与TB头盖骨中,仅V和Mn含量有显著性差异(P<0.05)。
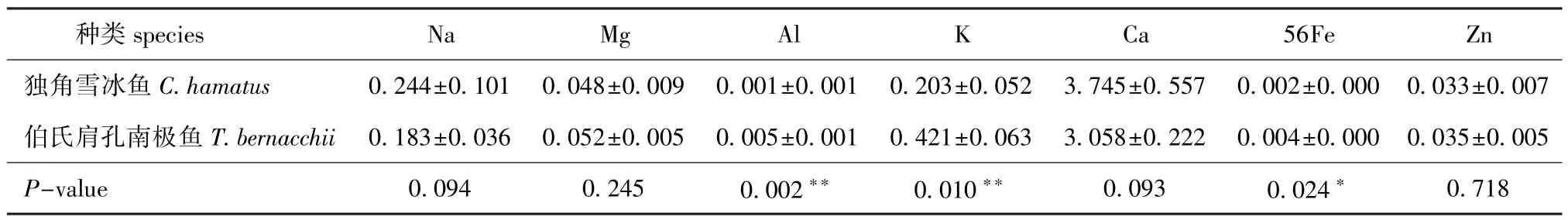
表1 独角雪冰鱼和伯氏肩孔南极鱼脊椎骨中常量元素Na、Mg、Al、K、Ca、56Fe、Zn的含量(n=3)Tab.1 Concentrations of macroelements Na,Mg,Al,K,Ca,56Fe,and Zn in the vertebrae of CH and TBmg/g
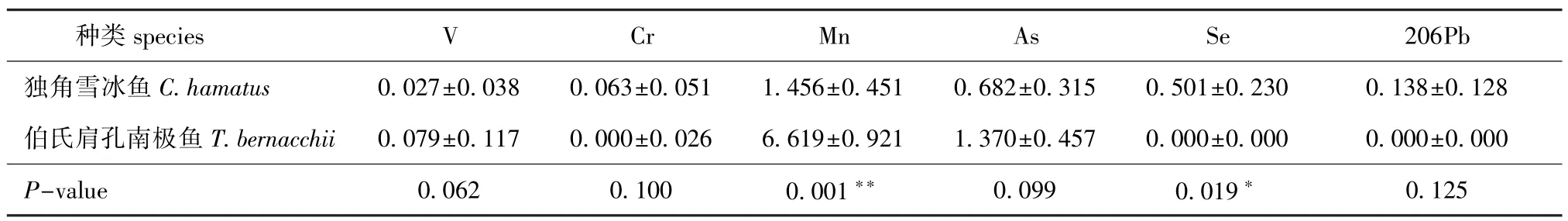
表2 独角雪冰鱼和伯氏肩孔南极鱼脊椎骨中微量元素V、Cr、Mn、As、Se、206Pb的含量(n=3)Tab.2 Concentrations of microelements V,Cr,Mn,As,Se,and 206Pb in the vertebrae of CH and TBμg/g
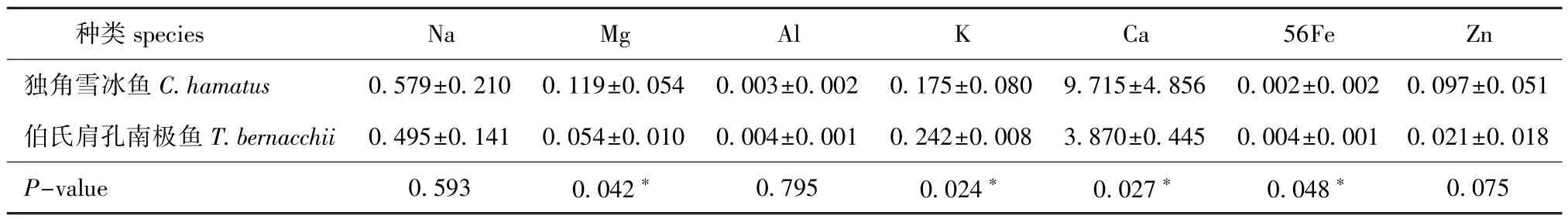
表3 独角雪冰鱼和伯氏肩孔南极鱼头盖骨中常量元素Na、Mg、Al、K、Ca、56Fe、Zn的含量(n=3)Tab.3 Concentrations of macroelements Na,Mg,Al,K,Ca,56Fe,and Zn in the skull of CH and TBmg/g

表4 独角雪冰鱼和伯氏肩孔南极鱼头盖骨中微量元素V、Cr、Mn、As、Se、206Pb的含量(n=3)Tab.4 Concentrations of microelements V,Cr,Mn,As,Se,and 206Pb in the skull of CH and TBμg/g
2.3 南极鱼和非南极鱼脊椎骨中元素含量的对比
南极鱼类长期生活在极端低温环境中,生理上已产生了一系列适应极端条件的适应性。为探究南极鱼类与其他非南极鱼类脊椎骨中所含元素的差异,将CH与TB脊椎骨中元素含量和淡水鱼类草鱼Ctenopharyngodon idellus[13]、鲶Silurus asotus Linnaeus[14]和鲫Carassius auratus[15],以及海水鱼类圆颌针鱼Tylosurus strongylurus[16]、金枪鱼Thunnus thynnus[17]、锤头双髻鲨Sphyrna zygaena、灰星鲨Mustelus griseus、赤魟Dasyatis akajei、班鳐Raja kenojei和孔鳐Rhinobatos porosa[18]脊椎骨中的元素含量进行比较。草鱼是常见的淡水经济鱼类,典型的草食性鱼类,属于鲤形目、鲤科、雅罗鱼亚科、草鱼属,其生长迅速,是中国淡水养殖的四大家鱼之一[19]。鲶属于鲶形目Siluriformes、鲶科Siluridae,是中国主要优质底栖淡水鱼类之一,广泛分布于国内的江河湖泊中[20-21]。鲫为常见淡水鱼,属于辐鳍鱼亚纲、鲤形目、鲤科、鲫属[22]。圆颌针鱼是上层远洋性洄游鱼类,喜欢栖息在河口海湾附近,常常随潮水游动,摄食小型鱼类和无脊椎动物[23]。金枪鱼属于硬骨鱼纲,主要分布于大西洋、印度洋、太平洋的热带地区,是大洋暖水性洄游鱼类[17]。锤头双髻鲨、灰星鲨、赤魟、班鳐和孔鳐等都属软骨鱼类[18]。南极鱼与其他非南极鱼脊椎骨中元素含量的比较结果见表5。
从表5可见:CH与TB脊椎骨中所含常量元素K、Na、Mg、56Fe、Zn含量明显比非南极鱼类少, Ca元素含量与草鱼和金枪鱼相比也显著地低,与几种软骨鱼类骨骼中Ca元素含量比较接近,这说明CH与TB等南极鱼脊椎骨骨骼的钙化程度比较低;此外,两种南极鱼(CH、TB)与7种海水鱼脊椎骨骨骼中所含重金属元素Mn、V、Cr明显少于淡水草鱼。
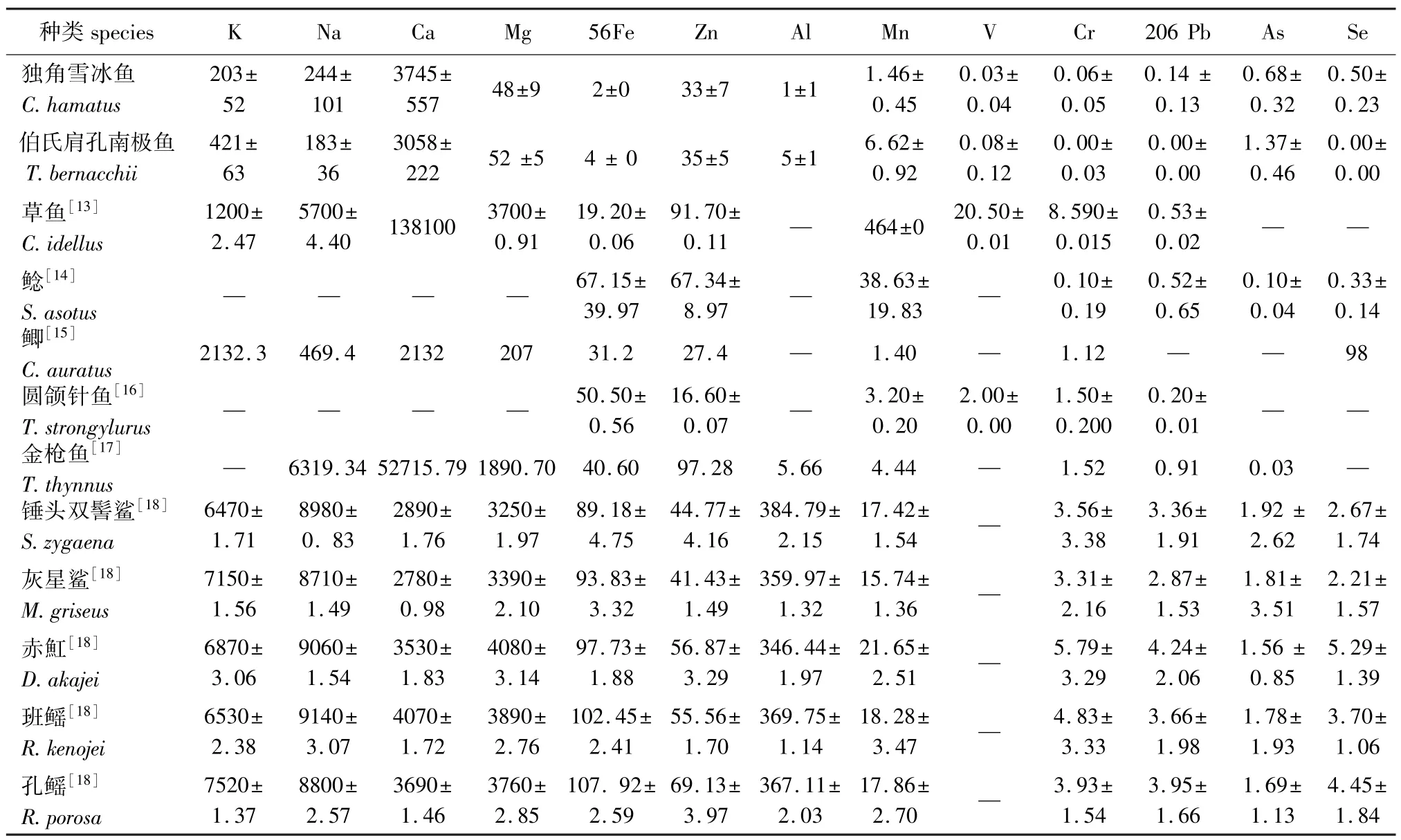
表5 南极鱼和非南极鱼类脊椎骨骨骼中元素含量的对比Tab.5 Comparisons of element concentrations in vertebrae between the Antarctic icefishes and non-Antarctic fishes mg/kg
3 讨论
3.1 两种南极鱼骨骼中常量元素含量的差异
独角雪冰鱼和伯氏肩孔南极鱼是生长在深海零度以下的无污染鱼类,栖息深度为400~600 m,两种南极鱼的生境接近,不同之处在于南极冰鱼体内中缺乏血红蛋白,也不产生功能性的血红细胞[24]。骨骼中常量元素测定结果显示(表1、表3),这两种鱼骨骼中的Ca、Na、K含量均较高。Ca是组成骨骼的主要成分,Na和K均是骨骼中电解质的主要成分,在维持骨骼细胞内外渗透压及酸碱平衡中起重要作用[25]。独角雪冰鱼体内的Na含量要高于伯氏肩孔南极鱼,而K含量则显著低于伯氏肩孔南极鱼(表1、表3)。根据钠钾泵的工作原理,生物体细胞内应该维持较低水平的K量和较高水平的Na量[26]。由于独角雪冰鱼体内含有较高水平的Na,可以推测认为,独角雪冰鱼的钠钾泵功能比伯氏肩孔南极鱼高。在细胞内,钠离子含量较高的情况下,生物动脉壁收缩加强,血管反应性加强,这些情况都有助于动脉血压升高[27]。因此,独角雪冰鱼的Na含量较高,可以推测认为,独角雪冰鱼内血压较高,血液循环相对较快。独角雪冰鱼由于缺乏运送氧气的血红蛋白,其体内较高的血压和较快的血液循环将有利于其从水中补充氧气,作为血红素缺失的补偿。
Ca可以维持肌肉和神经的正常兴奋性,参与细胞间的连接,还在细胞膜上调节受体结合和离子通透性[28]。独角雪冰鱼骨骼中的Ca含量高于伯氏肩孔南极鱼的原因之一,推测与独角雪冰鱼的生活习性有关,独角雪冰鱼常常会在水下数百米处摄食,由于南极水域水温较低,为维持肌肉和神经的正常兴奋性,需要垂体分泌相当数量的钙离子[29]。
独角雪冰鱼与伯氏肩孔南极鱼骨骼中56Fe的含量显著差异(表1、表3)。Fe是动物机体必需的微量元素之一,它的主要功能是构成血红蛋白,参与氧气的运输,是血红蛋白及细胞色素酶、过氧化物酶等的必需组成成分[30]。血红蛋白是红细胞中氧的载体,由于独角雪冰鱼内不含血红蛋白,所以Fe元素的含量极低。
3.2 两种南极鱼骨骼中微量元素含量的差异
独角雪冰鱼与伯氏肩孔南极鱼骨骼中除了常量元素含量存在差异外,其骨骼中的部分微量元素含量也存在着显著性差异(表2、表4)。如独角雪冰鱼头盖骨骼中V的含量显著低于伯氏肩孔南极鱼。钒在生物体内含量较低,但是对生理功能有着多方面的影响。高浓度的钒离子可以促进新陈代谢,刺激造血系统,促进生物体内血红素的形成,对于降血压有着明显的作用[31]。独角雪冰鱼体内缺乏血红素,也缺乏功能性血红细胞,推测这与冰鱼体内较低的钒含量相关。另一方面,独角雪冰鱼具有较大的血流输出,血压也相对较高,而钒具有降低血压的作用,这可能也与冰鱼体内具有较低钒含量相关[32]。
在两种南极鱼骨骼中这些微量元素中,Mn的含量明显高于其他微量元素。据报道,Mn主要在鱼体骨骼中蓄积。骨骼被认为是评价Mn生物利用率最敏感的组织。Mn元素在生物体中与造血过程紧密相关,它既是造血过程的原料,也对这个过程起到一定程度的调节作用,动物机体缺锰,可能产生骨骼畸形、骨软化及骨吸收增加等缺陷[33]。独角雪冰鱼骨骼中Mn元素含量较低,推测可能与其缺乏红细胞相关。另外,Se是维持心脏正常功能的重要元素,对心脏肌体有保护和修复作用[34]。独角雪冰鱼脊椎骨中含有Se元素,而伯氏肩孔南极鱼脊椎骨中不含Se,可以推测,独角雪冰鱼具有相对较强的免疫力。
3.3 南极鱼与非南极鱼脊椎骨元素含量的差异
独角雪冰鱼与伯氏肩孔南极鱼终年生长在极端寒冷的南极海域,其与非南极鱼脊椎骨骨骼中的元素含量差别较大(表5),以56Fe为例,两种南极鱼骨骼中56Fe的含量远低于其他几种非南极鱼。
本研究还发现,淡水鱼类(草鱼、鲶)脊椎骨骨骼中所含Zn元素含量明显高于几种海水鱼类(金枪鱼除外)。Zn的污染主要由汽车尾气排放、农药化肥滥用导致[35]。这表明,淡水鱼所生活的淡水环境比海洋环境受到的污染多,导致体内Zn的累积。本研究同时还发现,两种南极鱼脊椎骨骨骼中所含重金属元素Mn、V、Cr、Pb明显低于另外几种非南极鱼,这表明,远在南极海域的独角雪冰鱼和伯氏肩孔南极鱼,受到较轻的人类活动排放污染,体内重金属含量累积较少。
综上所述,独角雪冰鱼和伯氏肩孔南极鱼是生长于南极冰冻环境中极具研究价值的鱼类。本研究中首次测定了两种南极鱼脊椎骨、头盖骨中的元素含量,明确了南极鱼骨骼中元素含量的特征,初步探究了南极环境对南极鱼身体结构所产生的影响,这对于将来更好地研究和利用南极渔业资源具有重要意义。值得指出的是,本研究中仅测定了两种南极鱼脊椎骨、头盖骨中的元素含量,还不能完全反映出两种鱼生活习性的差异。为全方位探究独角雪冰鱼和伯氏肩孔南极鱼因元素含量差异而导致的一系列新陈代谢差异,还需测定更多组织的元素含量。
[1] Devries A L,Cheng C H C.Antifreeze proteins and organismal freezing avoidance in polar fishes[J].Fish Physiology Series, 2005,22(4):155-201.
[2] Cincinelli A,Martellini T,Vullo D,et al.Anion and sulfonamide inhibition studies of an α-carbonic anhydrase from the Antarctic hemoglobinless fish Chionodraco hamatus[J].Bioorganic&Medicinal Chemistry Letters,2015,25(23):5485-5489.
[3] 郭亚南,王丛丛,刘云,等.miR-210-5p对南极独角雪冰鱼心脏发育的作用机制研究[J].上海海洋大学学报,2016,25(4):481-487.
[4] Ferrerasa J M,Raguccib S,Citores L,et al.Insight into the phylogenetic relationship and structural features of vertebrate myoglobin family[J].International Journal of Biological Macromolecules, 2016,93:1041-1050.
[5] 王桂志,张永,张弛.等.独角雪冰鱼和伯氏肩孔南极鱼TRPV1基因的克隆及低温适应功能检测[J].大连海洋大学学报, 2016,31(2):124-129
[6] 刘秀荣.独角雪冰鱼和伯氏肩孔南极鱼Hsp70和Hsp90β基因的克隆、序列分析以及低温适应性研究[D].上海:上海海洋大学,2015.
[7] 郭岚.环境中的微量元素与人体健康[J].甘肃科技,2005,21 (8):110-112.
[8] 李清晓,李忠平.铁元素的营养作用及在动物生产上的应用[J].饲料博览,2004(4):4-6.
[9] 聂家全,董晓慧,谭北平,等.锰源和锰水平对斜带石斑鱼幼鱼生长、抗氧化功能和矿物元素沉积的影响[J].大连海洋大学学报,2015,30(2):196-202.
[10] 郭长江,赵文,石振广,等.达氏鳇幼鱼骨骼系统解剖研究[J].大连海洋大学学报,2016,31(3):301-306.
[11] 唐精,叶元,萧培珍,等.胡子鲶组织器官中4种微量元素的测定分析[J].饲料广角,2008(18):36-38.
[12] 王际英,马晶晶,李宝山,等.野生与人工养殖褐牙鲆亲鱼微量元素组成的差异比较[J].渔业科学进展,2012,33(2):29-34.
[13] 李明德,马锦秋,吴跃英,等.白洋淀鱼类的元素[J].水生生物学报,1996,20(1):86-90.
[14] Joviˇcic'K,Nikolic'D M,Višnjic'-Jeftic'Ž,et al.Mapping differential elemental accumulation in fish tissues:assessment of metal and trace element concentrations in wels catfish(Silurus glanis) from the Danube River by ICP-MS[J].Environmental Science and Pollution Research,2015,22(5):3820-3827.
[15] 史可江,马龙江,刘桂立.南四湖鱼类微量元素含量调查研究[J].肉品卫生,1997(7):13-15.
[16] Abaychi J,Al-Saad H T.Trace elements in fish from the Arabian Gulf and the Shatt al-Arab river,Iraq[J].Bulletin of Environmental Contamination and Toxicology,2008,40(6):226-232.
[17] 贾建萍,周彦钢,林赛君,等.金枪鱼骨营养成分分析[J].食品工业科技,2013,10(34):334-337.
[18] 罗红宇,王斌,冯刚,等.用ICP-MS法分析3种魟鱼和2种鲨鱼软骨中矿物元素含量[J].营养学报,2011,3(5):529-530.
[19] 雒林通,万红玲,杨立,等.天水湖草鱼体内微量元素含量的测定及相关性分析[J].光谱实验室,2013,30(4):1796-1800.
[20] 甄润英,陶秉春,马俪珍,等.3种鲶鱼肌肉主要营养成分的对比分析[J].食品与机械,2008,24(4):108-110.
[21] 樊晓丽,林植华,丁先龙,等.鲶鱼和胡子鲶的两性异形与雌性个体生育力[J].生态学报,2014,34(3):555-563.
[22] 杨丽华.重金属(镉、铜、锌和铬)对鲫鱼的生物毒性研究[D].广州:华南师范大学,2003.
[23] 程宝平,王宝良,高学兴.用流网充分开发颌针鱼资源[J].海洋渔业,1982,5(3):218-219.
[24] 胡星星,王丛丛,产久林,等.南极独角雪冰鱼(Chionodraco hamatus)miR-7132对红细胞发生的作用研究[J].海洋与湖沼,2016,47(3):587-592.
[25] 李博雅,房栋栋,闾坚强,等.乳酸与骨骼肌运动性疲劳关系的研究进展[J].医学综述,2016,22(4):640-642.
[26] 任衍钢.钠钾泵是怎样发现的[J].生物学通报,2011,46(3):60-62.
[27] 雷志洪,徐小清,惠嘉玉,等.鱼体微量元素的生态化学特征研究[J].水生生物学报,1994,18(4):309-315.
[28] 朱晓霞,汪国英,朱贤英.钙的作用机理及如何安全补钙的研究现状[J].保健医学研究与实践,2014,11(1):91-94.
[29] Shih A J,Tai B L,Zhang L H,et al.Prediction of bone grinding temperature in skull base neurosurgery[J].CIRP Annals-Manufacturing Technology,2012,61(1):307-310.
[30] 魏志静,刘希玉,张玉军.儿童血液必需元素与血红蛋白浓度的相关分析[J].西华大学学报,2006,25(1):60-62.
[31] 颜晓平,徐建雄.钒的生物学作用研究进展[J].上海交通大学学报:农业科学版,2003,21(3):254-256.
[32] 郭玉华,赵健亚,周为琴.钒的生物学功能及其最新研究进展[J].家畜生态学报,2004,25(2):47-49.
[33] 秦林林,包安德,马海波,等.微量元素锌、铜、锰的摄入对生长期大鼠骨骼的影响研究[J].微量元素与健康研究,1999, 16(2):7-9.
[34] 顾云,王盛宇,黄益民,等.硒对心脏移植大鼠外周血单个核细胞免疫分子、基因表达的影响[J].中国药学杂志,2006,41 (24):1866-1871.
[35] 贾茜茜,姜宇强,袁刚,等.浙江大洋水库沉积物重金属、营养盐生态风险评价[J].大连海洋大学学报,2016,31(4):410-415.
Analysis of elements in skeleton of Antarctic icefishes Chionodraco hamatus and Trematomus bernacchii
SUN Jian-fei1,LIU Yang1,2,XU Qiang-hua1,2,3,4
(1.College of Marine Sciences,Shanghai Ocean University,Shanghai 201306,China;2.Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources,Ministry of Education,Shanghai 201306,China;3.National Distant-water Fisheries Engineering Research Center,Shanghai 201306, China;4.Collaborative Innovation Center for Distant-water Fisheries,Shanghai 201306,China)
The contents of macroelements(Na,Mg,Al,K,Ca,56 Fe and Zn)and microelements(V,Cr,Mn, As,Se and 206Pb)were detected in Antarctic icefishes Chionodraco hamatus(CH)and Trematomus bernacchii (TB)using inductively coupled plasma mass spectrometry(ICP-MS)method in order to compare the element concentrations of hemoglobinless between Antarctic icefish CH and TB.Results showed that there were significant differences in concentrations of Al,K,56Fe,Se and Mn in the vertebrae and significant differences in concentrations of Mg,K,Ca,56Fe,V and Mn in the skull between CH and TB(P<0.05),which might be attributed to the functional red blood cells shortage in the Antarctic icefish.The comparison of element concentrations in vertebrae between the two Antarctic icefishes and seven non-Antarctic marine fishes and three freshwater fishes revealed that there were much lower levels of macroelements K,Na,Mg,56Fe,Zn,and Ca in skeletons of the two Antarctic icefishes,significantly lower levels of microelements Mn,V,Cr and 206Pb in the two Antarctic fishes than those in the non-Antarctic fishes,which is involved in much lower contamination in the Antarctic marine environment.The first insight into element concentration of the two Antarctic fishes will provide some scientific basis for the physiological difference comparison in Antarctic icefishes through tissue element analysis.
Chionodraco hamatus;Trematomus bernacchii;macroelement;microelement;skeleton
Q581
A
10.16535/j.cnki.dlhyxb.2017.04.002
2095-1388(2017)04-0387-06
2016-11-22
国家自然科学基金面上项目(31572598);上海市教育发展基金会和上海市教育委员会曙光计划项目(13SG51);上海市浦江人才计划项目(16PJ1404000);上海市教委水产学高峰学科项目
孙建飞(1991—),男,硕士研究生。E-mail:1210357105@qq.com
许强华(1974—),女,博士,教授,博士生导师。E-mail:qhxu@shou.edu.cn
