音乐节奏与动作节奏的序列匹配认知机制研究①
2017-06-28王东雪河南师范大学音乐舞蹈学院河南新乡453007
王东雪 (河南师范大学 音乐舞蹈学院,河南 新乡 453007)
音乐节奏与动作节奏的序列匹配认知机制研究①
王东雪 (河南师范大学 音乐舞蹈学院,河南 新乡 453007)
演奏家是通过听觉对音乐感知,以动作节奏来诠释音乐内涵与演奏风格,动作节奏不仅展现出了肌肉收缩的动态变化,而且在视觉空间中与听觉联合进而展现身体动静结合的姿态。人体这种在身体生理性与心理神经性的复杂关系,足以说明动作节奏与音乐节奏互为同步,而且演奏时,动作节奏中的动作序列是按照音乐节奏时序而逐一展现出来的,也就是说二者是以“序列匹配”为认知基础的,本研究试图从脑认知神经科学的视角出发,对音乐节奏与动作节奏的这种序列匹配关系进行深入剖析。
音乐节奏时序;动作节奏序列;序列匹配认知
引 言
钢琴、小提琴等乐器演奏家在进行表演时,他们正是凭借对音乐节奏的认知:“通过中枢神经系统的兴奋扩散效应,诱发动觉中枢的兴奋,从而产生节奏感,即听觉和动觉的联合知觉”[1],以动作节奏来“回应”音乐节奏。这两种不同类型、不同性质的节奏是互为同步的:因为音乐节奏是以一定时间间隔先后顺序出现的,具有时序性,所以,动作组合中那些既定顺序排列的动作——动作序列(如:钢琴家手指触键顺序)与音乐节奏时序是相互匹配的。也就是说,音乐节奏与动作节奏之间存在着序列匹配认知关系。本研究拟从音乐节奏序列认知和动作节奏时序认知入手,来阐述二者的序列匹配认知机制。
一、关于序列匹配认知
(一)音乐节奏时序
从作曲理论角度来看,音乐节奏有着如下界定:“节奏这个概念在音乐理论中具有不同层次的意义。狭义地说,它仅作为与‘旋律’(而不是‘曲调’)相对应的一个概念,单纯只指音乐中的时间因素。……再扩大一些来看,节奏还涉及音乐作品时间段(即‘句法’或‘曲式结构’)的划分、比例与平衡问题(彭志敏,1997)”[2],“节奏是循环的拍子和重音,规律的或是零散的。节奏还与速度的概念不同。节奏是时间中的比例和顺序,但又不是时间本身(姚恒路,2003)”[3];“在音乐中,音的长短、强弱有组织地进行叫做节奏。节奏是音乐在时间上的组织(晏成佺,童忠良,2006)”[4]。从上述界定来看,至少有一点认识是较为统一的:即音乐节奏与时间相关,而“音乐节奏本质上就是音符时值序列(时间间隔)”[5]。所以,将节奏中音符时值的顺序排列称之为音乐节奏时序。从以上分析来看,音乐节奏序列具有以下特征:(1)听觉时序性。因为音乐节奏的产生是通过听觉信息传导:即听觉系统通过外耳接收声波、辨别声源的,由内耳使感音器官——耳蜗的基底膜震动,从而刺激基底膜上的毛细胞,产生神经冲动,“这些神经冲动以特定的频率和组合形式编码声音信息”[6],“在听觉通道,初级听皮层中的神经细胞对于声音频率是有选择性的,两个不同频率的音调能被整合为一个按照出现顺序而具有不同频谱分布的知觉事件”[7],又以音符时值序列作为时间间隔标准。(2)强、弱性。在音乐中,节奏和节拍总是同时存在的,相同时值的强拍和弱拍会进行有规律的循环出现,这样乐器演奏家就可以随着强、弱节拍展现出一定视觉意义上的动作节奏表征:力度大小。(3)速度性。音乐进行的快慢通常是根据乐曲的风格、内容而定,每首作品都有自己确切的速度。如在谱中有“=108”的标记,指的是使用节拍器标示速度,即速率,意思是每分钟演奏108个四分音符。另外,目前国际上通用的意大利文速度术语对音乐作品的速度有进行明确的规定,一般在中国科协制造的节拍器就有像行板(Andante)为每分钟拍数66、中板(Moderato)每分钟拍数88、广板(Largo)每分钟拍数46、快板(Allegro)每分钟拍数132、急板(Presto)每分钟拍数189、最急板(Prestissimo)每分钟拍数210等的速率界定[8]。这充分表明:乐器演奏家只有准确地“听出”音乐速度才能产生出正确的动作速度(如:动作的快、慢)。
(二)动作节奏序列
在运动训练学理论中,动作节奏是指:在完成动作过程中的时间特征。包括用力的大小、时间间隔的长短、动作幅度的大小及动作快慢等因素[9]。《体育科学词典》(2000)对动作节奏的衍生概念——“节奏感”进行定义:训练学上的节奏感(sense of rhythm)指运动员练习过程中,在完成动作的时间和力度上呈现出来的快慢、强弱有序变化的能力;心理学上的节奏感(rhythmic perception)则指个体对运动表象或自身运动的时间与空间动态特征的知觉[10]。无论是从何种角度出发,毋庸置疑,在运动过程中,动作节奏实质上是以动作组合中按照既定顺序排列的动作——即动作序列为核心的,所以,本研究将动作节奏中的动作序列定义为“动作节奏序列”。动作节奏序列有以下特征:(1)时间性。动作的完成是一个从开始到结束的完整过程,这一过程本身就是以节奏的时间间隔为特征的。(2)顺序性。从其定义上来看,成套动作中单个动作之间是以既定先后顺序连接而成的,以保证动作的连续性。(3)动态性。动作序列表现出的是各种不同动作的空间性与时间性,是一种从生理——心理到肌肉输出动作的过程。
(三)序列匹配认知的界定
音乐节奏与动作节奏保持同步,是需要接受两类感觉信息进行“意识性”的动作转化:“第一类是来自听觉、视觉的信息,但有研究证实:与视觉节奏相比,节奏性运动更倾向于受到听觉节奏的影响(Bruno H. Repp,Amandine Penel,2004)”[11],而后大脑听觉中枢将“声音信息”(听觉信息)传导至中枢神经系统,并对“声音信息”——音乐节奏进行认知;第二类感觉信息:即“来自肌肉、关节和前庭器官的传入冲动,提供关于肌肉长度、张力、关节位置、身体的空间位置等信息”[12],简而言之,动作节奏序列与音乐节奏时序是一一对应匹配的。本研究将这一现象的心理活动过程称之为:序列匹配认知。
二、音乐节奏时序认知机制
音乐节奏时序认知,是指在既定节拍下,将不同时值长短的音乐节奏按照时间先后顺序进行排列的心理过程。音乐节奏时序认知机制如下:(1)与时间间隔有关的脑区:神经动力学通过脑磁图测量(MEG)研究,发现初级听觉皮层与节奏间隔变化相关[13];右侧DLPFC参与时间间隔的估计[14];当节奏以固定时间间隔出现特定重音(给人的感觉是一个简单的节拍)时,基底神经节、pre-SMA/SMA和前上颞脑回表现出较大的活跃[15]。(2)与时机相关的脑区:小脑在时间加工中具有重要的作用,比如前馈和纠错计算,以及感觉——运动的整合[16];基底神经节也具有时机作用[17]。时机作用的发生,还与脑对时间顺序的控制有关,时间控制主要涉及前辅助运动区、额下回、中央前沟、颞脑回,而且辅助运动区(SMA)、颞上回、下顶叶皮质这组区域对于抽象概念、运动独立、时序控制来说是重要的[18],后顶叶皮层参与了时间信息加工[19]。(3)与时间序列有关的脑区:前运动皮质内侧(Schubotz和von Cramon,2001b)和外侧(Schubotz和von Cramon,2001a;Schubotz等,2003)与时间序列相关;而辅助运动区(SMA)的“恰当时机”和背侧前运动皮层(PMd)的顺序序列有关[20];颞上回(STG)包括皮层在颞-顶叶的结点、次皮质、基底神经节和小脑,这些区域对时间序列性能扮演不同角色[21],而且在音乐背景下,基底神经节、dPMC和小脑涉及到了更复杂的序列活动[22];小脑后叶主要与非节奏性的时间模式有关,而在组块形成后,前叶有助于组织节奏模式[23];前额皮质(PFC)在时序知觉中的作用可能与注意和工作记忆对时间信息的获得、维持和组织等功能有关[24]。
三、动作节奏序列认知机制
前文提到动作节奏的核心就是动作序列,所以,这里主要针对动作序列展开讨论。美国学者拉什利较早地提出了运动顺序原理,认为运动是以一连串动作的方式进行的,一个运动顺序将要完成时,另一个顺序即准备启动[25],所以,动作……大多数都是具有完整序列的运动(Bernstein,1996)[26]。像乐器演奏家在学习动作技能时:“新的运动可以被分割成块(分为组块),按照一定的节奏进行学习。渐渐地,组块被组合以形成整个动作序列”[27],经过长期练习后,将动作序列以程序性记忆(是指对动作和认知技能的学习[28])的形式记下来,形成“运动程序”储存在大脑里。这样一来,动作技能达到高度自动化,加上动作序列是“依赖于中枢模式发生器(central pattern generators)——脊髓内产生运动,输出一定节律模式的神经机制”[29],使得“动作序列具有节奏性,而且节奏反映出(动作)序列被组块分为短序列的运动(节奏有利于动作技能的学习,是通过连续的动作组块学习而实现自动化)”[30]。由此推之,动作节奏序列认知是指:通过心理活动(如:感觉与知觉、学习与记忆、注意与意识等等),对一定节奏,按照既定动作序列所进行的信息编码与加工过程。
PET研究显示出在动作序列过程中,辅助运动复合体(SMC,supplementary motor complex)有所激活。前辅助运动区在动作组合或正确顺序的动作链接中也有一定的作用(Parashkev Nachev, Christopher Kennard,Masud Husain,2008)[31],还和SMA主要参与组织或更复杂的、分块的运动序列;前运动皮质参与相对复杂序列的任务,其可能促进了对运动的预测;研究显示,如果是更复杂的序列则需要来自基底神经节、dPMC和小脑的共同活动(Doyon, J等,2003;Penhune, V等,2005;Hikosaka, O.等2002;Thach, W. T,2005;Sakai, K.等,2004;Kennerley, S. W.,2004;Janata, P.等,2003;Schubotz, R.I.等;2003;Bengtsson, S. L.等,2004)[16],而小脑对于序列学习和个人动作序列整合统一来说是重要的(Doyon, J., Penhune, V. B.等,2003;Penhune, V. B. & Doyon, J.等,2005;Hikosaka, O., Nakamura等,2002;Thach, W. T.,1998;Garraux, G.等,2005)[16],动作节奏序列认知机制具体如下(如图1所示):(1)前额叶皮质和辅助运动皮质在计划和按特定顺序组织一个运动的快速序列中是重要的[32];(2)当人们在视觉、听觉或感觉运动的反馈线索指导下完成动作时,运动前区(前运动皮质)激活[33],又和辅助运动皮质负责动作序列,其中实现快速肢体动作和精细灵巧的任务,启动和终止运动与小脑半球外侧有关,皮质小脑促成运动准备(前馈和预期)活动;(3)由脊髓、小脑对运动执行或(反馈)调整,其中间区域复制运动程序,通过运动皮质(负责动作执行)发送到肌肉[34],输出动作序列。
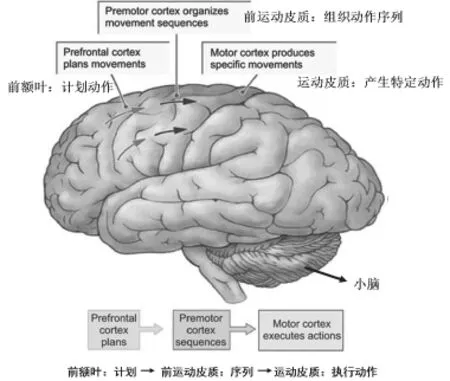
图1.动作序列和计划的皮质基础与机制[34]
乐器演奏最大的特点是音乐与身体运动(抬指、触键、运弓、滚揉等)互为融合。所以,运动程序中除了有对动作序列学习记忆外,还应包括音乐的记忆,即当行动者预计即将到来的音调时,尤其是当旋律曲调与运动转变保持一致时,……听觉反馈的预计导致了较强的联合知觉以控制运动时间和运动(cf.Wolpert,Miall与kawato,1998)[35],一旦听觉接收到“与序列中特定的动作相对应的关键音乐元素”[36],与序列动作对应的音乐就成为一种外在刺激,运动皮质就开始“按图索骥”,对应的运动程序启动,动作序列就会逐一展现出来。尽管动作节奏本身就具有时间特征,但这种“时间特征”已转移到“音乐节奏”上:学习新的动作时,“新的动作被分成组块,按照一定的节奏进行学习。然后,将组块组合以形成完整动作序列。所以,音乐节奏就以时间节律的形式来‘引导’动作序列的发生(Sakai,2004)”[37],这使得动作序列必须在音乐节奏的“约束”下完成,乐器演奏家需要根据音乐节奏对动作序列进行“时序(时间顺序)”分配。
四、音乐节奏与动作节奏的序列匹配认知机制
(一)音乐节奏与动作节奏的脑区匹配
人脑对音乐节奏产生认知,所涉及到的脑区主要有:(1)左、右脑区:左半球优先激活熟悉的音乐、音程任务和节奏,右半球主要负责音色任务。……节奏任务激活左下BA44/6区,并延伸到邻近的岛叶,意味着这一脑区的任务是对连续的声音进行加工处理(Platel H, Price C等,1997)[38]。对连续听觉节奏(auditory rhythmic sequences)的处理加工涉及到一个特定的左脑半球颞缓冲区(Di Pietro M 等,2004)[39]。(2)额叶、顶叶、颞叶、小脑、丘脑等。额叶皮层、尤其是Broca区对节奏知觉具有一定作用[40],而且左额(BA6)、顶叶皮层和右前小脑负责记忆熟悉、定期的节奏(Katsuyuki Sakai等,1999)[41]。另一方面,右额叶、额(BA6)、顶叶区域,以及双侧后小脑则负责记忆不熟悉的节奏(Lawrence M,2001)[42]。再有,音色特征的处理与小脑的认知区域被激活有关(Alluri V,Toiviainen P等,2012)[43]。旋律方面,颞上回对旋律的处理起着重要的作用(Liégeois-Chauvel C, Peretz I等,1998)[44],可能因为前颞部分参与和弦的认知(Satoh M,Takeda K 等,2007)[45]。双侧额叶脑回、丘脑内侧、和背扣带皮层能被主旋律和次旋律调动起来(Mizuno和Sugishita ,2007)[46]。(3)基底神经节对于音乐节奏的认知作用是非常重要的。其主要作用是在大脑内部产生节拍,而且,视觉节奏也可以激活内部听觉节奏(Grahn JA,Henry MJ, McAuley JD,2010)[47]。同时,对一系列短暂音程的跳跃、或节拍的感知,也与基底神经节活动有关(Grahn JA, Rowe JB,2012)[48]。
当然,乐器演奏家之所以能够伴随音乐节奏进行动作技能的表演,不仅仅是因为脑能够对音乐节奏产生认知(见前文部分),而且其脑运动皮质也同时对音乐节奏有所认知:特别是在外侧运动前区、辅助运动区和躯体运动区(Popescu M, Otsuka A 等,2004)[49]、双侧额下回、颞上回和前运动皮层在内的双侧皮质网络(Minati L 等,2008)[50]、左腹侧前运动皮层(PMV)的激活与音乐节奏有关(Kornysheva等,2010)[51]。另外,在神经结构存在一个共享网络(双侧颞区、左顶下小叶、右额岛盖区)主要负责节奏认知,且不受音乐背景影响(Limb CJ, Kemeny S等,2006)[52]。基底神经节和SMA已被认为与韵律和脉冲知觉有关,并已被证明当倾听韵律性的节奏时二者表现得更为活跃,而仅仅是听节奏就不可能诱发脉冲认知(Grahn and Brett,2007;Grahn,2009;Grahn and Rowe, 2009)[53]。一项正电子发射断层扫描术(PET)研究(Penhune 等,1998)还证实了外侧小脑和基底神经节具有重现节奏的功能[54]。随后的功能磁共振成像研究发现,在不同水平的音乐训练中,节奏均能引起基底神经节和辅助运动区(SMA)较高的活性……前运动皮层、小脑和SMAs(辅助运动前区和辅助运动区)的激活增加与节奏类型无关,但基底神经节和SMAs可能对节拍知觉起到调解作用(Grahn JA, Brett M等,2007)[55]。Heather L.Chapin等研究者(2010)[56]对神经回路中潜在的节奏和韵律知觉进行调查研究,发现脑区之间的功能是部分重叠的,对节奏序列的产生和有关运动较为敏感(Dhamala 等,2003;Chen等,2006, 2008b;Karabanov等, 2009;Thaut等, 2009)。节奏知觉可以激活相关运动脑区,即使没有明显的动作,在初级运动皮层(PMC)(Schubotz等, 2000; Grahn and Brett, 2007;Chen等,2008a;Bengtsson等,2009;Grahn, 2009;Grahn and Rowe,2009)、小脑(Schubotz等, 2000;Grahn and Brett, 2007;Chen等, 2008a;Bengtsson等,2009)、前辅助运动区(Schubotz等,2000;Grahn and Brett, 2007;Bengtsson等,2009)、辅助运动区(SMA)(Schubotz等,2000;Grahn and Brett,2007;Chen等,2008a;Bengtsson等, 2009;Grahn,2009; Grahn and Rowe,2009)和基底神经节((Schubotz等,2000;Grahn and Brett, 2007;Grahn,2009;Grahn and Rowe,2009)仍然能显示出活动状态。
至此,不难发现音乐认知脑区与运动脑区及其功能是相互叠加、匹配的,从而使音乐节奏与动作节奏认知同步。各脑区作用总结如下(见图2):
(1)辅助运动区:计划运动与编码;对音乐节奏产生认知。(2)额叶皮质:主要根据各种信号(光、声音等)输出计划运动;具有节奏知觉,能够记忆音乐节奏。(3)前运动皮质:与辅助运动皮质共同参与运动计划与编码等;能够对音乐节奏产生偏好。(4)顶叶皮质:感知空间(躯体、视听觉)知觉,对音乐节奏产生认知。(5)基底神经节:控制动作力度,负责动作执行等;产生或参与内部节拍,调节节拍知觉;(6)小脑:对运动计时,协调与执行动作等;能够记忆与识别音乐节奏。(7)脑干—脊髓系统:负责动作执行;识别音乐小调。
(二)音乐节奏与动作节奏的序列匹配认知机制
前文分别对音乐节奏序列与动作节奏序列的认知机制进行了解释,那么二者之间的序列匹配认知机制是如何发生的呢?见图3所示:
1.听觉与记忆是互为交互的。一旦听觉接收到音乐节奏,凡是能够对音乐节奏产生认知的相关运动脑区(诸如:辅助运动区、前运动皮质、额叶皮质、顶叶皮质、基底神经节、小脑、脑干—脊髓等)就会被激活,继而触发额叶皮质做出运动计划,SMA前部也参与了运动计划和运动决策(Marsden等,1996),特别是pre-SMA负责提前运动准备,指导内部线索(Deiber等,1991)[57]。额叶皮质产生节奏知觉的同时,开始启动程序性记忆,恢复对接收音乐节奏的记忆,搜寻与之对应的运动程序。
2.与此同时,前运动皮质准备运动程序,前额皮质通过小脑与听觉来控制时间,组织动作序列,并对动作序列进行时间分配,在该过程中,额叶皮质中的内侧运动区和基底神经节在多个动作的时间序列扮演了特别重要的角色,还有辅助和前辅助运动区领域参与了构建多个动作序列执行的时间结构[58]。动作(前额叶皮层)意图的产生依靠pre-SMA,它又与基底节连接启动行动,并确定动作序列的时间结构,即pre-SMA主要对即将发生的动作行为进行时间结构策划(Mita等,2009),小脑可能参与了对动作的时间微调[59]。

图3.音乐节奏与动作节奏的认知序列匹配机制图②图3改编自Arnold L. Glass. http://www.rci.rutgers.edu/~aglass/02_MotorSystem.ppt[EB/OL]和Sonja A. E. Kotz,Michael Schwartze. Differential input of the supplementary motor area to a dedicated temporal processing network: functional and clinical implications[J].Front Integr Neurosci,2011,5:86.
3.音乐节奏序列与动作节奏序列的整合。在伴随音乐进行的动作中,听觉对于音乐节奏的辨识非常重要,因为动作与音乐被作为一个整体加以记忆。也就是说,最初学习动作时,动作“序列被组成若干个短序列的组块[60]”,而音乐节奏根据时间间隔也被划分成与动作“短序列组块”相对应的组块,由于运动技能学习是一种定时模式(Sakai K.等,2004)[61]:小脑后叶和前额叶皮层被用于节奏的形成,这一加工过程需要联合(认知)电路和关注,而节奏一经形成,由前小脑、运动前区组成运动路线就会变得活跃[62],所以,演奏家根据音乐节奏来完成各种动作,“每一个动作的产生能有相对应的声音(Zatorre等,2007)”[63],而且只有“听觉才能够影响到随后的一个个动作(Robert J. Zatorre等,2007)”[16]。
4.音乐节奏时序认知的最终信息与负责组织动作序列的关键脑区——前运动皮质“会合”,而此时,节奏感知和产生的区域——小脑和基底神经节(Ivry,Keele 1989;Janata,Grafton,2003)以及多个领域,如前运动皮层和辅助运动区(Halsband等,1993)开始活跃,定时、同步和输送可能会由小脑中的分层控制振荡器系统负责(Ivry 和Hazeltine 1995;Ivry 和Schlerf 2008;Sternberg 等,1982),促进节奏感的产生(Levitin和Cook,1996)[64],初级运动区与初级躯体感觉皮质共同协调,由脑干、脊髓控制,小脑和基底神经节执行与控制相关肌肉的收缩以产生动作节奏。
结 论
乐器演奏家是比较特殊的,特殊之处就是在于他(她)们需要在音乐节奏中完成动作。在完成动作过程中,动作节奏通过音乐节奏来准确展现动作的表征,即:肌肉收缩的动态变化——动作力度、快慢、动静,乐器演奏家以动作节奏来诠释音乐的内涵以及表现个人技术风格,并与音乐节奏交相呼应。研究发现:音乐节奏与动作节奏的认知脑区及其功能是重叠的,足以说明二者认知是同步进行的。既然二者同步,而且动作节奏序列是以动作序列为核心的,音乐节奏时序又是以音符时值序列(时间间隔)为标准,那么二者的认知应该是建立在“序列”匹配基础之上的,也就是说音乐节奏与动作节奏之间存在序列匹配认知的关系:(1)听觉与记忆是互为交互的。一旦听觉接收到音乐节奏,凡是能够对音乐节奏产生认知的相关运动脑区(诸如:辅助运动区、前运动皮质、额叶皮质、顶叶皮质、基底神经节、小脑、脑干—脊髓等)就会被激活,听觉能够启动程序性记忆中与音乐节奏相应的运动程序。(2)前额叶负责计划动作,前运动皮质组织动作序列,由运动皮质执行动作的输出,而此过程中的动作节奏序列会按照音乐节奏中音符时值的顺序——音乐节奏时序排列而逐一展现出来。(3)动作节奏序列与音乐节奏序列互为整合。因为动作节奏序列在学习初,是以“序列组块”加以记忆的,并与音乐节奏中音符时值的顺序排列——音乐节奏时序的划分相匹配。(4)音乐节奏时序认知的最终信息与负责组织动作序列的关键脑区——前运动皮质“会合”,产生动作节奏输出。
[1] 张力为,毛志雄.运动心理学[M].上海:华东师范大学出版社,2003:145.
[2] 彭志敏.音乐分析基础教程[M].北京:人民音乐出版社,1997:99.
[3] 姚恒路.现代音乐分析方法教程[M].长沙:湖南文艺出版社,2003:20.
[4] 晏成佺,童忠良.基本乐理教程(第2版)[M].北京:人民音乐出版社,2006:13.
[5] Daniel J. Levitin,Parag Chordia,Vinod Menonc.Musical rhythm spectra from Bach to Joplin obey a 1/f power law[EB/OL].www.pnas.org/cgi/ doi/10.1073/pnas.1113828109.
[6] 王庭槐.生理学(第2版)[M].北京:高等教育出版社,2008:247.
[7] 尹天子,陈有国,黄希庭.时序知觉的认知与神经机制[J].西南大学学报(社会科学版)[J].2010(5):28-32.
[8] 晏成佺,童忠良.基本乐理教程(第2版)[M].北京:人民音乐出版社,2006:171.
[9] 体育学院通用教材.运动训练学[M].北京:人民体育出版社,2000.
[10] 中国体育科学学会,香港体育学院.体育科学词典[M].北京:高等教育出版社,2000:147.
[11] Bruno H. Repp,Amandine Penel.Rhythmic movement is attracted more strongly to auditory than to visual rhythms[J].Psychological Research, Vol.68,Aug 2004:252-270.
[12] 张君邦.神经科学教程[M].北京:科学出版社,2005:136.
[13] Thaut MH.Neural basis of rhythmic timing networks in the human brain[J].Ann N Y Acad Sci,Vol.999,No.1,Nov 2003:364-373.
[14] Jason R. Tregellas,Deana B. Davalos,Donald C. Rojas.Effect of task difficulty on the functional anatomy of temporal processing[J].NeuroImage, Vol.32,No.1,2006:307-315.
[15] Daniel J. Levitin, Anna K. Tirovolas.Current Advances in the Cognitive Neuroscience of Music[J].Annals of the New York Academy of Sciences, Vol.1156,No.1,2009:211-231.
[16] Robert J. Zatorre, Joyce L. Chen,Virginia B. Penhune.When the brain plays music: auditory-motor interactions in music perception and production[J].NEUROSCIENCE, Vol.8,No.7,2007:547-558.
[17] Deborah L. Harrington,Gabriel N. Castillo,et.al.Neurobehavioral Mechanisms of Temporal Processing Deficits in Parkinson’s Disease[J].PLoS ONE, www.plosone. Vol.6,No.2,2011:e17461.
[18] Ullén F, Bengtsson SL, Ehrsson HH, Forssberg H. Neural control of rhythmic sequences[J].Ann N Y Acad Sci, Vol.1060,No.1,2005:368-376.[19] Azanon E, Longo MR, Soto-Faraco S, Haggard P .The posterior parietal cortex remaps touch into external space[J]. Curr Biol, Vol.20,No.14,2010,:1304-1309.
[20] Jill X. O’Reilly, Katharine J. McCarthy, Mariagrazia Capizzi,Anna Christina Nobre.Acquisition of the Temporal and Ordinal Structure of Movement Sequences in Incidental Learning[J].J Neurophysiol, Vol.99,No.5,2008:2731-2735.
[21] Anke Karabanov,Örjan Blom,Lea Forsman, Fredrik Ullén.The dorsal auditory pathway is involved in performance of both visual and auditory rhythms[J]. NeuroImage, Vol.44,No.2,2009:480-488.
[22] Zatorre, R.J., Chen, J.L., Penhune, V.B. When the brain plays music: auditory-motor interactions in music perception and production[J].Nat.Rev. Neurosci, Vol.8,No.7,2007:547-558.
[23] Katsuyuki Sakai, Okihide Hikosaka,Kae Nakamura. Emergence of rhythm during motor learning[J]. TRENDS in Cognitive Sciences, Vol.8,No.12,2004:547-553.
[24] 尹天子,陈有国,黄希庭.时序知觉的认知与神经机制[J].西南大学学报(社会科学版)[J].2010(5):28-32.
[25] 梅锦荣.神经心理学[M].北京:中国人民出版社,2011:290.
[26] Scott T.Grafton,Antonia F. de C. Hamilton. Evidence for a distributed hierarchy of action representation in the brain[J].Hum Mov Sci, Vol.26,No.4,2007:590-616.
[27] Andreas R. Luft,Manuel M. Buitrago. Stages of Motor Skill Learning[J]. Molecular Neurobiology, Vol.32,No.3,2005: 205-216.
[28] M·W·艾森克,M·T·基恩.认知心理学(第五版)[M].上海:华东师范大学出版社,2009:283.
[29](美)詹姆斯·卡拉.生物心理学[M].北京:人民邮电出版社,2011:246.
[30] Katsuyuki Sakai, Okihide Hikosaka,Kae Nakamura. Emergence of rhythm during motor learning[J]. TRENDS in Cognitive Sciences, Vol.8,No.12,2004:547-553.
[31] Parashkev Nachev, Christopher Kennard,Masud Husain.Functional role of the supplementary and pre-supplementary motor areas[J].Nature Reviews Neurcience, Vol.9,No.11,2008:856-869.
[32](美)詹姆斯·卡拉特.生物心理学[M].北京:人民邮电出版社,2011:251.
[33](美)Michael S.Gazzzniga,Richard B.Ivry,George R.Mangun.认知神经科学——关于心智的生物学[M].北京:中国轻工业出版社,2011:246.
[34] Gerry Leisman,Robert Melillo.Effects of motor sequence training on attentional performance in ADHD children[J].Int J Disabil Hum, Vol.9,No.4,2010:1-8.
[35] Peter E. Keller,Simone Dalla Bella,Iring Koch.Auditory Imagery Shapes Movement Timing and Kinematics:Evidence From a Musical Task[J].Journal of Experimental Psychology: Human Perception and Performance, Vol.36,No.2,2010:508-513.
[36] Greg payne,葛培新,梁国立.人类动作发展概论[M].北京:人民教育出版社,2008:405.
[37] Sakai K., Hikosaka O., Nakamura K. Emergence of rhythm during motor learning[J].Trends Cogn Sci,2004,8:547-553.转引自Andreas R.Luft,Manuel M. Buitrago. Stages of Motor Skill Learning[J]. Molecular Neurobiology, Vol.32,No.3,2005: 205-216.
[38] Platel H, Price C, Baron JC, Wise R, Lambert J, Frackowiak RS, Lechevalier B, Eustache F.The structural components of music perception. A functional anatomical study[J].Brain, Vol.120,No.2,1997:229-243.
[39] Di Pietro M, Laganaro M, Leemann B, Schnider A.Receptive amusia: temporal auditory processing deficit in a professional musician following a left temporo-parietal lesion[J]. Neuropsychologia.Vol.42,No.7,2004:868-877.
[40] Lechevalier B.Perception of musical sounds: contributions of positron emission tomography[J].Bull Acad Natl Med, Vol.181,No.6,1997:1191-1199.
[41] Katsuyuki Sakai,Okihide Hikosaka,Satoru Miyauchi,Ryousuke Takino,Tomoe Tamada,Nobue Kobayashi Iwata,Mathew Nielsen. Neural representation of a rhythm depends on its interval ratio[J]. The Journal of Neuroscience, Vol.19,No.22,1999:10074-10081.
[42] LAWRENCE M. PARSONS.Exploring the Functional Neuroanatomy of Music Performance, Perception, and Comprehension[J]. Annals of the New York Academy of Sciences Volume 930, THE BIOLOGICAL FOUNDATIONS OF MUSIC, Vol.930,No.1,2001:211-231.
[43] Alluri V,Toiviainen P,Jääskeläinen IP,Glerean E, Sams M, Brattico E.Large-scale brain networks emerge from dynamic processing of musicaltimbre, key and rhythm[J].Neuroimage, Vol.59,No.4,2012:3677-3689.
[44] Liégeois-Chauvel C, Peretz I, Babaï M, Laguitton V, Chauvel P.Contribution of different cortical areas in the temporal lobes to music processing[J].Brain, Vol.121,No.10,1998:1853-1867.
[45] Satoh M,Takeda K, Kuzuhara S.Music therapy and neuropsychology: a proposal to music therapy based on the cognitive processing of music[J]. Rinsho Shinkeigaku, Vol.47,No.11,2007:868-870.
[46] Daniel J. Levitin, Anna K. Tirovolas.Current Advances in the Cognitive Neuroscience of Music[J].Annals of the New York Academy of Sciences, Vol.1156,No.1,2009:211-231.
[47] Grahn JA,Henry MJ, McAuley JD.FMRI investigation of cross-modal interactions in beat perception: audition primes vision, but not vice versa[J].Neuroimage. Vol.54,No.2,2011:1231-1243.
[48] Grahn JA, Rowe JB. Finding and Feeling the Musical Beat: Striatal Dissociations between Detection and Prediction of Regularity[J].Cereb Cortex, Vol.23,No.4, 2013:913-921.
[49] Popescu M, Otsuka A, Ioannides AA.Dynamics of brain activity in motor and frontal cortical areas during music listening: a magnetoencephalographic study[J].Neuroimage. Vol.21,No.4,2004:1622-1638.
[50] Minati L, Rosazza C, D’Incerti L, Pietrocini E, Valentini L, Scaioli V, Loveday C, Bruzzone MG.FMRI/ERP of musical syntax: comparison of melodies and unstructured note sequences[J]. Neuroreport. Vol.19,No.14,2008:1381-1385.
[51] Kornysheva K, von Anshelm-Schiffer AM, Schubotz RI.Inhibitory stimulation of the ventral premotor cortex temporarily interferes with musical beat rate preference[J]. Hum Brain Mapp, Vol.32,No.8,2011:1300-1310.
[52] Limb CJ, Kemeny S, Ortigoza EB, Rouhani S, Braun AR.Left hemispheric lateralization of brain activity during passive rhythm perception in musicians[J].Anat Rec A Discov Mol Cell Evol Biol. Vol.288,No.4,2006:382-389.
[53] Heather L.Chapin,Theodore Zanto,Kelly J.Jantzen,Scott J.A.Kelso,Fred Steinberg,Edward W.Large.Neural responses to complex auditory rhythms: the role of attending [J].Frontiers in Psychology, Vol.1,No.4,2010:224.
[54] Lauren Stewart, Katharina von Kriegstein, ason D. Warren4 and Timothy D. Griffiths.Music and the brain:disorders of musical listening[J].Brain, Vol.129,No.5,2006:2533-2553.
[55] Grahn JA, Brett M.Rhythm and beat perception in motor areas of the brain[J].J Cogn Neurosci. Vol.19,No.5,2007:893-906.
[56] Heather L.Chapin,Theodore Zanto,Kelly J.Jantzen,Scott J.A.Kelso,Fred Steinberg,Edward W.Large.Neural responses to complex auditory rhythms: the role of attending[J]. Frontiers in Psychology, Vol.1,No.4,2010:224.
[57] Cornelia Exner,Janka Koschack,Eva Irle.The Differential Role of Premotor Frontal Cortex and Basal Ganglia in Motor Sequence Learning: Evidence From Focal Basal Ganglia Lesions[J].LEARNING & MEMORY, Vol.9,No.6,2002:376-386.
[58] Tanji J.Sequential organization of multiple movements:involvement of cortical motor areas[J]. Annu Rev Neurosci, Vol.24,No.1,2001:631-651.
[59] Sonja A. E. Kotz,Michael Schwartze. Differential input of the supplementary motor area to a dedicated temporal processing network: functional and clinical implications[J].Frontiers in Integrative Neuroscience, Vol.5,No.3,2011:439-430.
[60] Katsuyuki Sakai,Okihide Hikosaka,Kae Nakamura.Emergence of rhythm during motor learning[J]. TRENDS in Cognitive Sciences, Vol.8,No.12,2004:547-553.
[61] 转引自:Sakai K., Hikosaka O., Nakamura K. Emergence of rhythm during motor learning[J]. Trends Cogn. Sci. Vol.8,No.12,2004:547-553.[62] Andreas R. Luft,Manuel M. Buitrago. Stages of Motor Skill Learning[J].Molecular Neurobiology, Vol.32,No.3,2005: 205-216.
[63] Virginia Conde, Eckart Altenmüller,Arno Villringer, Patrick Ragert.Task-Irrelevant Auditory Feedback Facilitates Motor Performance in Musicians[J].Frontiers in Psychology, Vol.3,2012:146.
[64] Daniel J. Levitin, Anna K. Tirovolas.Current Advances in the Cognitive Neuroscience of Music[J].Annals of the New York Academy of Sciences, Vol.1156,No.1,2009:211-231.
(责任编辑:李小戈)
J613.7;J60-05
A
1008-9667(2017)01-0099-08
2015-11-28
王东雪(1980—),女,吉林镇赉人,音乐学博士,河南师范大学音乐舞蹈学院校聘副教授,硕士生导师,研究方向:民族音乐学、音乐教育学。
①本文为河南师范大学博士科研启动费支持课题《中国民族音乐学科研方法应用及现状分析》(项目编号:11169)阶段性研究成果之一。
