鱼腥草素钠及其与红霉素联用对产膜表皮葡萄球菌luxS,agr/RNAⅢ 影响的研究
2017-06-22许甘霏王晶晶吴大强官妍
许甘霏+王晶晶+吴大强+官妍

[摘要] 細菌的群体感应及特异基因表达调控在细菌生物被膜形成中有十分重要的作用。luxS,agr均是表皮葡萄球菌群体感应调控中的关键基因,RNAⅢ是agr系统的效应器分子。为了评估鱼腥草素钠及其与红霉素联用对产膜表皮葡萄球菌转录水平的影响,采用连续稀释法测定鱼腥草素钠、红霉素、万古霉素对表皮葡萄球菌的MIC;荧光定量PCR测定鱼腥草素钠及其与红霉素联用、万古霉素、红霉素等对表皮葡萄球菌作用后不同时间段luxS,agr/RNAⅢ的转录水平。结果表明1/2MIC,1/4MIC鱼腥草素钠,1/2MIC鱼腥草素钠+1/2MIC红霉素,1/4MIC鱼腥草素钠+1/4MIC红霉素,1/8MIC鱼腥草素钠+1/8MIC红霉素等在作用于ATCC 35984伊始,就可迅速上调luxS 的表达,与阴性对照相比,存在显著差异(P<0.05);在作用6,12,48 h后,鱼腥草素钠依然存在对luxS的上调作用。鱼腥草素钠在MIC,1/2MIC浓度时对agr表达均有显著下调作用(P<0.05),1/2MIC鱼腥草素钠与1/2MIC红霉素联用、1/4 MIC鱼腥草素钠与1/4MIC红霉素联用,在作用6,12,24 h等时间段对agr表达的下调作用也很显著(P<0.05)。鱼腥草素钠在MIC浓度时,对RNAⅢ表达亦有显著下调作用(P<0.05),1/2MIC鱼腥草素钠与1/2MIC红霉素联用对RNAⅢ表达的下调效果也很显著(P<0.05)。提示鱼腥草素钠及其与红霉素联用,可以迅速上调表皮葡萄球菌luxS转录水平的表达,在特定浓度下可下调agr/RNAⅢ的表达,具有抑制表皮葡萄球菌生物被膜菌之间的相互黏附聚集、抑制膜内营养和输水通道的形成、阻止膜内细菌脱离、并抑制细菌外毒素形成等作用的可能。
[关键词] 表皮葡萄球菌;鱼腥草素钠;红霉素;群体感应;luxS;agr/RNAⅢ;生物被膜
[Abstract] Quorum sensing of bacteria and its specific gene expression regulation have a very important role in bacterial biofilm formation. LuxS and agr are the key regulatory genes in quorum sensing of Staphylococcus epidermidis,and RNA Ⅲ is the effector molecule of agr system. In order to evaluate the effects of sodium houttuyfonate in combination with erythromycin on the transcription level of S. epidermidis,serial dilution method was used to determine the MIC of sodium houttuyfonate,erythromycin and vancomycin on S. epidermidis,and fluorescent quantitative PCR method was used to detect the transcription levels of luxS,agr/RNAⅢ in different time periods after treatment on S. epidermidis by sodium houttuyfonate in combination with erythromycin,vancomycin,and erythromycin alone. Our results showed that in treatment by 1/2MIC,1/4MIC sodium houttuyfonate,1/2MIC sodium houttuyfonate +1/2MIC erythromycin, 1/4MIC sodium houttuyfonate+1/4MIC erythromycin,and 1/8MIC sodium houttuyfonate+1/8MIC erythromycin for ATCC 35984,they could rapidly up-regulate the expression of luxS of S. epidermidis from the beginning as compared with negative control,with significant differences (P<0.05);furthermore,sodium houttuyfonate can still up-regulate the expression of luxS even after treatment for 6,12 and 48 h. Sodium houttuyfonate in MIC and 1/2MIC concentration can significantly down-regulate the expression of agr (P<0.05);1/2MIC sodium houttuyfonate+1/2MIC erythromycin,1/4MIC sodium houttuyfonate+1/4MIC erythromycin,can also significantly down-regulate the expression of agr in 6 h,12 h and 24 h(P<0.05). Sodium houttuyfonate in MIC,can significantly down-regulate the expression of RNA Ⅲ (P<0.05),and 1/2MIC sodium houttuyfonate+1/2MIC erythromycin can also significantly down-regulate the expression of RNAⅢ(P<0.05). Therefore,our presented results showed that sodium houttuyfonate in combination with erythromycin can rapidly up-regulate the transcription of luxS of S. epidermidis,and can down-regulate the expression of agr/RNA Ⅲ in certain concentrations,and suggested that sodium houttuyfonate in combination of erythromycin could inhibit mutual aggregation between S. epidermidis and biofilm bacteria,inhibit membrane nutrition and formation of water transport channels,prevent separation of bacterial cells in biofilm,and inhibit the formation of bacterial exotoxin of S. epidermidis.
[Key words] Staphylococus epidermidis;sodium houttuyfonate;erythromycin;quorum sensing;luxS;agr/RNAⅢ;biofilm
以表皮葡萄球菌Staphylococus epidermidis(Se)为代表的凝固酶阴性的葡萄球菌是临床导致以生物材料为中心感染的主要致病菌之一,在假体植入、手术缝合部位、心脏瓣膜置换、留置导管、静脉中心导管等医源性感染灶中检出率较高。表皮葡萄球菌能否形成生物被膜(biofilm),是目前用来评价其是否具有致病性的最重要的指标[1]。细菌生物被膜是指细菌黏附于惰性或活性实体表面,被自身分泌的胞外黏质物所包裹,具有高度组织化的多细胞细菌群体结构[2]。在难治性感染病灶中,几乎都存在细菌生物被膜感染源。生物被膜结构受诸多因素影响,有很复杂的调节机制,如基因调控、生长的环境条件等。
群体感应(quorum sensing,QS)是指细菌通过自身产生的自诱导因子,去感知周围同类细菌的密度或多寡并调控基因表达的系统[3]。自Davies等发现[4] 细菌的QS及特异基因表达调控在细菌生物被膜形成中有着十分重要的作用以来,干扰QS系统已成为研究新型抗菌药的新方向,在新的抗感染治疗发展中,是一种有吸引力的靶位点。葡萄球菌具有獨特和完善的QS系统,Xu等[5]通过构建和分析表皮葡萄球菌△luxS突变株,证实了luxS(luciferase,荧光素酶)通过AI-2(autoinducer-2,自诱导物-2)的细胞间信号转导,下调ica操纵子,影响PIA(polysaccharide intercellular adhesion,多糖胞间黏附素)的产生,因而抑制了表皮葡萄球菌生物被膜的形成;agr(accessory gene regulator,附属基因调节子)亦是表皮葡萄球菌QS调控中的关键基因[6],RNAⅢ是agr系统的效应器分子。
由于在感染灶的细菌一旦形成生物被膜,不仅引发难以治疗的感染,还易产生多重耐药。因此,如何有效治疗由生物被膜引起的相关感染,已成为临床抗感染研究的重要课题。从中国传统中药中寻求有效抗生物被膜菌的相关药物,研究中西药联用,以达到药物减量增效、降低毒副作用等措施,目前被认为都是可行的途径。
作者以往的研究[7]已证实,亚抑菌浓度的鱼腥草素钠及其与红霉素联用对表皮葡萄球菌生物被膜形成有显著影响,对表皮葡萄球菌黏附和代谢均有抑制作用,因而选择鱼腥草素钠及其与红霉素联用,在不同时间段观察药物对悬浮状态下的产膜表皮葡萄球菌luxS,agr/RNAⅢ 转录水平的影响,以期从分子水平评估鱼腥草素钠及其与红霉素联用抗表皮葡萄球菌感染的效果,为中药及中西药联用治疗表皮葡萄球菌引起的相关感染,提供研究依据和实践参考。
1 材料
1.1 菌株与培养基
ATCC 35984(表皮葡萄球菌产膜标准菌株,复旦大学上海医学院瞿涤教授馈赠);TSB培养基(胰蛋白胨大豆肉汤培养基,杭州微生物试剂有限公司);MH液体培养基(杭州微生物试剂有限公司)。
1.2 药品与试剂
万古霉素标准品、鱼腥草素钠标准品、红霉素标准品(中国食品药品检定研究院);二甲基亚砜(天津市光复精细化工研究所);SYBR GreenⅠRT-PCR试剂盒(宝生物工程有限公司);SYBR Premix Ex TaqⅡ(Tli RNaseH Plus)(宝生物工程有限公司);其余所用试剂均为国产分析纯。
1.3 仪器
紫外-可见分光光度计(上海龙尼科仪器有限公司);PCR仪(Eppendorf);凝胶成像系统 Tanon-1600(Tanon);核酸电泳系统(Bio-Rad);高速冷冻离心机(日立公司);qRT-PCR仪(QuantStudio TM 6 Flex美国赛默飞世尔):恒温摇床(上海智城分析仪器制造有限公司)。
1.4 引物合成
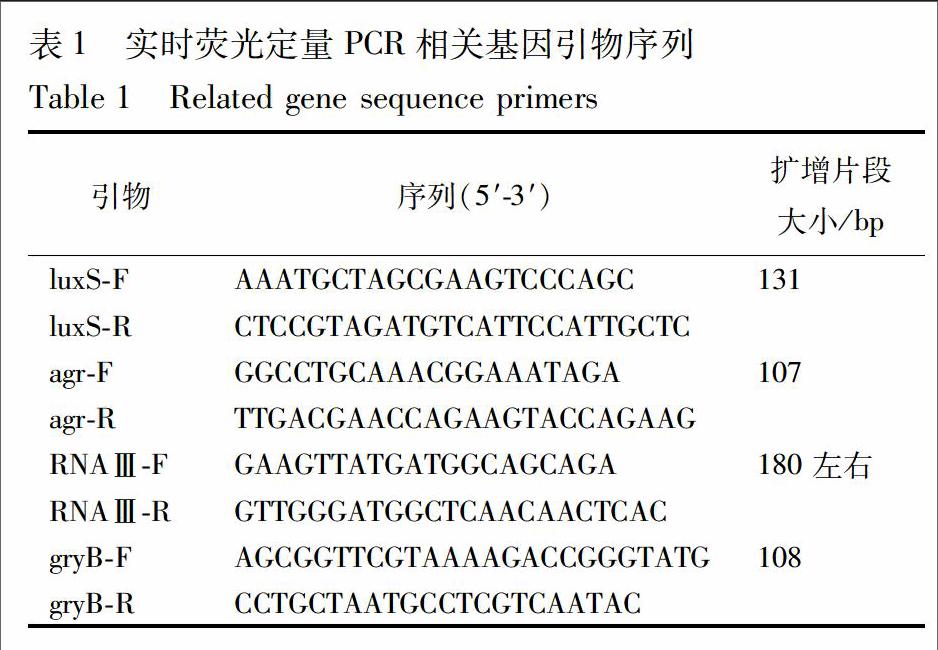
根据NCBI中 ATCC 35984 gryB和luxS,agr,RNAⅢ的序列设计、合成的扩增引物序列见表1。
2 方法
2.1 鱼腥草素钠、万古霉素、红霉素对产膜表皮葡萄球菌MIC的测定
用MH液体培养基,以连续稀释法使每支试管2 mL培养基中药物的终浓度分别为128,64,32,16,8,4,2,1,0.5,0.25 mg·L-1,向各试管中加入200 μL 0.5 Mc菌液;另设空白对照(不含药物和菌液的培养基),阴性对照(只含有菌液而不添加药物的培养基),每组设置2个平行。37 ℃条件下恒温培养24 h,分光光度计法测A600,与空白对照比较得药物的MIC。
2.2 药物对悬浮状态下产膜表皮葡萄球菌的作用
2.2.1 菌细胞制备 取2 mL 0.5 Mc菌液,按1%的接种量接种于200 mL TSB液体培养基中,于37 ℃,150 r·min-1条件下培养;分光光度计测A600达到1.6时,8 000 r·min-1,4 ℃离心30 min,弃上清;分别用新鲜的TSB培养基200 mL重悬细胞。
2.2.2 药物处理 不添加药物(阴性对照),MIC万古霉素(阳性药对照),1/2MIC万古霉素(阳性药对照),MIC红霉素,1/2MIC红霉素,1/4MIC红霉素,MIC鱼腥草素钠,1/2MIC鱼腥草素钠,1/4MIC鱼腥草素钠,1/2MIC鱼腥草素钠+1/2MIC红霉素,1/4MIC鱼腥草素钠+1/4MIC红霉素,1/8MIC鱼腥草素钠+1/8MIC红霉素。
2.2.3 药物处理后菌细胞的保存 阴性对照及加入药物的各组菌液,继续于37 ℃,150 r·min-1培养;在1,6,12,24,48 h分别取样10 mL于10 mL Ep管(DEPC水处理并灭菌)中,12 000 r·min-1,4 ℃离心8 min,弃上清,-70 ℃保存备用。
2.3 表皮葡萄球菌RNA抽提
取冻存于-70 ℃的菌体,加液氮研磨菌体至粉末状;加入350 μL裂解液RL,涡旋振荡30 s,12 000 r·min-1,4 ℃离心5 min;取上清加入250 μL无水乙醇,转入吸附柱CR3中(吸附柱放在收集管中),12 000 r·min-1,4 ℃离心1 min;向吸附柱CR3中加入350 μL去蛋白液RW1,12 000 r·min-1,4 ℃离心1 min,弃废液;再向吸附柱CR3中央加入80 μL DNaseⅠ工作液,放置10 min;加入350 μL去蛋白液RW1,12 000 r·min-1,4 ℃离心1 min;加入500 μL漂洗液RW,放置2 min,12 000 r·min-1,4 ℃离心1 min(重复该步骤1次),弃废液。吸附柱放置5 min,彻底晾干吸附柱材料中残余的漂洗液;将吸附柱CR3转入一个新的RNase-Free离心管中,向吸附膜中央悬空滴加40 μL RNase-Free ddH2O,放置2 min,12 000 r·min-1,4 ℃离心2 min,得到RNA溶液。-70 ℃保存备用。
2.4 目的基因表达的测定
采用荧光定量PCR检测产膜表皮葡萄球菌ATCC 35984 的luxS,agr,RNAⅢ转录水平。
2.4.1 将RNA逆转录为DNA 去除基因组DNA反应:按SYBR GreenⅠRT-PCR试剂盒说明书操作。反应体系均为10 μL,包括:5×gDNA Eraser 缓冲液2 μL,gDNA Eraser 1 μL,Total RNA 小于1 μg,剩余用RNase Free dH2O补至10 μL。反应条件为42 ℃反应2 min。
逆转录反应:逆转录反应体系均为20 μL,包括:上一步骤的反应液10 μL,PrimeScript RT Enzyme MixⅠ1 μL,RT Primer Mix 4 μL,5×PrimeScript buffer 2(for Real time) 4 μL,RNase Free dH2O 1 μL。反应条件为37 ℃反应15 min,85 ℃反应5 s。反应产物均为cDNA。
2.4.2 荧光定量PCR检测表皮葡萄球菌ATCC 35984菌株luxS,agr,RNAⅢ转录水平 分别用反转录获得的cDNA作为模板,进行荧光定量PCR以检测内参基因gryB和目的基因luxS,agr,RNAⅢ的转录水平。每样本做3复孔,同时设置阴性对照。按照SYBR Premix Ex TaqⅡ(Tli RNase H Plus)试剂盒说明书操作,引物参照表1并配制成10 μmol·L-1。反应体系为20 μL,包括:SYBR Premix Ex TaqⅡ(Tli RNaseH Plus)(2×)10 μL,10 μmol·L-1引物(正向、反向)各0.8 μL,ROX Reference DyeⅡ(50×)0.4 μL,RT反应液(cDNA溶液)2 μL,dH2O(灭菌蒸馏水)6 μL。实时荧光定量PCR扩增仪QuantStudio TM 6 Flex进行检测。扩增条件:95 ℃预变性30 s;95 ℃变性5 s,60 ℃退火34 s,共40個循环,60 ℃收集荧光信号。最后做融解曲线分析,确定反应产物的单一性。
2.5 数据分析
2.5.1 qRT-PCR结果计算 自动调节基线(base line)至适宜处,各扩增曲线与阈值线的交叉点对应的横坐标即为Ct值,根据标准曲线上浓度与Ct值的对应关系,可求出各待测标本的初始浓度。以gryB作内参基因,luxS/gryB,agr/gryB,RNAⅢ/gryB作为衡量luxS,agr,RNAⅢ转录水平的指标。
2.5.2 统计学分析 将设置的每个时间段未经药物处理的ATCC 35984 luxS,agr,RNAⅢ 的表达量作为“1”,计算出相对量,再用SPSS 10.0统计软件进行单因素方差分析,两样本之间进行t检验,数据以±s表示,P<0.05为差异有统计学意义。
3 结果
3.1 鱼腥草素钠、红霉素、万古霉素对产膜表皮葡萄球菌MIC的结果
鱼腥草素钠对ATCC 35984 MIC为64 mg·L-1,万古霉素对ATCC 35984 MIC为8 mg·L-1,红霉素对ATCC 35984 MIC为8 mg·L-1。
3.2 药物干预后目的基因表达的结果
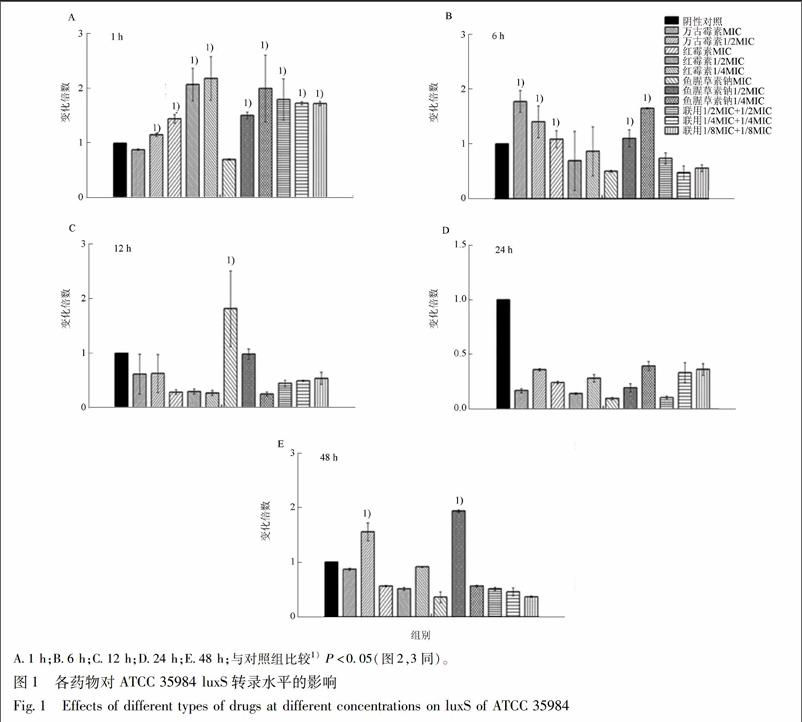
3.2.1 药物对luxS表达的影响 1/2MIC,1/4MIC鱼腥草素钠,1/2MIC鱼腥草素钠+1/2MIC红霉素,1/4MIC鱼腥草素钠+1/4MIC红霉素,1/8MIC鱼腥草素钠+1/8MIC红霉素等在作用于ATCC 35984伊始,就可迅速上调luxS的表达,与阴性对照相比,存在显著差异(P<0.05);在作用6,12,48 h后,鱼腥草素钠依然存在对luxS的上调作用,见图1。
3.2.2 药物对agr表达的影响 MIC,1/2MIC的鱼腥草素钠在作用的各时间段对agr表达均有显著下调作用(P<0.05),见图2;1/2MIC鱼腥草素钠与1/2MIC红霉素联用、1/4 MIC鱼腥草素钠与1/4MIC红霉素联用,在作用6,12,24 h等时间段,对agr表达的下调作用也很显著(P<0.05),见图2。
3.2.3 药物对RNAⅢ表达的影响 MIC的鱼腥草素钠在作用的各时间段对RNAⅢ表达亦有显著下调作用(P<0.05),见图3;1/2MIC鱼腥草素钠与1/2MIC红霉素联用对RNAⅢ表达的下调效果也很显著(P<0.05),见图3。
4 讨论
表皮葡萄球菌生物被膜的形成是个动态过程。首先是由菌体表面疏水性蛋白或多糖黏附素对生物材料的初始附着,形成细菌群落;随后细菌相互聚集,形成生物被膜[8]。其中,PIA是细菌生物被膜形成的聚集阶段所必需的物质[9]。Ica基因座编码PIA,由icaA,icaD,icaB,icaC 4种基因组成,形成操纵子icaADBC;其中,icaA在PIA的形成中起决定作用。
luxS在多种革兰阳性菌和革兰阴性菌中均高度保守,这些微生物能够产生类似的AI-2信号分子,因此AI-2分子被认为是各种细菌进行中间交流的通用语言,luxS为AI-2形成的标志基因。Xu等[5]通过构建和分析△luxS突变株,研究了表皮葡萄球菌中的luxS/AI-2感应系统的活性;模型证实luxS通过AI-2的细胞间信号转导下调ica操纵子,因而影响了PIA的产生,减少了细菌间黏附聚集,抑制生物被膜的形成,并减弱了它在动物体内的致病力。
生物被膜并不是细菌经常性的生活方式,生物被膜成熟后,膜内的细菌还会从膜内分散出来形成新的感染灶。细菌从生物被膜上的分离,是导致疾病扩散的生理基础之一。目前已知的与细菌分散有关的因素包括:QS系统、表面活性剂、基质降解酶等;在葡萄球菌中,分散的机制是通过agr系统来调控的。葡萄球菌的致病能力和所致感染的严重程度还与其产生的毒素有关。葡萄球菌的agr亦是最重要的毒力因子调节系统,负责对毒力因子生长阶段进行依赖性调节[10];RNAⅢ是agr系统的效应器分子。RNAⅢ可编码δ-毒素,δ-毒素具有去垢剂样作用,可以帮助生物被膜形成输水及营养通道,并可使生物被膜基质降解,有助于生物被膜中的细菌脱离等[11];也有学者研究表明,RNAⅢ还可以上调细胞外毒性因子的产生和下调细胞壁表面相关蛋白的转录,促进外毒素的产生[12]。
针对生物被膜形成过程的治疗方法包括阻止细菌起始黏附及细菌间相互黏附;阻断QS机制所需的生物被膜组件基因的表达;抑制组成生物被膜基质多聚糖和胞外蛋白的生物合成;降解生物被膜基质所需酶类等[13]。目前,临床抗葡萄球菌感染常使用的抗生素以红霉素、环丙沙星为代表,但临床耐药性的问题还很突出;万古霉素作为抗葡萄球菌有效的药物,但临床对其中度耐药的表皮葡萄球菌也逐渐增多[14]。因此,寻求有效抗生物被膜菌的药物,研究中西药联用以达到药物减量增效、降低毒副作用等措施,已日益得到人们的重视。
鱼腥草,性寒味辛,具有清热解毒、行水消肿、祛瘀生新之功效,入肝肺二经。体外试验已证明,鱼腥草煎剂对多种革兰阳性菌、革兰阴性菌均有不同程度的抑制作用。鱼腥草中抗致病微生物的主要成分是挥发油,挥发油中主要为癸酰乙醛。癸酰乙醛已能人工合成,被称为鱼腥草素,但该种物质性质不稳定,而其亚硫酸氢钠加成物(鱼腥草素钠),则性质稳定而又保留其抗菌活性。
基于以往的研究[7],发现包括亚抑菌浓度在内的鱼腥草素钠及与红霉素联用,对ATCC 35984的初始黏附及在生物被膜内的代谢均有明显的抑制作用,并干预了生物被膜的形成。本实验在不同时间段(时间段的设置主要参考表皮葡萄球菌生物被膜形成的一般规律[15];0~4 h完成初始黏附,6 h黏附的细菌开始相互聚集,24 h生物被膜基本成熟,30~48 h部分生物被膜开始崩解),观察各药物对悬浮状态下的产膜表皮葡萄球菌luxS,agr,RNAⅢ的转录水平的影响;结果显示鱼腥草素钠及其与红霉素联用可以迅速上调luxS的表达,效果并不弱于万古霉素、红霉素等抗生素,从药物作用的持久性来看,鱼腥草素鈉的效果甚至优于抗生素,提示鱼腥草素钠及与红霉素联用可调控luxS,因此具有抑制表皮葡萄球菌生物被膜的形成,并减弱其致病力的可能。鱼腥草素钠与红霉素联用在作用初期对luxS的上调作用尤为明显,但作用持续时间较短,具体原因尚不清楚。
鱼腥草素钠在MIC,1/2MIC浓度时,对agr表达均有显著下调作用(图2);1/2MIC鱼腥草素钠与1/2MIC红霉素联用、1/4 MIC鱼腥草素钠与1/4MIC红霉素联用,在作用6,12,24 h等时间段,对agr表达的下调作用也很显著。同样,也发现鱼腥草素钠在MIC浓度时,对RNAⅢ表达亦有显著下调作用;1/2MIC鱼腥草素钠与1/2MIC红霉素联用对RNAⅢ表达的下调效果也很显著;提示鱼腥草素钠及与红霉素联用,在特定浓度下,可显著下调agr,RNAⅢ的表达;因此,就具有抑制表皮葡萄球菌生物被膜内营养和输水通道的形成、阻止生物被膜基质降解和生物被膜内细菌脱离分散、并抑制细菌外毒素形成等作用的可能。
luxS,agr均是表皮葡萄球菌QS系统的关键基因;由于细菌的QS系统是调控非细菌生存所必须的基因,以其作为药物靶标不会对细菌生存产生较大压力,因此不易使细菌产生耐药性[16],从而对筛选抗表皮葡萄球菌药物靶标有着重要指导意义。RNAⅢ分子是调控葡萄球菌毒力表达的关键分子,通过体外实验研究已证实[17-18],RNAⅢ抑制肽可以有效抑制金黄色葡萄球菌肠毒素和溶血素的产生,可以抑制表皮葡萄球菌对关节假体材料表面的黏附。本研究亦证实了特定浓度的鱼腥草素钠及与红霉素联用对luxS的上调和对agr,RNAⅢ的下调作用,为评估鱼腥草素钠及与红霉素联用成为抗表皮葡萄球菌生物被膜感染药物,提供新思路及实践依据,也可为临床用药提供参考。但细菌的QS系统毕竟是很复杂的调控系统,许多机制尚未被完全了解,并且作者发现药物在对luxS,agr,RNAⅢ等调控随药物浓度的变化而改变,其规律需在以后实验中进一步探究。另外,本实验是以强产膜标准株ATCC 35984为实验对象,今后还应扩大对临床株样本的实验,以使结果更加客观、可信。
[参考文献]
[1] Qin Z,Yang X,Yang L,et al. Formation and preperties of in vitro biofilms of ica-negative Staphylococus epidermidis clinical isolates[J]. J Med Microbiol,2007,56(1):83.
[2] Nobile C J,Mitchell A P. Microbial biofilms: epluribus unum[J]. Curr Biol,2007,17(10):R349.
[3] 刘琳,谭小娟,贾爱群. 细菌群体感应与细菌生物被膜形成之间的关系[J].微生物学报,2012,52(3):271.
[4] Davies D G,Parsek M R,Pearson J P,et al. The involvement of cell-to-cell signals in the development of a bacterial biofilm[J]. Science,1998,280(5361):295.
[5] Xu L,Li H,Vuong C,et al. Role of the luxS quorum-sensing system in biofilm formation and virulence of Staphylococcus epidermidis[J]. Infect Immunol,2006,74(1):488.
[6] Li J,Wang W,Xu S X,et al. Lactobacillus reuteri-produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci[J]. Proc Natl Acad Sci USA,2011,108(8):3360.
[7] 官妍,李春,石晶金,等.亚抑菌浓度鱼腥草素钠及与红霉素联合对表皮葡萄球菌生物被膜的作用[J].中国中药杂志,2013,38(5):731.
[8] Qin Z,Qu Y,Yang L,et al. Role of autolysin mediated DNA release in biofilm formation of Staphylococcus epidermidis[J]. Microbiology,2007,153(pt7):2083.
[9] Jeng W Y,Ko T P,Liu C I,et al. Crystal structure of IcaR,a repressor of the TetR family implicated in biofilm formation in Staphylococcus epidermidis[J]. Nucleic Acids,2008,36(5):1567.
[10] Vuong C,Gerke C,Someroille G A,et al. Quorum-sensing control of biofilm factors in Staphylococcus epidermidis[J]. J Infect Dis,2003,188(5): 706.
[11] Kiran M D,Akiyoshi D E,Giacometti A,et al. OpuC-an ABC transporter that is associated with staphylococcus aureus pathogenesis[J]. Int J Artif Organs,2009,32(9):600.
[12] Tsompanidou E,Sibbald M J,Chlebowicz M A,et al. Requirement of the agr locus for colony spreading of staphylococcus aureus[J]. J Bacteriol,2011,193(5):1267.
[13] Benghezal M. Inhibitors of bacterial virulence identified in a surrogate host model[J]. Cell Microbio,2007,9(5):1336.
[14] Otto M. Molecular basis of Staphylococus epidermidis infections[J]. Semin Immunopathol,2012,34(2):201.
[15] 叶联华,黄云超,杨达宽,等. 聚氯乙烯材料表面细菌生物膜结构观察[J].生物医学工程与临床,2007,11(4):251.
[16] Vendeville A,Winzer K,Heurlier K,et al. Making ‘sense of metabolism: Autoinducer-2,LuxS and pathogenic bacteria[J]. Nat Rev Microbiol,2005,3:383.
[17] 邢庆昌,郝立波,王继芳. 应用RNAⅢ抑制肽(RIP)抑制葡萄球菌毒素产生的實验研究[J].科学技术与工程,2011,11(2):317.
[18] 郝立波,邢庆昌,王继芳,等. RNAⅢ抑制肽抑制葡萄球菌在人工关节材料表面粘附的实验研究[J].中国矫形外科杂志,2008,16(23):1814.
[责任编辑 张宁宁]
