囊盘菌(Ascocoryne)挥发性气体成分多样性及其产烃分析
2017-06-19严东辉王凯英李鸿昌
高 茜,严东辉*,王凯英,李鸿昌,任 菲
(1.中国林业科学研究院森林生态环境与保护研究所,北京 100091; 2. 中国林业科学研究院华北林业实验中心,北京 102300)
囊盘菌(Ascocoryne)挥发性气体成分多样性及其产烃分析
高 茜1,严东辉1*,王凯英1,李鸿昌1,任 菲2
(1.中国林业科学研究院森林生态环境与保护研究所,北京 100091; 2. 中国林业科学研究院华北林业实验中心,北京 102300)
[目的]内生真菌紫胶囊盘菌(菌株NRRL 50072)首次被发现能产生石化燃料类似成分的挥发性有机代谢产物,这为生物质能源的发展开辟了新的途径,但囊盘菌属的真菌是否都具有该类代谢属性需要进一步明确。[方法]通过组织表面消毒分离囊盘菌不同菌株,经最大似然法构建分子系统发育树明确亲缘关系,利用顶空固相微萃取方法结合气质联用技术(HSSPEM-GC-MS),进行挥发性成分测定和分析。[结果]不同囊盘菌菌株均能产生挥发性烃、醇、酯、醛、酮和酸等代谢产物,但产物存在高度的多样性,即使近缘的菌株之间其挥发性产物也存在很大差异。[结论]囊盘菌属真菌可能普遍具有产挥发性烃类的能力,但其代谢产物种类具有菌株专化性。因此,发现或遗传工程创造目标新菌株资源是深入研究和开发利用囊盘菌真菌生物柴油的重要基础。
Ascocoryne;挥发性气体;SPEM-GC-MS;生物质能;烃类化合物
由于石化燃料终将耗尽,寻求可更新的替代能源资源日趋重要,生物质能是极具发展潜力的可替代能源之一,尤其是不争耕地、能利用农林废弃物木质纤维进行能源转化的生物质资源受到重视[1]。真菌作为地球上最主要的木质纤维分解者,不仅成为醇基燃料生物质能转化过程中木质纤维素酶的重要来源;而且最近研究还发现,一些植物内生真菌在分解利用木质纤维的同时,能直接产生类似柴油、汽油和航空用油的成分[1-2]。由于内生真菌通常是植物死亡或近于死亡时的第一批分解者[3],这使得内生真菌不仅可以作为提供木质纤维素分解酶的来源[4],也为探索直接利用内生真菌分解农林废弃物产生所需燃料奠定了基础,使之成为可更新和兼容现存引擎基础设施的生物质能新燃油资源[2]。
目前已发现多种内生真菌能产生类似汽油或柴油的挥发性成分,如来自Ascocoryne、Muscodor、Hypoxylon等属的内生真菌、以及内生菌Nodulisporiumsp.、Phomasp.和Phomopsissp.中的菌株其挥发性产物中存在烷烃、烯烃、烷基酯、苯衍生物,萜类化合物、多环芳烃等[2]。其中AscocornyesarcoidesNRRL 50072 由Strobel等在巴塔哥尼亚(Patagonia)的植物Eucryphiacordifolia上分离获得,并能在木质纤维基质上产生酸、醇及烷烃的酯类和酮类衍生物[5,6]。该菌最先通过形态鉴定为Clonostachysrosea(=Gliocladiumroseum)[5],后经分子鉴定,结合对比参考菌株AscocorynesarcoidesAV-70培养性状, 更正为Ascocorynesarcoides[7]。该属真菌属子囊菌,子实体常发现在秋季阔叶林针叶林潮湿环境中的枯木中产生,但其菌丝在健康树木组织内长期行内共生生活,具内生真菌属性,至树木死亡才在枯木上开始形成子实体[8],我国记录有两个种,分别为A.sarcoides和A.cylichnium, 其中A.sarcoides是世界广布性种。该菌菌株(A.sarcoidesRRL 50072) 产生相对丰富的挥发性烃类,可能与其进化有关;在其基因组上发现有80多个可能与该生物代谢合成相关基因簇的存在[9]。并且发现生长阶段,pH、温度和培养基质组成等因素对该菌产生烃类型有显著的影响[10]。但迄今我们并不清楚产烃代谢途径只是该菌株的个体遗传行为,还是该属同种或近似种共有的群体属性。因此,我们利用顶空固相微萃取技术结合气质联用分析仪方法,对来自囊盘菌的不同菌株挥发性产物进行了研究分析。结果发现相同环境背景下,不同菌株所产生的挥发性代谢成分具有多样性。
1 材料与方法
1.1 菌株来源
供试Ascocoryne属菌株6株,编号分别为FPYF3013-FPYF3018。其中FPYF3013-FPYF3016采集分离于国内海南尖峰岭和北京雾灵山地区山毛榉科树木上。FPYF3017和FPYF3018购自德国DSMZ菌库(保藏号分别为DSM4844、DSM4705)。上述6株菌株均保存在中国林科院森林病理室。
1.2 菌株鉴定
培养性状观察: 预先在PDA上培养7 d的菌落作为接种材料。用打孔器在菌落边缘取直径约0.5 cm圆片转接入PDA和MEA培养基中,培养皿直径为8 cm,23℃培养,隔天观察记录菌落生长直径和培养形状,至满皿为止。
分子鉴定: 菌株基因组DNA提取参考植物CTAB法[11]。 用于系统发育分析的分子标记序列为ITS,所用扩增引物为真菌通用引物ITS1和ITS4[11]。扩增反应及程序参考Amar等的方法[11]。扩增产物测序由博迈德生物技术有限公司完成。序列读取和序列拼接均用Bioedit0.7进行。完整ITS序列递交NCBI获取序列号。
系统发育树构建:参与构建系统发育树的Ascocoryne参考菌株ITS序列来自NCBI,其序列登录号见表1,参考菌株的来源地点等背景材料可通过NCBI网站登录了解。截止到2016年8月,在NCBI中有Ascocoryne的ITS序列录入数共52条,其中只有3个明确种,分别是A.sarcoides、A.cylichnium和A.solitaria;其余为不确定种记录。序列利用MAFFT在线(http://mafft.cbrc.jp/alignment/software/)进行联配比对,通过MrModel3.0获得建树模型(GTR)及其参数,运用PhyML3.1构建ML系统发育树,自检值设定为1000。
1.3 菌株产挥发性成分分析
1.3.1 顶空固相微萃取(HS-SPME) 将菌种接种到PDA培养基上,每个菌株重复3次;放置于恒温培养箱培养18天,取出备用。萃取头老化(气相法)温度分别为柱温150℃,后进样口 温度250℃,前检测器 200℃,老化时间15 min。萃取过程:手动操作萃取。用0.6 mm电钻在培养皿的侧面穿孔,插入萃取针(50/30 μm DVB/CAR/PDMS,SUPELCO),推出萃取纤维,萃取时间40 min。
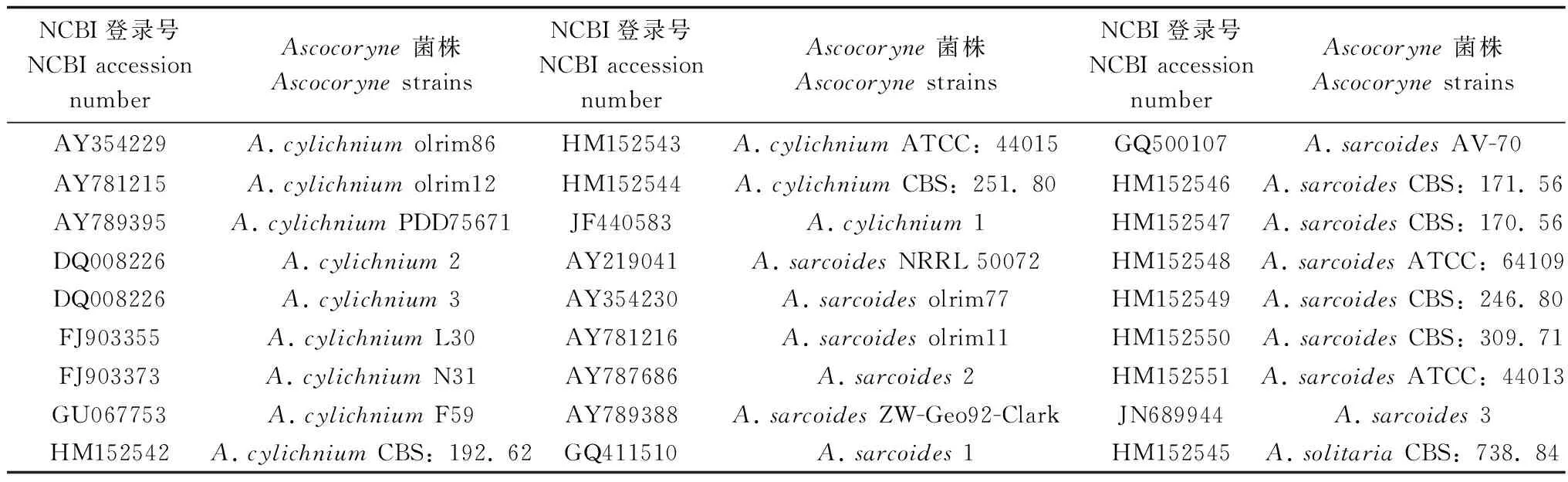
表1 参与建树的参考菌株NCBI登录号
1.3.2 气相色谱-质谱联用分析(GC-MS) 气相色谱条件HP-5MS型毛细管柱(涂层:(5%苯基)-甲基聚硅氧烷,30.0 m×0.25 mm,涂层厚度:0.25 μm);升温程序如下:初始温度33℃,保留2 min;以7℃·min-1的速度升温至220℃,保留7 min;载气为氮气,流速为39 mL·min-1。
质谱条件:EI电离源,电子能量70 eV,离子源温度150℃,质谱扫描速率为:5.000 0 scans·s-1。质谱范围41~560 amu(m·z-1)。
1.3.3 数据处理 产物成分鉴定由质谱仪中NIST 数据库匹配求得,统计匹配度大于80(最大值为100)的鉴定结果才予以报道。
2 结果与分析
2.1 供试菌株及与A.sarcoidesNRRL 50072菌株间的系统发育关系
供试6菌株的ITS序列同源性搜索结果显示,这些菌株各自的最高同源性菌株均来自Ascocoryne。同源性参数覆盖率(cover)、E值、一致性(identity)中,除FPYF3014和FPYF3015与各自最高同源Ascocoryne参考菌株的覆盖率为99%外,其余菌株与各自最高同源参考菌株覆盖率达100%;E值和一致性上,所有菌株均分别为0和100%。因此,这些菌株鉴定为Ascocoryne属的真菌,且依据最高同源参考菌株,FPYF3013、FPYF3014及FPYF3018可能为A.sarcoides;FPYF3015可能为A.cylichnium; FPYF3016和FPYF3017则可能是A.solitaria。 这结果与用另一分子标记基因ACTIN序列搜索的结果一致(数据未列出)。来自这些菌株的ITS序列已获得NCBI编号,依FPYF3013-FPYF3018次序分别为KX976479-KX976484。

图1 基于ITS序列的囊盘菌属真菌ML系统发育树。Fig.1 Phylogenetic analysis on the Ascocoryne strains based on ITS sequences by ML method
依据ITS序列建立的最大似然法的系统发育树(图1)可见,FPYF3013、FPYF3014和FPYF3018与A.sarcoides的菌株聚在一起,聚类支持率68%; FPYF3014和FPYF3018同A.sarcoidesNRRL50072的亲缘关系得到更高的87%支持率。FPYF3015与A. cylichnium的亲缘关系也有近于70%的支持率。而FPYF3016和FPYF3017则散成单系(singleton),与作为参考的来自Ascocoryne所有菌株无聚类关系。可以肯定地认为这两菌株远离A.cylichnium和A.sarcoides,而与A.solitaria存在可能的亲缘关系。这与在NCBI中进行BLASTn的结果基本一致。
2.2 菌株生长差异
菌株在PDA和MEA培养基的培养性状表明,除FPYF3015外,其余5株菌株均向培养基中渗透紫色色素且程度不等,而FPYF3015的菌落不具有该渗透特点。向培养基质中分泌渗透性紫色色素是该属Ascocornye的典型培养性状之一。6株菌株的生长情况在PDA和MEA上表现有差异(图2,3)。在PDA上(图2),6株菌株菌落的生长速度,FPYF3018最快,FPYF3016最慢,二者菌落扩展至满皿(直径8cm)的日期相差8天。期间生长速率快慢菌株依次为FPYF3013、3017、3015和3014;并且FPYF3013两周后生长变缓,菌落直径上分别被FPYF3017和FPYF3015超过。FPYF3014在试验末期,扩增速度出现下降。在MEA培养基上(图3),菌株之间的生长情况有些变化,FPYF3018菌落扩展速度依旧是最快的,而最慢的菌株是FPYF3015,二者满皿生长相差10天。其它菌株菌落的扩展速度从快到慢依次是FPYF3014、FPYF3013、FPYF3017和FPYF3016。FPYF3013在生长近两周时,生长速率同PDA中的生长表现出现些微下降,而与FPYF3017相近(图2)。总体上,6菌株在MEA培养基上比在PDA培养基上生长快,依不同菌株菌落扩展至满皿而言,可以有4~10天的生长时间差异,如FPYF3018在PDA和MEA长满培养皿需要天数分别是12天(图1)和16天(图2),而FPYF3014则相差10天,这说明不同营养对菌株生长有很大影响。
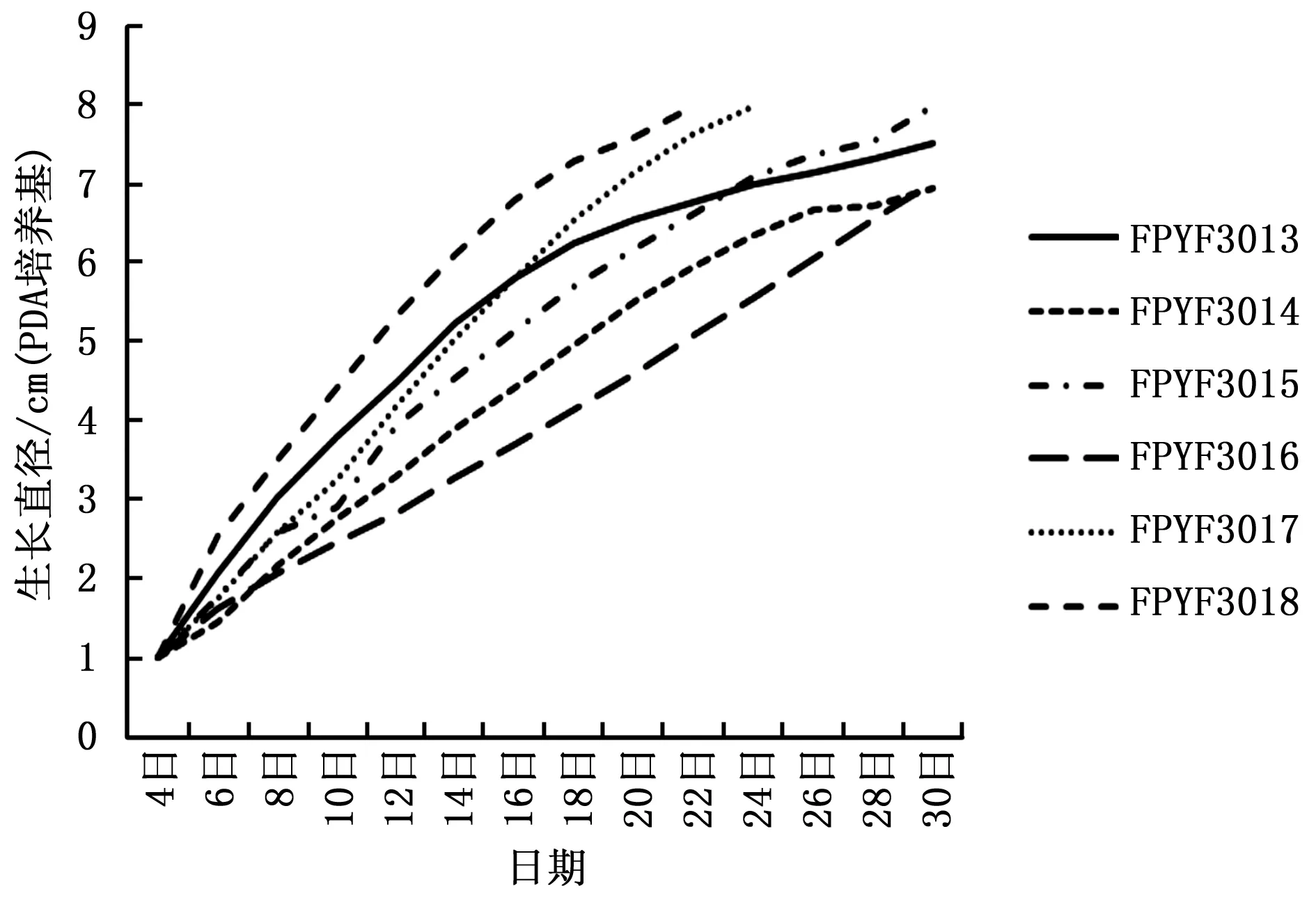
图2 囊盘菌6菌株在PDA培养基中25℃条件下的生长速率Fig.2 Growth rates of Ascocoryne fungi in PDA at 25℃
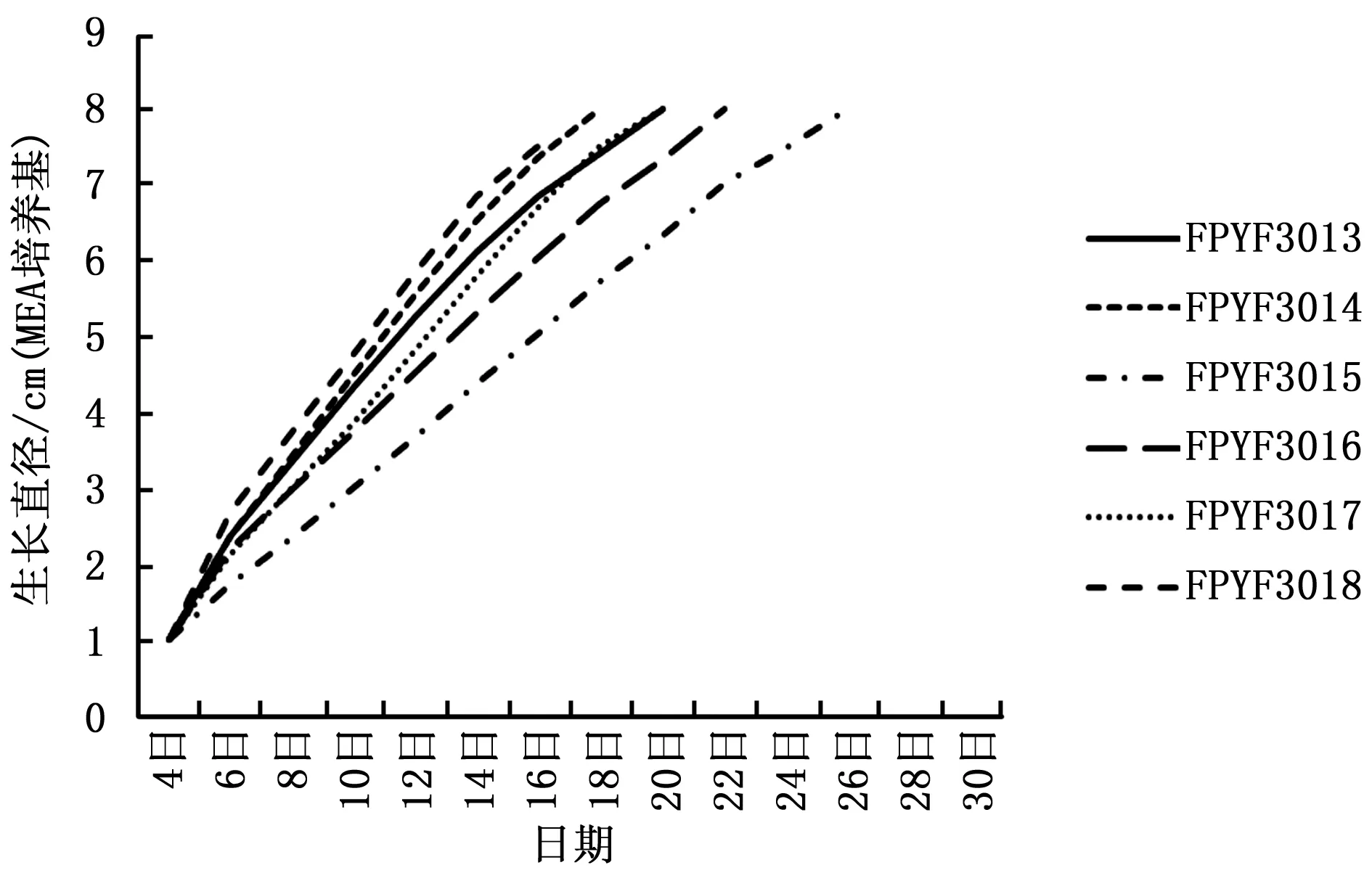
图3 囊盘菌6菌株在MEA培养基中25℃条件下的生长速率Fig.3 Growth rates of Ascocoryne fungi in MEA at 25℃
2.3 菌株挥发性成分分析
供试6个菌株在PDA培养基中产出80%以上匹配度的挥发性成分较多。按烃、醇、酯、醛、酮和酸等大类及具体化合物,分别总结于表2及表3。从中可见,菌株FPYF3016仅检测出3类5种挥发物, FPYF3013-3015菌株能产出5~6类中的10~20种化合物;FPYF3017和FPYF3018检测出6~7类的30种化合物。其中,FPYF3013和FPYF3015的烃类成分是优势化合物,占各自总成分的55.6%(10/18) 和62.3%(9/14);FPYF3014、FPYF3017和FPYF3018则是醇类成分占优势,可分别占有各自总成分的41.2%、33.3%(10/30)和40.0%(12/30);所有菌株都产少量的酯、酮和酸等成分,而醛类仅有FPYF3013产生,且只有1种化合物(表2)。具体成分上(表3),6株菌株均能产生3-甲基-1-丁醇;除FPYF3017外的5株菌株能产2-甲基-1-丙醇;除FPYF3015外的5株菌株能产乙醇。烷烃类产物上,两株菌株FPYF3016和FPYF3017未检出烷烃成分;FPYF3013、3034、3015及FPFY3018菌株共产烷烃12种。这些烷烃产物中,直链烷是1个十六烷由FPYF3013产出、1个二十七烷由FPYF3015产生,其余10种全是支链烷。这些支链烷中,分子量最低的是含两甲基支链共5个C的丙烷,由FPYF3013产生;最高的是含1个甲基支链的十六烷,由FPYF3015产生。支链庚烷有两个,分别由FPYF3014和FPYF3018产生;支链葵烷有三个,分别由FPYF3014、FPYF3015和FPYF3018产生;十一支烷两个,分别由FPYF3013和FPYF3018产生;十六支烷一个,由FPYF3015产生。菌株之间没有相同的烷烃产物。产烯烃情况是,菌株FPYF3016未检测到烯烃,菌株FPYF3018只检测到一种烯烃δ-塞林烯,菌株FPYF3014有3种烯烃,分别是α-Muurol烯、环己烯和萘,其余三株菌株能产5-6种烯烃,这些烯烃主要为C15H24的不饱和烃。

表2 囊盘菌6菌株挥发性产物归类统计

表3 囊盘菌6菌株挥发性产物
续表3

编号number化合物(英文)compounds分子式molecularformula化合物(中文)compoundsAscocoryne菌株Ascocorynestrains131415161718391H-Cycloprop[e]azulene,1a,2,3,4,4a,5,6,7b-octahydro-1,1,4,7-tetramethyl-,[1aR-(1aα,4α,4aβ,7bα)]-C15H241H-环丙并[e]薁,1A,2,3,4,4a,5,6,7b八氢1,1,4,7四甲基-[1aR-(1aα,4α,4aβ,7bα)]-√-----40δ-SelineneC15H24δ-塞林烯----√√412-DecanolC10H22O2癸醇-√--√√42BenzaldehydeC7H6O苯甲醛√----√431H-Cycloprop[e]azulene,1a,2,3,4,4a,5,6,7b-octahydro-1,1,4,7-tetramethyl-,[1aR-(1aα,4α,4aβ,7bα)]-C15H241H-环丙并[e]薁,1A,2,3,4,4a,5,6,7b八氢1,1,4,7四甲基-[1aR-(1aα,4α,4aβ,7bα)]---√-√-44Propanoicacid,2-methylC4H8O2丙酸,2-甲基√-----451-Octanol,2-methylC9H20O1-辛醇,2-甲基----√-461S,2S,5R-1,4,4-Trimethyltricyclo[6.3.1.0(2,5)]dodec-8(9)-eneC15H241S,2S,5R-1,4,4-三甲基三环[6.3.1.0(2,5)]十二碳-8-(9)-烯√-----472-Isopropenyl-4a,8-dimethyl-1,2,3,4,4a,5,6,8a-octahydronaphtha-leneC15H242-异丙烯基-4a,8-二甲桥-1,2,3,4,4a,5,6,8a八氢萘--√---48β-HumuleneC15H24β-律草烯√-----49AromandendreneC15H24香橙烯----√-50α-MuuroleneC15H24α-Muurol烯-√--√-512-DodecanoneC12H24O二十二烷酮-----√52Aceticacid,decylesterC12H24O2乙酸,癸酯-----√53β-GuaieneC15H24β-乖烯----√-54Butanoicacid,3-methylC5H10O2丁酸,3-甲基-----√55Hexanoicacid,2-methyl-C7H14O2己酸,2-甲基√-----56Naphthalene,1,2,4a,5,6,8a-hexahydro-4,7-dimethyl-1-(1-meth-ylethyl)-C15H24萘,1,2,4a,5,6,8a-六氢-4,7-二甲基-1-(1-甲基乙基)-√--√-571-Undecanol,acetateC13H26O21十一醇,醋酸----√-58cis-(-)-2,4a,5,6,9a-Hexahydro-3,5,5,9-tetramethyl(1H)benzo-cyclohepteneC15H24顺-(-)-2,4a,5,6,9a六氢3,5,5,9四甲基(1H)-苯并环庚烯----√-593-Cyclohexene-1-methanol,α,α,4-trimethyl-,acetateC12H20O23-环己烯-1-甲醇,α,α,4-三甲基,乙√-----60α-IononeC13H20Oα-紫罗兰酮√-----61Cyclohexene,3-methyl-6-(1-methyle-thylidene)-C10H16环己烯,3-甲基-6-(1-甲基亚乙基)√√----621-DecanolC10H22O1-癸醇-----√63AzuleneC10H8薁--√---64Cyclodecane,methyl-C11H22癸烷,甲基----√-65Formicacid,undecylesterC12H24O2甲酸,十一烷基酯----√-663,6-Octadien-1-ol,3,7-dimethyl-,(Z)-C10H18O3,6-辛二烯-1-醇,3,7-二甲基-,(Z)---√---67n-Tridecan-1-olC13H28O正十三烷-1-醇-----√68BenzylalcoholC7H8O苯甲醇-√----691H-Indene,1-ethylidene-C11H10-1H-茚,1-亚乙基--√---70Naphthalene,1-methyl-C11H10萘,1-甲基--√---71(Z)6-Pentadecen-1-olC15H30OZ-6-十五碳烯-1-醇-----√72PhenylethylAlcoholC8H10O苯乙醇-√--√√731-HexadecanolC16H34O1-十六烷醇-----√741,3a-Ethano(1H)inden-4-ol,octa-hydro-2,2,4,7a-tetramethyl-C15H26O1,3A-Ethano(1H)茚-4-醇,八氢2,2,4,7a四甲基----√-75CyclotetradecaneC14H28环四癸烷-----√76Phenol,2,4-bis(1,1-dimethyleth-yl)-C14H22O苯酚,2,4-双(1,1-二甲基乙基)------√77MethylhinokiateC16H24O2甲基hinokiate----√-78Nootkaton-11,12-epoxideC15H22O2香柏酮基-11,12-环氧----√-
续表3

编号number化合物(英文)compounds分子式molecularformula化合物(中文)compoundsAscocoryne菌株Ascocorynestrains131415161718792-Naphthalenemethanol,1,2,3,4,4a,5,6,8a-octahydro-α,α,4a,8-tetramethyl-,[2R-(2α,4aα,8aβ)]-C15H26O2-萘甲醇,-1,2,3,5,6,8a八氢-α,α,4α,8-四甲基-,〔2R-(2α,4aα,8αβ)〕----√-802,4,7,14-Tetramethyl-4-vinyl-tri-cyclo[5.4.3.0(1,8)]tetrade-can-6-olC20H34O2,4,7,14四甲基-4-乙烯基-三环[5.4.3.0(1,8)]四癸-6-醇----√-
3 讨论
囊盘菌属属于子囊菌门Ascomycota、柔膜菌目Helotiales、柔膜菌科Helotiaceae,目前在真菌库(Mycobank,http://www.mycobank.org)列有9种、在全球生物物种名录(http://www.catalogueoflife.org/)中登记有已知种8种、真菌索引(Index Fungorum,http://www.indexfungorum.org/)中收录6种。在NCBI核酸数据库中,仅有A.sarcoides、A.cylichnium和A.solitaria这3个种菌株的ITS参考序列,其它种的分子系统发育分子标记核酸序列缺乏,这说明目前该属真菌认识较稳定和清楚的只有这3个种的菌株。并且,该属真菌因为其生态功能和重要性不被了解和重视,其种类鉴定主要还是依据形态特征,缺乏基于核酸序列的系统发育关系的认识,这种情况常造成菌株在分类地位上不一致。本文用于建系统发育树的参考菌株A.cylichnium没有共聚一个分支的原因,很大程度上与此相关(图1),菌株FPYF3014归属种类则主要参考BLASTn结果中的最高同源性菌株。该属真菌基于核酸序列的系统发育关系亟待建立。本研究供试的6个菌株涉及该3个种,并且A.sarcoides和A.solitaria各含2个菌株,因此,本研究的结果对该属真菌可以具有一定的代表性。我们的研究结果还显示,FPYF3013和FPYF3018与已知产烃的A.sarcoidesNRRL 50072菌株亲缘关系高度同源。NRRL 50072菌株是以A.sarcodiesAV-10为参考菌株,从Gliocladiumroseum[5]订正为A.sarcoides的[7]。本文Ascocoryne属系统发育树中这2株菌株与FPYF3013和FPYF3018菌株,以87%高可信度聚类在同一簇上(图1)。这结果支持了菌株AV-10和NRRL 50072之间的这一关系[7]。同时,由于NRRL 50072的产烃性能,使得同一聚类组中的其他10多个菌株的产烃情况理应也受到关注研究。本文的结果还表明了所供试的3个种的6个菌株之间,在ITS序列、系统发育和培养性状都有明显的差异(图1-3);在生长速度上,不仅种间菌株,种内菌株之间也有差异。因此,6个菌株在遗传背景上是存在差异的。
囊盘菌菌株A.sarcoidesNRRL 50072在PDA培养基上,能产生烃类物质2,6-二甲基十一烷(undecane, 2,6-dimethyl)、3,3,5-三甲基葵烷(decane, 3,3,5-trimethyl)、4-甲基环己烷(cyclohexene,4-methyl)、3,3,6-三甲基葵烷(decane,3,3,6-trimethyl)和4,4-二甲基十一烷(undecane, 4,4-dimethyl)等庚烷、辛烷、苯、和一些支链烃挥发性代谢产物[5],这些产物中含有不少与现有石化柴油或汽油燃料相同的烃类成分,因此,该菌的挥发性化合物被称为“微生物柴油”(mycodisele)[2]。虽然之后在其他属的内生真菌发现有产类似石化燃料成分的挥发性代谢产物[2],但在Ascocoryne属真菌的其它菌株上还未见有产该类代谢产物的后续报道。我们的研究结果显示(表2、3),分别来自A.sarcoides和A.cylichnium的3个菌株中,每株能产生3种以上的烷烃,而来自A.solitaria的2个菌株未见烷烃产物;能产3种以上烯烃成分的菌株有5株,这5株菌株包括来自不产烷烃的A.solitaria1株。可见虽然本属真菌产挥发烃类物质可能是个常规属性,至少对来自A.sarcoides和A.cylichnium的菌株而言是如此,但最终的烃产物之间是会存在差别的。结果中(表3)表明,仅有一种产物2,6,11-三甲基十二烷(产物编号19)在3个菌株的VOC中存在。菌株FPYF3013和FPYF3018所产生的挥发性烃类(表3中产物编号12、19、20、24和6、15、19)与近缘的A.sarcoidesNRRL 50072相比较[5],也无相同之处,这显示出该属真菌多样性成员之间的烃产物上也存在多样性,而这是否是各自的产烃代谢调控过程或途径具有多样性?是个值得进一步揭示的问题,也可能与菌株间发生有不同程度的烃代谢进化有关。A.sarcoidesNRRL 50072 基因组与烃类代谢相关的基因簇可高达80个,该菌株能产10多种烷烃类[9 ]。与NRRI 50072菌株产丰富的烃类物质比较,本文研究的6个菌株产有限种类的烃产物,这除了上面分析的遗传进化和基因表达不同外,也与培养条件密切相关[10]。最后,该属真菌成员在挥发性醇类的代谢上较之烃类的代谢更表现有保守性(表3),尤其在低级醇如乙醇、丙醇和丁醇上。6株菌株均产3-甲基-1-丁醇(产物编号17,表3),该产物几乎在每次的测定中均显著而稳定地被检测出;该产物也始终伴随在A.sarcoidesNRRL 50072的挥发性代谢物中[5,10],说明3-甲基-1-丁醇可能是该属真菌挥发性有机化合物的特征成分。
4 结论
通过研究在遗传和代谢上具有多样性的囊盘菌株所产生的挥发性有机化合物的组成,得出囊盘菌可能普遍具有产挥发性烃类的能力,但其产物种类具有菌株专化性。该发现为研究产挥发性烃类物质的真菌代谢途径提供了参考,也为发现或通过遗传工程创造符合目标的新菌株,进而利用该属真菌或其他真菌来发展生物燃料奠定了基础。
[1] Strobel G. The use of endophytic fungi for the conversion of agricultural wastes to hydrocarbons[J]. Biofuels, 2014, 5(4): 447-455.
[2]Strobel G. The story of mycodiesel[J]. Current Opinion in Microbiology, 2014, 19: 52-58.
[3]Song Z, Kennedy P G, Liew F J,etal. Fungal endophytes as priority colonizers initiating wood decomposition[J]. Functional Ecology, 2017, 31(2): 407-418.
[4]Suryanarayanan T S, Thirunavukkarasu N, Govindarajulu M B,etal. Fungal endophytes: an untapped source of biocatalysts[J]. Fungal Diversity, 2012, 54(1): 19-30.
[5]Strobel G A, Knighton B, Kluck K,etal. The production of myco-diesel hydrocarbons and their derivatives by the endophytic fungus Gliocladium roseum (NRRL 50072)[J]. Microbiology, 2008, 154(11): 3319-3328.
[6]Mallette N D, Knighton W B, Strobel G A,etal. Resolution of volatile fuel compound profiles from Ascocoryne sarcoides: a comparison by proton transfer reaction-mass spectrometryand solid phase microextraction gas chromatography-mass spectrometry[J]. AMB Express, 2012, 2, 23-35.
[7]Strobel G, Tomsheck A, Geary B,etal. Endophyte Strain NRRL 50072 producing volatile organics is a species of Ascocoryne[J]. Mycology, 2010, 1(3): 187-194.
[8]Roll-Hansen F, Roll-Hansen H. Microflora of sound-looking wood inPiceaabiesstems[J]. European Journal of Forest Pathology, 1979, 9(5): 308-316.
[9]Gianoulis T A, Griffin M A, Spakowicz D J,etal. Genomic Analysis of the Hydrocarbon-Producing, Cellulolytic, Endophytic FungusAscocorynesarcoides[J]. Plos Genetics, 2012, 8(3): e1002558.
[10]Mallette N, Pankratz E M, Parker A E,etal. Evaluation of Cellulose as a Substrate for Hydrocarbon Fuel Production byAscocorynesarcoides(NRRL 50072)[J]. Journal of Sustainable Bioenergy Systems, 2014, 4(1): 33-49.
[11]Aamir S, Sutar S, Singh S,etal. A rapid and efficient method of fungal genomic DNA extraction, suitable for PCR based molecular methods[J]. Plant Pathology & Quarantine, 2015, 5(2): 74-81.
(责任编辑:崔 贝)
Metabolites Diversity and Hydrocarbons Analysis on Volatile Organic Compounds Produced byAscocoryneStrains
GAO Qian1, YAN Dong-hui1, WANG Kai-ying1, LI Hong-chang1, REN Fei2
(1. Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China; 2. Forestry Experiment Center of North China, Chinese Academy of Forestry, Beijing 102300, China)
[Objective] Endophytic fungusAscocorynesarcoides NRRL 50072 was firstly found to be able to produce volatile compounds similar to those in fossil fuels. This could be a new avenue to biofuel development. The purpose of this study is to clarify whether the other strains from the same species or related species inAscocorynehave similar metabolisms to produce biofuel compounds. [Method] Different strains fromAscocorynewere isolated with tissue-surface sterilization. The evolutionary relationship between the strains in the genus was determined through constituting a ML phylogenetic tree. The compounds in volatile gas from the strains were analyzed and identified in NIST database with HSSPEM-GC-MS. [Result] All the strains studied were able to produce volatile metabolites containing hydrocarbons, alcohols, esters, aldehydes, acids, etc. However, the volatile compounds among the strains were highly diverse, even among closely related strains with significantly different compositions in their volatile compounds. [Conclusion] Most fungi fromAscocorynecould have the capability to produce volatile hydrocarbons. The compounds of hydrocarbons may be strain-specific. Therefore, it is fundamental for discovering or genetic engineering new strains to study and develop myco-diesel fromAscocorynefungi.
Ascocoryne;VOCs;SPEM-GC-MS;bioenergy;hydrocarbon
10.13275/j.cnki.lykxyj.2017.03.003
2016-10-16
水解纤维素产烃树木内生菌资源利用技术引进(2013-4-10)、效应物在杨盘二孢半活养方式及亲和寄主致病性中的作用(31370645)、芯片通量病原检测技术在杨树病害防控中的应用([2016]10)
高茜,博士,助理研究员,环境生态学,E-mail:gq8778@caf.ac.cn
* 通讯作者:严东辉,博士,研究员,主要从事森林病理与树木微生物学,E-mail:yandh@caf.ac.cn
S567.3
A
1001-1498(2017)03-0376-08
