云冷杉过伐林垂直结构特征分析
2017-06-19陈科屹张会儒雷相东娄明华
陈科屹,张会儒*,雷相东,娄明华,卢 军
(1.中国林业科学研究院资源信息研究所,北京 100091; 2.宁波市农业科学研究院,浙江 宁波 315040)
云冷杉过伐林垂直结构特征分析
陈科屹1,张会儒1*,雷相东1,娄明华2,卢 军1
(1.中国林业科学研究院资源信息研究所,北京 100091; 2.宁波市农业科学研究院,浙江 宁波 315040)
[目的]以云冷杉过伐林为研究对象,综合分析其在垂直层次上的结构特征,进一步探索云冷杉过伐林的生长过程、演替规律,更加准确和全面地认识云冷杉过伐林的状态和发展趋势,从而为合理地制定和调整针对过伐林的经营措施提供理论依据。 [方法]基于12块面积1hm2的标准地数据,首先,采用定量的树冠光竞争高法划分林层,将林分从垂直层次上划分为上林层、中林层和下林层;其次,在垂直结构的基础上,进行林分结构特征分析,包括各林层的直径分布、树种组成、树种隔离程度、竞争以及分布格局。[结果]表明:(1)以树冠光竞争高度法划分的林层分层效果较好,各林层树冠光竞争高、平均胸径和蓄积量均在0.01水平上差异极显著;(2)随林层高度的增加直径分布由反“J”形分布转变为左偏单峰状分布,中林层和上林层峰值差别较大;(3)各林层树种构成差异不大,但下林层中针叶树种蓄积比重偏低;(4)上林层更有可能出现较高的混交度,不同树种随林层高度的增加其混交度的变化趋势有所差异;(5)随林层高度的增加,林分平均竞争压力逐渐减小,各树种竞争压力随林层的变化趋势与各林层总体平均变化趋势基本一致;在不同林层,不同生长习性的树种竞争差异明显;(6)各林层多呈聚集和随机分布,上林层更有可能出现随机分布,而下林层更有可能出现聚集分布。[结论]综合分析可知研究区的林分结构相对稳定,处于由过伐林向原始林恢复演替的进程中。垂直结构定量化的方法能更加全面的认识和描述过伐林的结构现状及未来的发展方向,研究结果可为长白山地区过伐林的可持续经营提供参考。
云冷杉;过伐林;垂直结构;定量化分析;综合特征
长白山林区于20世纪30年代、解放前后以及20世纪60年代经过几次高强度采伐后,森林类型已由原来以红松-云杉-冷杉为主的原始林变成目前以云冷杉和部分阔叶树种为主的过伐林。这些森林已经失去了原始林整体的结构和功能,如何恢复其原有的结构,使其逐步发展为健康稳定的森林生态系统,保障森林的多种效益,实现可持续的利用是目前经营过伐林亟待解决的问题。通过合理的经营改造措施,过伐林较次生林更容易恢复到原始针阔混交林的状态[1]。但是,过伐林与其他森林类型相比具有鲜明的林分结构特点,需要据此制定针对性较强的经营改造措施才能实现森林结构和功能的恢复[2]。因此,分析掌握云冷杉过伐林的林分结构特征是科学制定合理经营措施的重要前提。
目前,在云冷杉过伐林结构特征分析方面已经开展了大量研究,这些研究主要集中在物种组成、直径结构分布、生物多样性、水平空间结构等方面[3-7]。缺乏从垂直结构方面,特别是垂直结构定量化方面综合分析云冷杉过伐林结构特征的相关研究。基于林分垂直层次结构展开林分结构特征综合分析,对于获取更完整合理的过伐林林分结构特征信息,从生态系统的角度指导森林经营具有重要意义[8-9]。此外,近年来有学者对部分林分结构指标进行了修正和改进,为林分结构定量化分析提供了更为合理的方法[10-11]。因此,本研究以云冷杉过伐林为研究对象,综合分析其在垂直层次上的结构特征,为进一步探索云冷杉过伐林的生长过程、演替规律,为合理地制定和调整针对过伐林的经营措施提供重要的理论依据。
1 研究区概况
研究区位于吉林省汪清林业局的金沟岭林场,属长白山系老爷岭山脉雪岭支脉,地理坐标为130°10′ E, 43°22′ N。林场东接荒沟林场,西靠塔子沟林场,南与十里坪林场接壤,北临地阴沟林场。林区地貌属低山丘陵,海拔在300~1 200 m之间,坡度主要集中在5°~25°,个别陡坡在35°以上。该区属季风型气候,1月份最低平均气温-32℃,7月份最高平均气温32℃,全年平均气温为4℃;年降水量600~700 mm,且多集中在7月份。本地区土壤多为灰棕壤土。研究区植被主要树种有鱼鳞云杉(PiceajezoensisCarr.)、红皮云杉(PiceakoraiensisNakai)、冷杉(Abiesnephrolepis(Trautv.) Maxim.)、落叶松(Larixgmelinii(Ruprecht) Kuzeneva.)、红松(PinuskoraiensisSiebold et Zuccarini.)、枫桦(BetulacostataTrautv.)、椴树(TiliaamurensisRupr.)、白桦(BetulaplatyphyllaSuk.)、榆树(UlmuspumilaLinn.)、杨树(PopulusussuriensisKom.)、色木(AcermonoMaxim.)、水曲柳(FraxinusmandschuricaRupr.)、与青楷槭(AcertegmentosumMaxim.)和花楷槭(AcerukurunduenseTrautv. et Mey.)等。
2 材料与方法
2.1 数据来源
在研究区设置12块面积为100 m×100 m的标准地,按相邻格子法将标准地划分成100个10 m×10 m的小样方,在每个小样方内检尺胸径5 cm以上所有林木的特征值,包括树种、胸径、树高、枝下高、冠幅等因子,并用皮尺测定每株林木的空间位置信息。各标准地基本信息见表1。为消除边缘效应,将标准地内距各边界5 m之内的范围确定为缓冲区,其余部分为核心区。核心区内的林木全部作为对象木计算空间结构参数,缓冲区内部的林木仅作为相邻木,不参与空间结构参数统计[12-13]。选择对象木周围最近的4株林木作为邻近木组建最小空间结构单元,据此计算相关空间结构参数[14-16]。

表1 标准地基本概况
2.2 分析方法
首先,对林分进行垂直结构层次划分,为避免传统定性划分方法的弊端,本次研究采用定量的树冠光竞争高法划分林层。其次,在垂直结构的基础上,再进行林分结构特征分析。除采用经典模型形式对直径分布特征进行拟合分析外,主要侧重于使用经过进一步改进完善后的空间结构参数,对林分的空间结构状态进行分析,分别是基于相邻木排列关系的混交度、基于交角的林木竞争指数和进行显著性检验的角尺度。
2.2.1 林层划分 本研究采用树冠光竞争高度(canopy competition height, CCH)的方法对云冷杉过伐林进行垂直空间分层划分,即遵循林分结构中不同高度层树叶在光合作用方面的贡献差异这一生态学现象,根据林分中林木的树高和冠长确定每一林层的高度值,再将林分中的每一株林木划入相应林层的方法和过程。具体采用以下计算公式确定树冠光竞争高度,即计算每层林层的分界值[17]。
CCH=a·CL+HW
(1)
式中:CCH为树冠光竞争高度;a为截止系数;CL为冠长;HW为枝下高。
根据学者们对林分分层的研究成果可知,通常将林分分为3~4层[18-19]。结合过伐林的垂直结构特点以及林分的现地观测情况,将研究区的林分分为3层,分别是上林层、中林层和下林层。考虑到林分内中上层林木的光照差异较大,而下层林木受光率普遍较低的特点,在学者们前期研究的基础上对树冠光竞争高度法作适当调整[17, 20-21]。具体方法如下:首先,找出树高最高冠长最长的一株林木计算其CCH1值,把该值作为上林层的最低高度分界点,将所有树高大于或等于CCH1的林木划入上林层;再以此类推,计算出CCH2值,将所有树高大于或等于CCH2的林木划入中林层;最后将树高低于CCH2值的所有林木划入下林层。式(1)中截止系数a的取值范围通常在0.3~0.5之间,结合数据处理情况,文中a取值为0.4。
2.2.2 直径分布拟合 根据各林层直径分布的曲线形式,选择适合的函数形式对直径分布进行拟合,找出其中拟合精度最高的函数用于描述不同林层的直径分布情况。常用的直径分布函数包括:Weibull函数、logistic函数、负指数函数、逆函数和多项式函数等[3, 22-24]。
2.2.3 树种隔离程度计算 本文采用娄明华等[25]提出的基于相邻木排列关系的混交度,描述树种间的相互隔离程度,计算公式为:
(2)
式中:n为空间结构单元中最近相邻木株数,vij为对象木i与最近相邻木j的树种异同数,不同时vij=1,反之,vij=0;vj,j+1为相邻最近相邻木的树种异同数,不同时vj,j+1=1,反之,vj,j+1=0,规定vn,n+1=vn,1;令uj=vj,j+1,当且仅当相邻两个uj值都等于零时,uj,j+1=0;反之,uj,j+1=1,规定un,n+1=un,1;ni为最近邻木的树种数,Mpi为林木点混交度,Mpi∈[0,1]。将Mpi的取值划分为0,(0,0.25 ],(0.25,0.5],(0.5,0.75],(0.75,1]5个区间,分别对应零度、弱度、中度、强度和极强度混交5个隔离程度。
2.2.4 林木竞争状况计算 本文采用惠刚盈等[26]提出的基于交角的林木竞争指数,描述林木个体承受压力的大小,计算公式为:
(3)
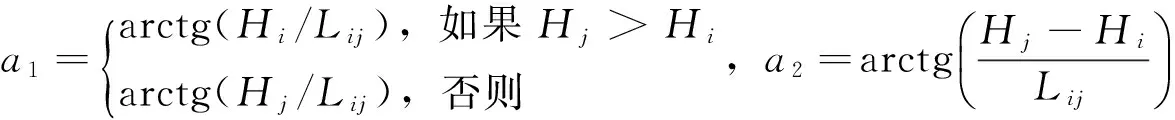
其中,CIH为竞争指数,Hi为对象木的树高,Hj为相邻木的树高,Lij为对象木与相邻木之间的距离,n为结构单元内相邻木的株数。CIH∈[0,1),数值越大,表明林木所承受的竞争压力越大。
2.2.5 林木空间分布格局计算 首先采用惠刚盈等[27]提出的角尺度法计算对象木的角尺度值,再用赵中华等[28]提出的显著性检验方法判断林木的水平分布格局。角尺度计算公式为:
(4)
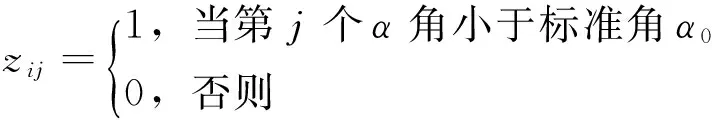
显著性检验的计算公式为:
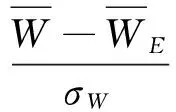
(5)
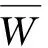
3 结果与分析
3.1 分层情况
各标准地分层情况见表2。上林层的树冠光竞争高度值分布范围在15.4~23.4 m之间,平均胸径在21.9~27.1 cm之间,蓄积量比重在31.8%~84.8%之间;中林层在上述统计量的分布范围分别是:7.4~13.6 m,11.7~18.5 cm,14.0%~60.9%;下林层各统计量分布范围是:2.4~4.1 m,7.0~9.4 cm,1.2%~8.4%。各林层在树冠光竞争高度的均值分别为19.4 m、10.9 m、3.2 m;平均胸径的均值分别为24.4 cm、15.5 cm、8.1 cm;蓄积量比重的均值分别为64.7%、31.0%、4.3%。将各统计量进行方差分析,结果见表3。由F检验结果可知,不同林层间统计量值在0.01水平上呈现极显著差异。各林层之间的统计量特征值差异极显著,表明林分分层效果较好。

表2 各标准地不同林层的基本情况

表3 各林层基本情况的方差分析
3.2 直径分布和树种组成

图1 不同林层直径分布情况Fig.1 Diameter class distribution of each storey
林分水平上各林层的直径分布见图1。从整体上看,由各林层直径分布曲线的峰值变化可以得知,随着林层高度的降低,林木的径阶随之变小,而林木的株数则随之增加。下林层的径阶分布呈现典型的反“J”分布,即随着径阶的增大,林木株数呈现急剧减少的趋势,达到一定直径后,株数减小幅度逐渐趋于平缓,其峰值在6径阶。中林层和上林层的径阶分布表现为左偏单峰状分布,峰值分别在14径阶和24径阶处。从各林层径阶分布的范围来看,上林层最宽、中林层次之、下林层最窄。用2.2.2节列举的多种函数进行曲线拟合后发现,Logistic方程:F=1/(1+ea-bx),能很好的拟合林分株数与株数累计频率值之间的函数关系,可将其作为描述各林层直径的分布情况的函数,拟合结果见表4。

表4 各林层直径分布Logistic方程参数拟合情况
林分水平上各林层的树种组成情况见表5。耐阴性较强的针叶树种在各林层皆占有一定的比重,且在上林层内,蓄积量最靠前的5个树种中,针叶树种占了4位,这类林分的林分结构通常较为稳定。但随着林层的下降,针叶树在其中的所占位数逐渐下降到3位、2位。从针阔比例上也可看出,随林层的下降,阔叶树在林分中的比重在整体上升。上林层是受当年高强度采伐的直接影响生长发育起来的林分,原始林中原本蓄积比重最大的云杉、冷杉和红松已经降到落叶松和枫桦之后。采伐后形成的大片林窗为喜光、生长较快的阔叶树种提供了适宜的生长环境,再加上针叶树种被大量采伐,严重阻碍了针叶树种的天然下种更新,进一步导致了针叶树种蓄积比重的下降。特别是在下林层中,云杉在该林层的比重已明显偏低,不利于原始针阔混交林结构的恢复。

表5 各林层树种组成情况
3.3 树种隔离程度

图2 各标准地各林层的平均混交度Fig.2 The mean mingling of each storey of each sample plots
各标准地不同林层平均混交度情况见图2。由图可知,各标准地不同林层的平均混交度均在0.5以上,林分总平均混交度为0.644(表6),处于强度混交状态,混交状况较好。标准地不同林层的平均混交度变化情况较为复杂,没有呈现明显的变化趋势。上、中、下林层的平均混交度分别为0.648、0.633、0.655,从整体上看差异极小。但从个别标准地不同林层的平均混交度变化情况以及林分水平上不同林层平均混交度的均值的细微差异可以看出,在过伐林林分内,下林层拥有最大平均混交度的可能性较高,这可能是由于采伐过后,过伐林内留下的面积较大的林窗造成的。面积较大的林窗为天然更新提供了有利的场所,林窗内部充足的生长空间使得混交林中的各类树种在幼龄阶段能够共同生长,因此这部分下林层林木能够保持较高的混交度水平,从而在一定程度上抬高了下林层的平均混交度。随着林木的生长,各林木间竞争压力的增大,林分中的某一类树种开始被分化和稀疏,使得下一生长阶段林分的混交度出现下降。在上林层和中林层内出现了较多0度混交的林木也能够应证上述情况。此外,由数据结果可以明确的一点是,该过伐林森林群落处于恢复演替期,林分结构随林木的生长会有较大幅度的变化。
各树种在不同林层的平均混交度情况见表6。除下林层中的落叶松外,各树种在各林层的平均混交度均在0.5以上,混交情况整体较好。白桦、色木、杨树在下林层中的平均混交度较低,随着林层高度的增加,平均混交度也出现增加。这是由于这类林木在更新时多聚集生长,导致其在幼龄阶段混交度较低;而在上层林中这类树种又不占优势,混交生长在各类针叶树种之间,导致其平均混交度变高。云杉、冷杉、红松在下林层中的平均混交度较高,随着林层高度的增高,平均混交度出现降低。这是由于此类暗叶树种能够在一定的郁闭环境下更新,因此生长发育出的小树在林分内较为分散,混交度较高;在上林层中,该类树种为主要树种,在林分中的相对比重大,因此混交度低。林分中红豆杉和黄菠萝的株数均极少,与周围林木皆不相同,但红豆杉在不同林层均具有较高的混交度,而黄菠萝在上林层混交度却不高,这是由于两者各自空间结构单元内邻近木之间的多样性差异和排列关系不同造成的。

表6 各林层不同树种的平均混交度
3.4 林木竞争状况
在标准地水平上看,上、中、下林层的平均竞争指数分别在0.078~0.149、0.217~0.438、0.524~0.636之间,中林层是下林层的2.2~3.5倍,上林层是中林层的1.5~2.4倍。在林分水平上,上、中、下林层平均竞争指数分别为0.112、0.307、0.571,中林层是上林层的2.8倍,下林层是中林层的1.9倍。结合图3可以直观地看出,随着林层高度的增加,林层平均竞争指数呈现逐级下降,且降幅较为明显。降幅最大的是12号标准地,最终降幅高达87%;降幅最小的是10号标准地,降幅也达到了77%。由于上林层多为优势木,其所处的竞争压力较小。
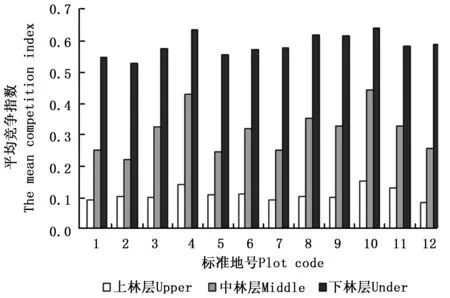
图3 各标准地各林层的平均竞争指数Fig.3 The mean competition index of each storey of each sample plots
各树种在不同林层的平均竞争指数见表7。除红豆杉和杨树在中林层表现出异常外,其余树种均呈现出随林层高度的增加,平均竞争指数逐级减小的趋势,说明上林层内的林木竞争压力最小。红豆杉的数量极少,在中林层内仅2株,不能代表树种的平均变化水平。黄菠萝在林分中的情况与红豆杉相似,也不能代表该树种的平均水平。杨树在中林层内的平均竞争指数较在下林层的平均竞争指数大,这可能是由于局部特殊情况造成的。杨树属于喜光先锋树种,在林下较阴暗环境中生长的杨树受到抑制,而杨树周围的针叶树种具有较强的耐阴性,生长阻力相对较小,这导致杨树在空间结构单元内的交角较大,导致竞争指数增加。但是,从整个中林层的内部情况来看,杨树的平均竞争指数是所有树种中最小的,说明杨树的整体竞争压力是最小的。过伐林受强度采伐影响,整体郁闭程度并不高,给杨树提供了较好的生长空间。这说明在中林层,杨树平均竞争指数出现的异常情况是由于在局部环境内生长的部分杨树造成的,具有一定的特殊性。在上林层中,平均竞争指数最小的是生长较慢的红松,最大的是生长较快的杂木;而在下林层中则相反,平均竞争指数最小的是生长较快的杨树,最大的是生长较慢的水曲柳。这是由于慢生树种在前期高生长过程中不如速生树种快,常常被速生树种的冠幅遮盖或部分遮盖,导致其在下林层中竞争压力较大。在上林层中则相反,耐阴的慢生树种慢慢成为主林层后不利的光照环境影响了喜光速生树种的生长,致使速生树种的竞争压力较耐阴较慢生树种大。

表7 各林层不同树种的平均竞争指数
3.5 林木空间分布格局
各标准地不同林层的林木分布情况见表8。可以看出,标准地内各林层处于聚集分布和随机分布状态,没有林层呈现均匀分布状态。3、6、8号标准地中各林层均呈聚集分布状态,10、12号标准地中各林层均呈随机分布。其余各标准地内林层之间分布状态有所差异,没有表现出明显的规律性。随机分布状态在各林层中均有可能出现,但出现在上层林中的可能性最大,而多数下林层呈聚集分布状态。这是因为更新的部分林木由于生理特性的要求和林分环境的约束,在幼龄阶段多聚集生长;随着竞争压力增大,林木开始出现分化,在自然稀疏过程中向着随机分布状态过渡。但在少部分标准地内,在上林层中也出现了聚集分布的情况,这可能是由于采伐活动多发生在上层林中,而采伐木的位置又具有较强的不确定性,这导致部分标准地上林层内的林木空间分布格局被改变。

表8 各标准地各林层分布格局判断结果
林分水平上不同林层角尺度分布频率见表9。可以看出,各林层中处于随机分布状态的林木占林木总株数的一半以上,其中上林层比重最大,为58.2%。这与上述数据体现出的随机分布状态更有可能出现在上林层的结果保持一致。与下林层相比,除角尺度取值在1处的比重较低以外,其余取值条件下比重均比下林层高。这说明下层林中处于聚集分布的林木较多,随着林木的生长分化,林木有向随机和均匀分布状态转移的可能。中林层与下林层相比,角尺度取值在0和1处的比重没有任何变化,仅在0.25、0.5、0.75三处呈现局部调整,表现出由下林层向上林层转变的一种演替或过渡状态。

表9 各林层平均角尺度频率分布
4 讨论
从演替的角度来看,下林层的林木会遵循一定的生长规律逐渐过渡到中林层,最终成为上林层。在理想情况下,下林层的林分结构决定了林分结构未来的演替方向。而上林层的林分结构状况正是下林层林分结构形成的基础,并为下林层林木的生长提供了生长场所。其次,上林层还会对整个林分的树种组成和蓄积量等结构特征产生关键性的影响。中林层在更多时候起到的是一种过渡和保障作用。由此可见,在整个林分中,各个林层具有各自的功能和作用。此外,过伐林由于受到人类活动的强烈干预,林分结构与其他类型的林分差异极大。因此,在对过伐林开展经营活动时,需要针对过伐林各林层的现状和可能的演替方向,以及各林层在林分中的功能和作用,制定综合性的经营措施。
从文中数据分析的结果来看,该过伐林处于恢复演替期,距离原始林的顶级林分结构还有较大的差距,随着时间的推移各林层的林分结构还会发生较大幅度的变化。因此,对于云冷杉过伐林林分综合结构特征的研究应该是一个长期的过程,探索不同林层在时间维度上的动态变化是完善林分结构特征研究的一个重要方向。
5 结论
研究区云冷杉过伐林保留了原始林的部分结构特征,垂直结构层次分明,上、中、下林层平均树冠光竞争高分别为19.4 m、10.9 m、3.2 m,胸径的平均值分别为24.4 cm、15.5 cm、8.1 cm;蓄积量比重的平均值分别为64.7%、31.0%、4.3%。各林层直径分布差异明显,下林层呈反“J”形分布,中林层和上林层呈左偏单峰形分布,但峰值差异较大。用Logistic方程能较好的描述各林层的直径分布结构。
林分水平上各林层树种构成相对一致,且耐阴性较强的树种在各林层皆占有一定比重,林分整体结构相对稳定。各树种的比重在不同林层有一定的差异。上林层的树种组成情况对整个林分树种组成情况的影响至关重要,仍以耐荫的针叶树为主。下林层中针叶树种比重偏低,特别是云杉,需要采取适当的人工干预措施。枫桦、白桦、杨树等阳性先锋树种在一定时期内仍将保持一定的比重,但由于在下林层中更新不良,在演替的过程中其优势度会逐渐下降。
林分混交水平整体较高,各标准地内各林层平均混交度差异较小,上林层出现较大混交度的可能性较大。各个树种在不同林层的整体混交度水平皆较高,不同的树种由于生理特性和生长环境差异在林层之间表现出一定的规律性变化。白桦、色木、杨树在林分中的平均混交度随林层的增高逐渐增加,而云杉、冷杉、红松在林分中的平均混交度随林层的增高逐渐降低。
在林分水平上,不同林层的竞争压力随林层高度的降低逐渐增大,且增长幅度十分明显。从树种的角度来看,除杨树以外,各树种的竞争压力随林层的变化趋势与林分水平上表现的变化趋势一致。杨树表现出的异常情况是由于局部环境造成的影响,不具有普遍性。从林层内部看,在下层林内,竞争压力较大的更有可能是耐阴性慢生树种,竞争压力较小的更有可能是喜光性速生树种;上林层中的情况则与之相反。
聚集分布和随机分布是各林层常见的分布状态,符合天然林的特征。上林层出现随机分布的可能性较大,下林层多呈聚集分布。采伐活动会直接影响上林层和间接影响下林层的林木空间分布格局。从林分水平上林木角尺度的分布频率来看,各个林层的大部分林木均处于随机分布状态,其中上林层的比重最大。下林层处于聚集分布的林木较多。中林层是处于上林层和下林层之间的一种过渡状态。
综合分析林分的空间结构信息可以看出,该林分的林分结构会在一定时期内保持相对稳定。结合历史研究资料得知,该林分的结构现状还远未达到研究区原有的合理林分结构状态,处于经强烈干扰后恢复演替的进程当中。
[1] 亢新刚, 赵俊卉, 刘 燕. 长白山云冷杉针阔混交过伐林优化结构研究[J]. 林业资源管理, 2008(03):57-62.
[2] 田树华, 邸宝良. 长白山过伐林经营[M]. 延吉: 延边大学出版社, 1994.
[3] 胡文力. 长白山过伐林区云冷杉针阔混交林林分结构的研究[D]. 北京:北京林业大学, 2003.
[4] 乌吉斯古楞, 王俊峰, 郑小贤, 等. 金沟岭林场过伐林更新幼苗空间结构分析[J]. 中南林业科技大学学报, 2009,29(04):21-25.
[5] 张会儒, 武纪成, 杨洪波, 等. 长白落叶松-云杉-冷杉混交林林分空间结构分析[J]. 浙江林学院学报, 2009,26(03):319-325.
[6] 曾祥谓, 玉 宝, 乌吉斯古楞, 等. 云冷杉过伐林主要树种结构特征分析[J]. 林业科学研究, 2014,27(04):481-486.
[7] 陈亚南, 杨 华, 马士友, 等. 长白山2种针阔混交林空间结构多样性研究[J]. 北京林业大学学报, 2015,37(12):48-58.
[8] Franklin J F, Pelt R V. Spatial aspects of structural complexity in old-growth forests[J]. Journal of Forestry, 2004, 102(3): 22-28.
[9] Ruprecht H, Dhar A, Aigner B,etal. Structural diversity of English yew (TaxusbaccataL.) populations[J]. European Journal of Forest Research, 2010, 129(2): 189-198.
[10] 汤孟平. 森林空间结构研究现状与发展趋势[J]. 林业科学, 2010,46(1):117-122.
[11] 惠刚盈. 基于相邻木关系的林分空间结构参数应用研究[J]. 北京林业大学学报, 2013,35(4):1-9.
[12] Arne P. Evaluating structural indices by reversing forest structural analysis[J]. Forest Ecology and Management, 2006, 224(3):266-277.
[13] 汤孟平. 森林空间经营理论与实践[M]. 北京:中国林业出版社, 2007.
[14] 惠刚盈, 克劳斯, 冯佳多. 森林空间结构量化分析方法[J]. 北京: 中国科学技术出版社, 2003.
[15] Hui G, Pommerening A. Analysing tree species and size diversity patterns in multi-species uneven-aged forests of Northern China[J]. Forest Ecology and Management, 2014, 316(2): 125-138.
[16] Graz F P. The behaviour of the measure of surround in relation to the diameter and spatial structure of a forest stand[J]. European Journal of Forest Research, 2008, 127(2): 165-171.
[17] Latham P A, Zuuring H R, Coble D W. A method for quantifying vertical forest structure[J]. Forest Ecology and Management, 1998,104(1):157-170.
[18] 张 健, 郝占庆, 宋 波, 等. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J]. 应用生态学报, 2007,18(08):1681-1687.
[19] 李明辉, 何风华, 潘存德. 天山云杉天然林不同林层的空间格局和空间关联性[J]. 生态学报, 2011,31(3):620-628.
[20] 郑景明, 张春雨, 周金星, 等. 云蒙山典型森林群落垂直结构研究[J]. 林业科学研究, 2007,20(6):768-774.
[21] 玉 宝, 张秋良, 王立明. 中幼龄兴安落叶松过伐林垂直结构综合特征[J]. 林业科学, 2015,51(1):132-139.
[22] 惠刚盈, 盛炜彤. 林分直径结构模型的研究[J]. 林业科学研究, 1995,8(2):127-131.
[23] 亢新刚, 胡文力, 董景林, 等. 过伐林区检查法经营针阔混交林林分结构动态[J]. 北京林业大学学报, 2003,25(6):1-5.
[24] 乌吉斯古楞. 长白山过伐林区云冷杉针叶混交林经营模式研究[D]. 北京:北京林业大学, 2010.
[25] 娄明华, 汤孟平, 仇建习,等. 基于相邻木排列关系的混交度研究[J]. 生态学报, 2012,32(24):7774-7780.
[26] 惠刚盈, 胡艳波, 赵中华, 等. 基于交角的林木竞争指数[J]. 林业科学, 2013,49(6):68-73.
[27] 惠刚盈, K. V. Gadow, 胡艳波. 林分空间结构参数角尺度的标准角选择[J]. 林业科学研究, 2004,17(6):687-692.
[28] 赵中华, 惠刚盈, 胡艳波, 等. 角尺度判断林木水平分布格局的新方法[J]. 林业科学, 2016,52(2):10-16.
(责任编辑:彭南轩)
Analysis of Vertical Structure Characteristics for Spruce-Fir Over-cutting Forest
CHEN Ke-yi1, ZHANG Hui-ru1, LEI Xiang-dong1, LOU Ming-hua2, LU Jun1
(1. Research Institute of Forest Resources Information Techniques, Chinese Academy of Forestry, Beijing 100091, China; 2. Ningbo Academy of Agricultural Sciences, Ningbo 315040, Zhejiang, China)
[Objective]To further understand the status and development tendency of spruce-fir over-cutting forest more by quantitative analysis of the vertical structure characteristics for spruce-fir over-cutting forest in Jin’gouling Forest Farm of Jilin Province.[Method]Based on the 12 Spruce-fir over-cutting forest sample plots with area of 1 hm2, the stands were divided into upper, middle and lower layers according to the canopy competition height (CCH) in stand. Each layer was used to analyze the distribution of diameter at breast height (DBH), the species composition, the segregation, the competition and the distribution pattern by the investigation data. [Result] (1) The effect of the CCH method is reasonable. The CCH, the mean DBH and the volume showed extremely significant difference at 0.01 levels. (2) The diameter distribution shifted from the reverse "J" shape distribution to left-skewed unimodal distribution with the increase of the height of storey. The difference of peaks between upper and middle layer was great. (3) The difference of tree species composition in each layer was little, but the proportion of volume of the coniferous species in the lower layer was on the low side. (4) The upper layer was more likely to be higher on the mingling, and the variation tendency of mingling was different with the increase of the forest storey for different tree species. (5) The average competition pressure of the forest storey decreased gradually with the increase of the forest storey, and the variation tendency of the competition pressures of the tree species with the changes of forest stories was basically consistent with the overall average change trend. But in different forest storeys, the species with different growth habits had different competitiveness. (6) Most of the forest storey followed the aggregative and randomly distribution, the upper layer was more likely to occur in a random distribution, and the lower layer was more likely to occur in an aggregation distribution. [Conclusion] Based on the comprehensive analysis, it is find that the stand is still in the progress of restoration succession. By this method it is enable for us to more comprehensively understand and describe the status and future development direction of the over-cutting forest structure, which can provide a reference for the sustainable management of over-cutting forest in Changbaishan Mountain area of China.
spruce-fir stands; over-cutting forest; vertical structure; quantitative analysis; synthetic characteristics
10.13275/j.cnki.lykxyj.2017.03.013
2016-08-25
“十二五”国家科技支撑计划项目(2012BAD22B02)
陈科屹(1989—),男,博士研究生,主要研究方向:森林可持续经营。E-mail:Lowrychen@sina.com
* 通讯作者:张会儒(1964—),男,研究员,博导,主要研究方向:森林可持续经营等。E-mail:huiru@caf.ac.cn.
S718.54
A
1001-1498(2017)03-450-10
