亚热带常绿阔叶林常见树种木质部密度与生长速率的种内种间变异研究
2017-06-15张令珍郭志文郑景明
张令珍, 郭志文, 郑景明
(北京林业大学林学院,北京 100083)
亚热带常绿阔叶林常见树种木质部密度与生长速率的种内种间变异研究
张令珍, 郭志文, 郑景明
(北京林业大学林学院,北京 100083)
测定了亚热带常绿阔叶林的13种常见乔木树种的木质部密度、胸径生长速率和树高、冠幅等形态指标,探讨种内种间水平上各性状的变异程度,以及在物种和个体水平上各性状与生长速率的相关性.结果表明:(1)木质部密度与生长速率种内变异大于种间变异;(2)在种间水平上树高、胸径、冠幅以及木质部密度与生长速率显著相关,水平木质部密度与生长速率显著负相关,仅分别有2个树种的树高和胸径与生长速率显著相关,冠幅在个体水平上与生长速率没有显著相关性;(3)在种内、种间水平上木质部密度与生长速率均有显著相关性,且两者的相关性不受个体大小的影响.
亚热带常绿阔叶林; 种间变异; 种内变异; 生长速率
森林群落的建成及其动态变化受群落中种群大小和结构的影响[1].在特定环境中,各物种通过性状的变异适应环境,响应各种生物、非生物因子的作用,从而形成群落中物种共存及动态机制.植物功能性状是指对植物体定植、存活、生长以及死亡存在显著影响的一系列植物属性,而且这些属性能够单独或者组合指示生态系统对环境的响应,并且能够对生态系统过程产生强烈影响[2].它们是植物在长期进化过程中,以一定的生态策略或属性适应周围环境的结果[3].不同的植物功能性状与植物体代谢过程的特定阶段以及生态系统的特定功能密切相关,例如,木质部密度和叶片干物质与植物的生长速率以及生态系统的物质循环显著相关,种子的大小影响植物体的定植、扩散能力,树高、冠幅、比叶面积和冠层结构与植物邻体之间的竞争能力有关,木质部密度、树体结构和种子大小与植物体的耐受能力密切相关[4].
在基于性状的生态学(trait-based ecology)研究中,物种间功能性状的变异被认为是群落中物种共存的前提[4].然而,不同物种的个体是群落构成的核心,因此功能性状的变异研究也应该包括以物种个体为研究对象的变异,但是由于取样、研究尺度等原因,物种的功能性状通常是以该物种的平均值来反映不同环境梯度或者干扰下功能性状的变异,而对种内水平上功能性状变异研究甚少[5].越来越多的研究表明种内功能变异以及遗传多样能够显著影响生物群落动态变化以及生态系统功能[6].种内功能性状变异反映了一个物种在生物以及非生物梯度上的最大适应能力,本质上决定了该物种的生态位宽度[7-8],影响群落的建成及稳定性,对于自然选择以及新物种形成的基础过程具有重要意义[9].周刘丽等[10]研究浙江天童常绿阔叶林主要树种的功能性状种内变异的大小、来源以及空间尺度相关性,发现种内变异程度依赖于选取的性状和物种,杨建军等[11]在研究高山栲叶性状种内变异与环境因子的关系时发现,叶性状种内存在较大的变异,且环境条件是影响叶性状种内变异的主要因素,唐青青等[3]研究表明,常绿和落叶树种的不同性状种内种间变异贡献率均有差异,但国内关于种内性状变异研究多集中在叶的功能性状,如比叶面积、干物质含量,但对树干的功能性状,如木质部密度、生长速率的研究报道相对较少.
同一群落中不同物种因遗传特征不同,适应光等生境因子的生活史对策的差异,其功能性状如木质部密度和生长速率存在明显差异[12];同一物种则由于个体生长发育阶段、邻域竞争、物理小环境差异等的存在,其功能性状会表现出种内变异[13].此外,不但同一功能性状在种间、种内差异的程度会有所不同,不同功能性状之间的关系也可能在种内、种间水平上发生变化,例如,木质部密度与叶干物质量在种间水平上为显著相关,但在种内水平上则相关不显著[14].但此类研究尚比较少见.本文主要研究中亚热带常绿阔叶林中常见乔木种的木质部密度和生长速率两个重要功能性状在种内种间的变异特性,以及两者与树高等表型性状在种内种间水平上的相关性.由于调查对象为来自同一群落环境条件下的不同物种和不同大小的个体,具体研究:(1)木质部密度和生长速率的种内、种间变异程度;(2)两者之间的相关性在种内、种间水平是否一致以及个体大小对两者之间的相关关系是否有影响.
1 材料与方法
1.1 研究区概况
研究区位于江西大岗山森林生态系统国家野外科学观测研究站,东经114°30′—114°45′,北纬27°30′—27°50′,该区属亚热带湿润气候区,年平均气温在15.8 ℃,年均日照长度为1 656.9 h,年均降水量1 590.9 mm,其中4-6月份降雨量占全年的45% ,降水主要集中在5-7月,年蒸发量为1 503 mm, 无霜期269 d.本区属地带性低山丘陵红壤、黄壤类型及其亚类的分布区,黄壤分布最广,分布海拔为 300~700 m ,红壤多分布于海拔200 m以下的低山丘陵.天然常绿阔叶林是中亚热带的地带性植被,也是本地区天然林演替系列的顶级群落,但由于长期严重的人为干扰,地带性植被已破坏殆尽,现有的植被类型主要是:天然次生常绿阔叶林、落叶阔叶林、各类针阔混交林和毛竹林,常见树种有丝栗栲(Castanopsisfargesii)、苦槠(Castanopsissclerophylla)、刨花楠(Machiluspauhoi)[15]等.
1.2 取样方法与样品收集
2015年6-8月,在江西大岗山念珠林场的丝栗栲—刨花楠次生天然林中设置了1 km2的固定标准实验样地,对样地内DBH≥1 cm的木本植物进行标记、编号、每木检尺、坐标定位.确定样地内有30种乔木树种,计算各树种的重要值,选择重要值前20位的乔木树种,在固定样地周围4个方向上设立4个30 m×30 m临时样地,对其中目标树种用生长锥1.3m树高处相互垂直两方向钻取两根年轮条,并记录胸径、树高、冠幅等.在实验室内将年轮条固定在特制的木槽上,自然晾干后分别用800、600、360和120目的砂纸打磨,直至年轮清晰可见.用WinDENRO年轮宽度测定分析系统测定各年轮条的年轮宽度,以近十年内生长量的平均值作为生长速率.用排水法测定各年轮条的体积(V),在烘箱烘干后称重(G),年轮条密度D=G/V,取每个物种的所有年轮条的密度作为该物种的木质部密度值.共测定430株林木(860根年轮条)的生长量及木质部密度数据.因部分树种年轮不清晰,最后选定13个树种的胸径生长速率及木质部密度值进行分析(表1).
1.3 数据分析
在上述数据的基础上,确定每棵树近十年生长量求取平均值作为该林木个体的年平均生长量.计算不同水平的变异系数CV=(标准差SD÷平均值M)×100%,总体水平的变异系数用所有个体实测值计算,种间变异系数用每物种的平均值计算,种内变异用每物种个体实测值计算.利用相关分析分别检验不同水平上生长速率与木质部密度分别与其他性状的相关性.种间相关性用每个物种性状平均值分析,种内相关则是用每种个体实测值进行分析.为研究树木个体大小是否会影响物种间水平的相关性,以胸径为协变量进行偏相关分析.
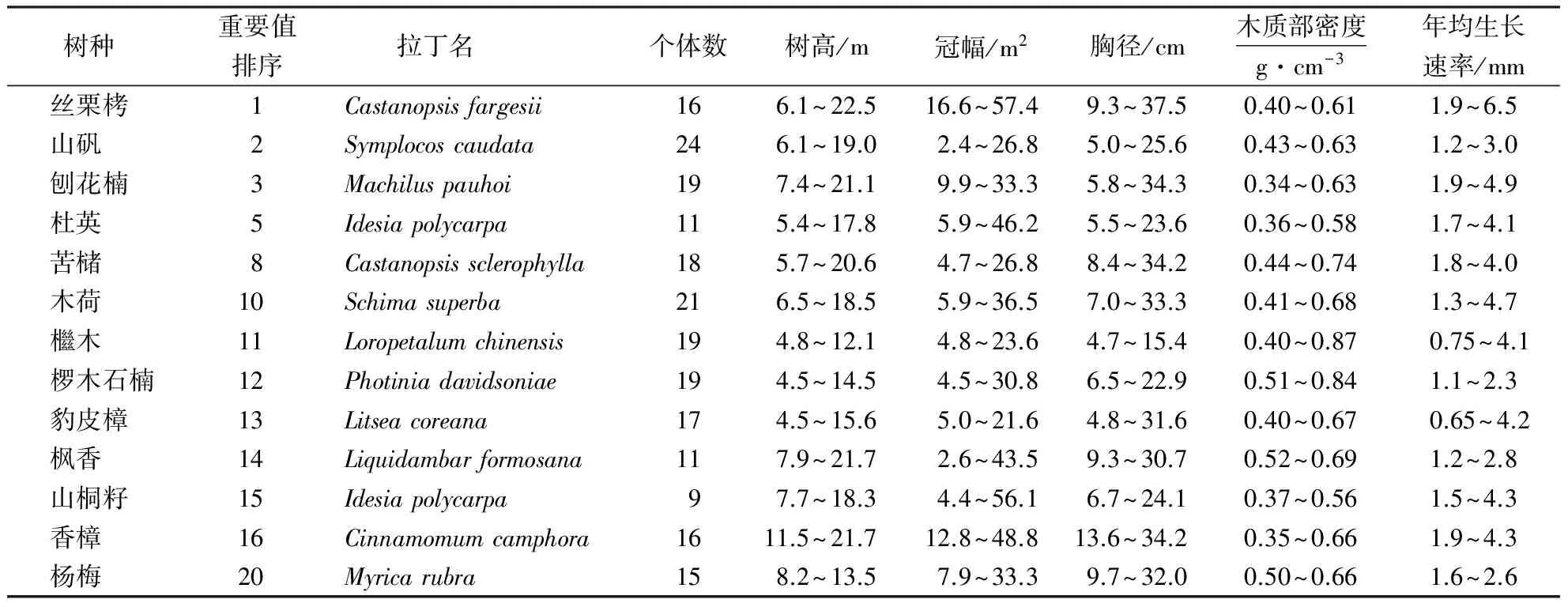
表1 各物种测定的性状及变异范围Table 1 Traits and their value range for 13 species
采用R软件(3.2.5)的ggm和psych程序包完成统计分析,显著度设定为P<0.05.
2 结果与分析
2.1 不同水平木质部密度和生长速率变异性
如图1所示,分别在物种和个体水平上研究各性状(种内变异、种间变异)的变异程度,在所测的树高、胸径、冠幅、木质部密度以及平均生长速率等功能性状中,木质部密度和生长速率的种内变异显著大于种间变异,且木质部密度无论是在种内水平还是种间水平,变异系数都是所有性状中最小的,说明木质部密度是相对稳定的功能性状.

a,b表示种内种间变异系数之间差异显著(P<0.05).图1 不同水平各性状变异系数比较Fig.1 Comparison of traits variation at different levels
2.2 种内、种间生长速率与各性状的相关性分析
以各物种性状平均值研究生长速率与各性状的种间相关性,结果表明,近十年生长速率与树高、胸径、冠幅以及木质部密度等都有显著相关性(P<0.05),以胸径为协变量研究偏相关关系,树高和冠幅与平均生长速率没有显著的偏相关关系,形态指标与生长速率的相关性受胸径大小的影响,而木质部密度与平均生长速率偏相关关系显著(表2),说明木质部密度与生长速率间的相关性不受个体大小的影响.
分别分析每个物种的生长速率与各因子的种内相关性.在13个物种中,生长速率与木质部密度表现出较强的相关性,6个树种的生长速率与木质部密度显著相关(P<0.05),胸径和树高分别有两个种与平均生长速率有显著相关关系(P<0.05)(表3).
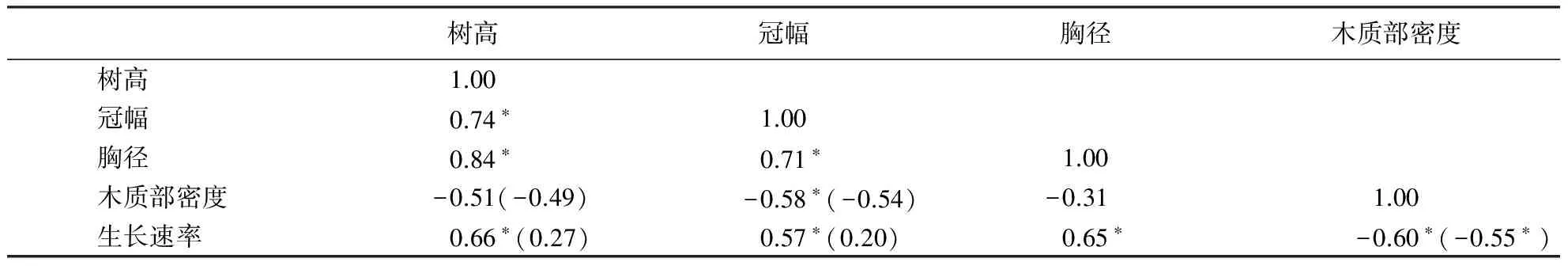
表2 生长速率与各性状种间相关性1)Table 2 Interspecific correlation between growth rate and other traits
1)括号内是以胸径为协变量的偏相关分析,*表示P<0.05.
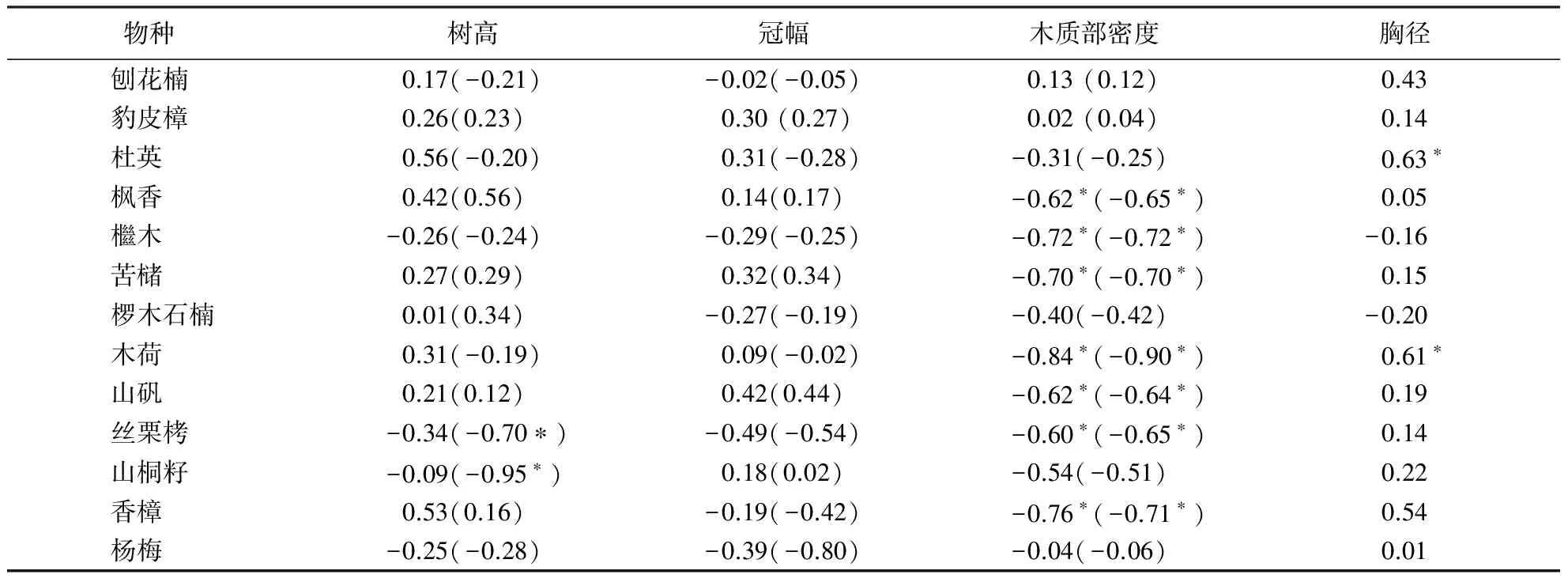
表3 生长速率与各性状种内相关分析1)Table 3 Intraspecific correlation between growth rate and other traits
1)括号内是以胸径为协变量的偏相关系数,*表示P<0.05.
2.3 木质部密度在种内种间相关性分析
在物种水平,木质部密度与其他因子的相关性分析中,冠幅与木质部密度有显著种间相关性(P<0.05),木质部密度与生长速率种间也有显著相关性(P<0.05),见表4;以每物种内所有个体的实测值计算种内相关系数,木质部密度与树高、冠幅以及胸径的种内相关关系均不显著(P>0.05),其中只有香樟的冠幅与木质部密度有着显著相关关系(P<0.05),杨梅的胸径与木质部密度有显著相关关系(P<0.05);以胸径为协变量研究个体大小对各性状之间相关性的影响,发现个体大小对木质部密度与其他性状的相关性影响较小,只有杜英和香樟的冠幅与木质部密度有显著相关性(P<0.05).
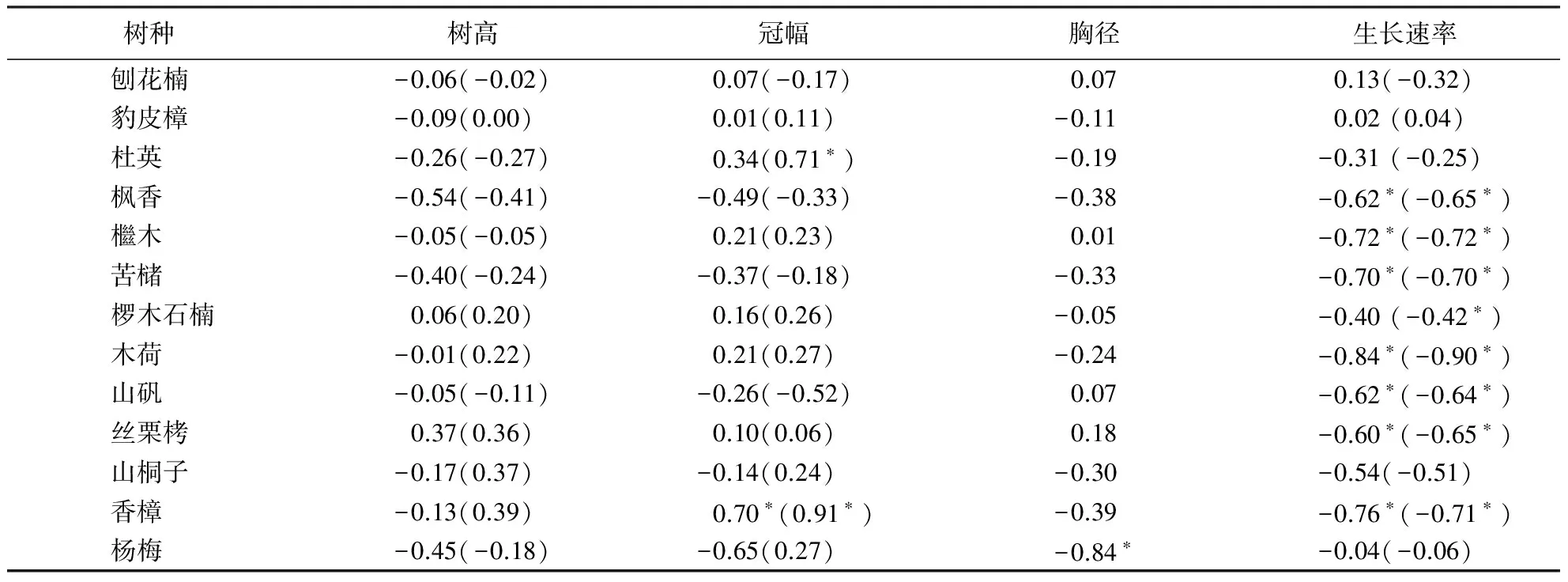
表4 木质部密度与各因子种内相关性分析1)Table 4 Intraspecific correlation between wood density and other traits
1)括号内是以胸径为协变量的偏相关系数,*表示P<0.05.
3 结论与讨论
3.1 各性状不同种内、种间水平的变异
植物功能性状变异对于群落的建成以及各物种的共存竞争有着非常重要的生态学意义[16].群落构建理论表明生境和生物竞争作用影响植物性状值的变异分布[17].生境过滤作用筛选具有相同性状的植物从而减小性状值得变化范围,而资源竞争导致生态位分化,使得性状值变异程度增大.种内变异在一定程度上反映了物种基因多样性和表型多态性,以及表明该物种表型可塑性强弱[18].本文研究结果木质部密度和生长速率种内变异大于种间变异,生境过滤作用使得具有相似功能性状的植物在同一生境中共存,而资源有限性使得这些物种通过种内变异呈现不同的生态策略,从而减少生物竞争强度,这是相似生境下生境过滤与生物竞争共同作用的结果.在以往的研究中人们主要把焦点放在种间变异上,忽略了种内变异在群落构建中的重要性,种内变异是物种生态可塑性以及生态位宽度的重要部分,是预测植物动态变化的重要内容[19].同时不同性状在种内种间分布有所差异,唐青青等[3]研究发现叶面积变异主要来自于种间,而比茎密度变异主要来自于种内,植物叶片和木质部密度的差异能够反映物种在生态策略的差异,通常高叶片面积表明树种采取的是高资源获取的策略,而高木质部密度采取的保守的策略.本研究在亚热带地带性植被常绿阔叶林的次生群落,各植物个体之间存在强烈的资源竞争关系,处于有利竞争地位的个体生长速率加快,而处在不利竞争地位的个体生长缓慢,从而生长速率变异表现出以种内变异大于种间变异.
3.2 形态指标在种内种间水平上与生长速率的相关性
不同性状对生长速率的预测能力不同,在物种水平上,胸径、树高以及冠幅与生长速率显著正相关,生长速率在物种水平上与胸径、树高以及冠幅呈显著正相关,木质部密度与生长速率显著负相关.全球范围403种热带以及温带树种中,大部分物种的生长速率是随着个体大小上升,可以持续到上百年[20],但是Wright et al[21]研究挪威云杉生长速率、个体形态以及立地环境与个体死亡率之间关系时是发现生长速率与树高的相关性与群落最大树高有一定联系,当群落最大树高低于25 m时,树高与生长速率显著相关,而当群落最大树高高于25 m时,生长速率与树高没有显著相关性.本文研究结果在种间水平上胸径、树高、冠幅以及木质部密度与生长速率都存在显著相关性,而在种内水平上所研究的13个树种中7个物种的木质部密度与生长速率显著相关,而只有两个树种的树高与生长速率显著相关,不同水平上各因子与生长速率有着不同的相关性,在很大程度上是因为各因子的种内变异程度要大于种间变异,当以物种水平研究各性状间相关性时,以每物种的平均值来计算,而忽略了种内变异,使得在种间水平上各性状与生长速率有着较强的相关性.因此,种内和种间的变异来源以及各性状间不同水平变异程度相差较大可能是在种内种间水平上生长速率与其他性状相关性发生较大差异的主要原因.另一方面,每个物种取样个体大小分布不均衡,也是造成这种情况发生的主要原因,冠幅、胸径以及树高在种内变异系数都超过30%,其中冠幅的种内变异甚至超过了50%,因此,在未来研究中应增加每物种的取样个数,尽量使胸径、树高以及冠幅等分布均匀,以降低取样引起的系统误差.
3.3 形态指标在种内种间水平上与木质部密度的相关性
不同生态功能组群植物的木质部密度会有所差异,如落叶性的植物功能组群有着相对较低的木质部密度;木质部密度与降水量有着显著相关关系,木质部密度的大小会直接影响植物个体吸收及运输水分的能力,木质部较低的植物导管组织容易发生内裂而形成树干内部空穴,低木质部密度的植物个体在导管运输组织上的低经济投入使它们有着相对较快的生长速度[22],高木质部密度的植物个体有着相对较低的生长速率和死亡率,它们在单位体积的生物量投入大,具有更长的寿命和更强的抵抗环境变化的能力,因此木质部密度是反映植物生长快慢和耐受性权衡关系的一个重要指标[23].本文研究结果显示木质部密度在种内和种间与生长速率都表现出显著的负相关关系,即低木质部密度的个体生长速率较快,高木质部密度的植物个体生长速率慢,当以胸径为协变量研究种内种间木质部密度与生长速率的偏相关关系时,发现胸径对于木质部密度与生长速率的相关性没有显著影响,也就是说明木质部密度与生长速率的相关性是独立于个体大小的,这在一定程度上说明不同植物功能组群的这种快生长与低耐受性的权衡关系不受个体大小的影响,而是植物在有限的生存环境资源空间下表现出的一种生长策略.
[1] REES M, CONDIT R, CRAWLEY M, et al. Long-term studies of vegetation dynamics[J]. Science, 2001,293,5530:650-655.
[2] REICH P B. The evolution of plant functional variation: traits, spectra, and strategies[J]. International Journal of Plant Sciences, 2003,3(S):143-164.
[3] 唐青青,黄永涛,丁易,等.亚热带常绿落叶阔叶混交林植物功能性状的种间和种内变异[J].生物多样性,2016,24(3):262-270.
[4] PEREZ-HARGUINDEGUY N, GARNIER E, LAVOREL S, et al. New handbook for standardised measurement of plant functional traits worldwide[J]. Australian Journal of Botany, 2013,61(3):167-234.
[5] 刘晓娟,马克平.植物功能性状研究进展[J].中国科学:生命科学,2015(4):325-339.
[6] ALBERT C H, THUILLER W, YOCCOZ N G, et al. A multi-trait approach reveals the structure and the relative importance of intra-vs. inter-specific variability in plant traits[J]. Functional Ecology, 2010,24(6):1 192-1 201.
[7] BOOTH R E, GRIME J P. Effects of genetic impoverishment on plant community diversity[J]. Journal of Ecology, 2003,91(5):721-730.
[8] FRIDLEY J D, GRIME J P, BILTON M. Genetic identity of interspecific neighbors mediates plant responses to competition and environmental variation in a species-rich grassland[J]. Journal of Ecology, 2007,95(5): 908-915.
[9] IIDA Y, POORTER L, STERCK F, et al. Linking size-dependent growth and mortality with architectural traits across 145 co-occurring tropical tree species[J]. Ecology, 2014,95(2):353-363.
[10] 周刘丽.浙江天童木本植物功能性状种内变异的大小、来源和空间结构[D].上海:华东师范大学,2016.
[11] 杨建军,苏文华,王玲玲,等.高山栲叶性状种内变异及其与环境因子的关系[J].广东农业科学,2015,42(12):152-158.
[12] 李耕耘,钱美玲,欧晓昆,等.种内变异及其群落效应[J].云南大学学报(自然科学版),2012,34(S2):412-418.
[13] 刘晓娟,马克平.植物功能性状研究进展[J].中国科学:生命科学,2015(4):325-339.
[14] ACKERLY D, CORNWELL W. A trait-based approach to community assembly: partitioning of species trait values into within and among community components[J]. Ecology Letters, 2007,10(2):135-145.
[15] 王燕,刘苑秋,杨清培,等.江西大岗山常绿阔叶林群落特征研究[J].江西农业大学学报,2009,31(6):1 055-1 062.
[16] LAVOREL S, GARNIER E. Predicting changes in community composition and ecosystem functioning from plant traits: revisiting the Holy Grail[J]. Functional Ecology, 2002,16(5):545-556.
[17] VIOLLE C, ENQUIST B J, MCGILL B J, et al. The return of the variance: intraspecific variability in community ecology[J]. Trends in Ecology & Evolution, 2012,27(4):244-252.
[18] JUNG V, ALBERT C H, VIOLLE C, et al. Intraspecific trait variability mediates the response of subalpine grassland communities to extreme drought events[J]. Journal of Ecology, 2014,102(1):45-53.
[19] WEIHER E, FREUND D, BUNTON T, et al. Advances, challenges and a developing synthesis of ecological community assembly theory[J]. Proceedings of Philosophical Transactions of the Royal Society B Biological Sciences, 2011,366,1576:2 403-2 413.
[20] STEPHENSON N L, DAS A J, CONDIT R, et al. Rate of tree carbon accumulation increases continuously with tree size[J]. Nature, 2014,507,7490:90-93.
[21] WRIGHT S J, KITAJIMA K, KRAFT N J B, et al. Functional traits and the growth-mortality trade-off in tropical trees[J]. Ecology, 2010,91(12):3 664-3 674.
[22] CHAVE J, COOMES D, JANSEN S, et al. Towards a worldwide wood economics spectrum[J]. Ecology Letters, 2009,12(4):351-366.
[23] LAFFOOREST-LAPOINTE I, MARTINEZ-VILALTA J, RETANA J. Intraspecific variability in functional traits matters: case study of Scots pine[J]. Oecologia, 2014,175(4):1 337-1 348.
(责任编辑:吴显达)
Intraspecific and interspecific variation of wood density and growth rates for the common tree species in subtropical evergreen broad-leaved forest
ZHANG Lingzhen, GUO Zhiwen, ZHENG Jingming
(Forestry College, Beijing Forestry University, Beijing 100083, China)
Wood density, growth rate of stem, tree height, diameter at breast height (DBH), crown area were measured for 13 common tree species in a subtropical evergreen broad-leaved forest. Trait variations of wood density and growth rate at interspecific and intraspecific level were determined, together with correlations between these 2 traits and other morphological indices. The results showed that intraspecific variations of wood density and growth rate were greater than those of interspecific variation. At interspecific level, tree height, DBH, crown area and wood density were significantly associated with growth rate. While at intraspecific level wood density and growth rate were negatively correlated, only two species of tree height and diameter at breast height were significantly associated with growth rate, the crown area had no significant association with a growth rate. Wood density was significantly correlated with growth rate at both intraspecific and interspecific level, regardless of tree size.
subtropical evergreen broad-leaved forest; interspecific variation; intraspecific variation; growth rate
2016-11-30
2016-12-20
林业公益性行业科研专项(201404303).
张令珍(1992-),女,硕士研究生.研究方向:生态恢复.Email:zhanglz92@163.com.通讯作者郑景明(1971-),男,副教授.研究方向:森林生态与恢复生态.Email:zhengjm@bjfu.edu.cn.
文献标识码: A 文章编号:1671-5470(2017)03-0293-06
10.13323/j.cnki.j.fafu(nat.sci.).2017.03.010
