H7N9亚型流感候选疫苗株的构建及免疫效力试验
2017-06-10蒋文明侯广宇李金平王素春袁万哲程善菊陈继明
蒋文明,侯广宇,李金平,王素春,袁万哲,王 灏,程善菊,陈继明
(1.中国动物卫生与流行病学中心,山东青岛 266032;2. 河北农业大学动物医学院,河北保定 071001)
H7N9亚型流感候选疫苗株的构建及免疫效力试验
蒋文明1,侯广宇1,李金平1,王素春1,袁万哲2,王 灏1,程善菊1,陈继明1
(1.中国动物卫生与流行病学中心,山东青岛 266032;2. 河北农业大学动物医学院,河北保定 071001)
近期的禽流感主动监测结果表明,我国部分H7N9亚型流感病毒已突变为高致病性毒株。为预防H7N9流感在禽群中引发大流行,本研究利用反向遗传技术,以鸡胚适应毒A/Puerto Rico/8/34(PR8)的内部基因为骨架,分别以H7N9亚型流感弱毒A/chicken/SH/3277/2016(S3277)和强毒A/chicken/SD/GDRZ/2017(GDRZ)的基因组为模板,扩增HA及NA基因,并对GDRZ HA基因进行修饰,去除裂解位点处的多个碱性氨基酸,使其获得低致病性流感病毒的分子特征,最终成功构建了H7N9流感疫苗候选株rS3277和rGDRZ。将两株病毒与自然分离的H7N3弱毒株A/duck/ZJ/1208/2009(ZJ1208)同时作为候选疫苗株,进行免疫效力试验。结果表明,rS3277、rGDRZ和ZJ1208灭活疫苗免疫SPF鸡21 d后,针对GDRZ的平均HI抗体效价分别达到7.2 log2、10.1 log2和6.2 log2。免疫攻毒保护试验结果表明,上述3种灭活疫苗均可100%保护SPF鸡免受H7N9强毒攻击。本研究所构建的rGDRZ重组候选疫苗株可为H7N9高致病性流感的防控提供支持。
流感病毒;H7N9;强毒;反向遗传;疫苗
2013年3月底H7N9亚型流感在我国出现并逐渐流行。虽然H7N9病毒弱毒株对家禽无致病性,但给我国养禽业(主要是鸡)造成了严重的经济损失,甚至导致了一定数量的人员感染和死亡,严重威胁着人类健康和公共卫生安全。更让人关注的是,2017年,我国有关机构主动监测发现,部分H7N9病毒已突变为对家禽具有高致病性的毒株,其对养禽业的威胁更大。2017年3月19日,湖南省永州市东安县的蛋鸡发生H7N9流感疫情,发病29 760只,死亡18 497只。该疫情虽然得到了及时有效的控制,但H7N9高致病性毒株的出现使得此次疫情备受关注。目前没有商品化的疫苗用于H7N9流感预防,因此有必要构建针对H7N9的疫苗候选株,以应对该病毒的流行和威胁。本研究以H7N9弱毒代表株S3277和强毒代表株GDRZ为表面基因供体,以PR8毒株的内部基因为骨架,构建了疫苗候选株rS3277和rGDRZ,同时以自然分离的H7N3弱毒作为候选疫苗株,进行免疫效力和攻毒保护试验,旨在为H7N9流感的防控提供储备疫苗。
1 材料与方法
1.1 病毒株、细胞、载体和实验动物
H7N9弱毒A/chicken/SH/3277/2016(S3277)、H7N9强毒A/chicken/SD/GDRZ/2017(GDRZ),本实验室分离保存;H7N3弱毒A/duck/ ZJ/1208/2009(ZJ1208),中国动物卫生与流行病学中心刘华雷博士赠送;双向表达载体pH205及表达PR8基因的pH205-PB2、pH205-PB1、pH205-PA、pH205-NP、pH205-M、pH205-NS,本实验室构建保存;293T细胞,本实验室保存;SPF鸡胚和SPF鸡,购自北京梅里亚维通实验动物技术有限公司。
1.2 主要试剂
QIAamp Viral RNA mini Kit,购自Qiagen公司;PrimeSTAR Max DNA Polymerase、PrimeScript Reverse Transcriptase、T4 DNA连接酶、pMD19-T、DL2000 DNA Marker、胶回收试剂盒、质粒提取试剂盒,购自TaKaRa公司;限制性内切酶BsmBI,购自NEB公司;转染试剂Lipofectamine 3000,购自Invitrogen公司。
1.3 引物设计
关于HA和NA基因的扩增引物,按照以前发表的文献合成[1]。设计引物使GDRZ强毒株HA裂解位点处的氨基酸序列“PKGKRTAR/GLF”突变成弱毒株的HA裂解位点“PKGK/GLF”(表1、图1)。

表1扩增HA、HA突变体、NA基因的引物
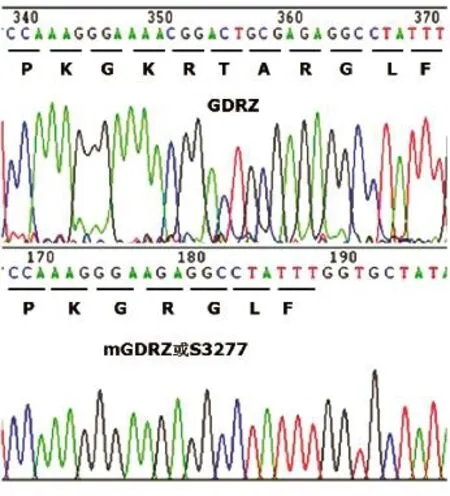
图1 GDRZ HA基因突变前后裂解位点处测序结果
1.4 病毒RNA的提取及基因扩增
对HA和NA基因的扩增,按照以前发表的文献进行[2]。按照QIAamp Viral RNA mini Kit说明书提取病毒RNA,利用PrimeScript Reverse Transcriptase进行反转录。以cDNA为模板,利用PrimeSTAR Max DNA Polymerase扩增病毒HA和N9基因片段。将PCR产物经琼脂糖凝胶电泳后,利用DNA胶回收试剂盒纯化回收,然后与T载体连接,将阳性重组质粒送青岛志远生物测序鉴定。
对于HA突变体的构建,以鉴定正确的GDRZ HA阳性重组质粒为模板,将2对引物Bm-HA-F/ HA1-R和HA2-F/Bm-HA-R分别扩增出HA1和HA2片段,然后以HA1和HA2的混合物为模板,Bm-HA-F/Bm-HA-R为引物,用PrimeSTAR Max DNA Polymerase进行重叠延伸PCR(SOE-PCR),扩增出HA突变体片段。将PCR产物经胶回收后,与T载体连接,对阳性重组质粒进行测序鉴定。
1.5 重组表达质粒的构建
将鉴定正确的GDRZ HAmut以及S3277 HA和N9重组质粒经BsmBI酶切后,与BsmBI酶切的表达载体pH205相连,提取重组质粒,进行测序鉴定,并利用DNAStar7.0软件进行序列分析。
1.6 病毒拯救与鉴定
具体操作按照Lipofectamine 3000脂质体转染试剂盒说明进行。将PR8的6个内部基因重组表达质粒、HA或HA突变体和对应的N9表面基因重组表达质粒各0.6 μg混合,共同转染293T细胞。转染48 h后,将细胞悬液接种10日龄SPF鸡胚,0.2 mL/枚,37 ℃继续孵化,72 h后收集尿囊液,测定其血凝活性。将血凝阳性的尿囊液经10 000倍稀释后接种10日龄SPF鸡胚,扩增病毒,0.1 mL/枚,72 h后收集尿囊液,于-70 ℃保存备用。重组病毒命名为rS3277和rGDRZ。对提取救获的病毒RNA,用RT-PCR扩增全基因组并测序。
1.7 疫苗制备
将病毒经10 000倍稀释后接种10日龄SPF鸡胚,扩增病毒,0.1 mL/枚,72 h后收集尿囊液,加入甲醛至终浓度0.1%,2~8 ℃灭活48 h。将尿囊液和白油佐剂按照1:2的比例,乳化制成灭活油佐剂疫苗。
1.8 免疫保护试验
将40只21日龄SPF鸡分成4组,每组10只;将各组鸡分别颈部皮下接种rS3277、rGDRZ和ZJ1208灭活油苗,接种剂量均为0.3 mL/只,其中1组不接种疫苗,直接攻毒组作为对照。免疫21 d后采血[3],分离血清,测定抗体效价,并用H7N9强毒GDRZ(EID50=106)对各组鸡进行滴鼻攻毒,连续观察10 d,统计排毒、发病和死亡情况。
2 结果
2.1 表达HA或HA突变体以及N9重组质粒的构建
采用SOE-PCR,扩增GDRZ株的HA基因,并将其克隆至pH205载体中,构建pH205-GDRZmHA重组质粒。测序结果表明(图1),HA裂解位点的氨基酸序列已由强毒病毒HA分子的“-PKGKRTAR-”突变为弱毒病毒HA分子的“-PKGK-”。按照常规方法扩增HA和N9,构建重组表达载体pH205-GDRZNA、pH205-S3277HA、pH205-S3277NA。
2.2 重组病毒的拯救与鉴定
将成功构建的8个重组质粒转染293T细胞,利用反向遗传技术,拯救出重组病毒rS3277和rGDRZ;将其在鸡胚中繁殖1代后,提取病毒RNA,进行反转录,对拯救病毒的全基因组进行扩增、测序。测序结果表明,HA基因裂解位点已突变为“-PKGK-”,为低致病性H7N9病毒分子特征,所有8个片段序列均与预期的一致。
2.3 候选疫苗的免疫保护和交叉免疫保护试验
候选疫苗的免疫和攻毒保护试验结果见表2。免疫21 d后,rS3277疫苗对母本病毒的平均HI滴度为10.3 log2,对强毒GDRZ的平均HI滴度为7.2 log2;rGDRZ疫苗对母本病毒的平均HI滴度为10.1 log2;ZJ1208疫苗对母本病毒的平均HI滴度为9.7 log2,对强毒GDRZ的平均HI滴度为6.2 log2。

表2候选疫苗对SPF鸡的免疫和攻毒保护试验结果
攻毒保护试验结果显示,rS3277和rGDRZ重组灭活疫苗,以及H7N3自然分离弱毒制备的灭活疫苗,为SPF鸡提供的针对H7N9强毒GDRZ的保护率均为100%。未免疫对照组,经强毒GDRZ攻毒后5 d内全部死亡。
3 讨论
2013年,H7N9亚型流感在我国长三角地区出现,并迅速向全国蔓延[4]。2017年以前,在家禽中分离的H7N9病毒绝大多数为弱毒株,对家禽没有致病力。2017年前监测数据显示,有些地区的部分H7N9病毒突变为高致病性毒株,对鸡有高致病力,对养禽业带来极大威胁。
前期试验研究表明,以分离的H7N3自然弱毒ZJ1208免疫小鼠,可以产生针对H7N9病毒的HI抗体和中和抗体[5]。因此,本试验将ZJ1208作为候选疫苗株进行试验。同时,在生产中,自然分离的H7N9病毒可能存在一定的潜在危险,所以本研究利用反向遗传技术,以鸡胚高产适应毒A/Puerto Rico/8/34(PR8)的内部基因为骨架,分别以弱毒株S3277和强毒株GDRZ的HA和NA表面基因为供体,通过对GDRZ HA基因进行分子突变修饰,去除HA蛋白裂解位点处的多个碱性氨基酸,使其获得H7N9低致病性流感的分子特征,对鸡胚和SPF鸡均不能发病和致死,同时又保证了重组病毒与流行株的抗原匹配性,成功构建了针对H7N9亚型流感的疫苗候选株rS3277和rGDRZ。
SPF鸡免疫结果显示,免疫rS3277、rGDRZ和ZJ1208灭活油苗21 d后,血清中针对母本病毒的平均HI抗体效价分别达到10.3 log2、10.1 log2和9.7 log2,说明重组疫苗具备良好的免疫原性。但是,免疫rS3277和ZJ1208产生的,针对GDRZ强毒株的平均HI抗体效价分别为7.8 log2和6.2 log2,与针对母本病毒的平均HI抗体效价分别相差了3.1 log2和3.5 log2,说明rS3277和ZJ1208与GDRZ强毒株之间有一定的抗原性差异。HA氨基酸序列对比结果显示,S3277和ZJ1208与GDRZ之间的同源性分别为97.1%和95.9%,这与HI试验结果一致。
攻毒保护试验结果表明,rS3277和rGDRZ重组灭活疫苗,以及自然分离的ZJ1208制备的灭活疫苗,均可以对SPF鸡提供针对强毒100%的保护。但是相对rS3277和ZJ1208候选疫苗株,rGDRZ可以产生针对强毒更高的HI抗体滴度。本研究构建的rGDRZ重组病毒株作为候选疫苗株,为H7N9流感的防控提供了可行的探索和尝试,为H7N9流感的防控提供了技术支持。
[1] HOFFMANN E,STECH J,GUAN Y,et al. Universal primer set for the full-length amplification of all influenza A viruses[J]. Arch Virol,2001,146(12):2275-2289.
[2] 蒋文明,侯广宇,王素春,等. H5N6亚型禽流感病毒反向遗传疫苗株的构建及免疫保护试验[J]. 中国动物检疫,2015,32(1):64-67.
[3] 宋洋铭,谷春阳,曾显营,等. Clade7.2 H5N1亚型禽流感重组病毒疫苗株的构建及免疫效力评价[J]. 中国预防兽医学报,2016,38(7):580-584.
[4] 蒋文明. 从“H7N9禽流感”命名说开去[J]. 中国动物检疫,2014,31(8):80-82.
[5] JIANG W M,WANG S C,LIU H L,et al. Evaluation of avian inf l uenza virus isolated from ducks as a potential live vaccine candidate against novel H7N9 viruses[J]. Vaccine,2014,32(48):6433-6439.
(责任编辑:朱迪国)
Construction of H7N9 Subtype Inf l uenza Candidate Vaccine Strain and its Immune Eff i cacy Test
Jiang Wenming1,Hou Guangyu1,Li Jinping1,Wang Suchun1,Yuan Wanzhe2,Wang Hao1,Cheng Shanju1,Chen Jiming1
(1. China Animal Health and Epidemiology Center,Qingdao,Shandong 266032;2. College of Animal Medicine,Hebei Agricultural University,Baoding,Hebei 071001)
Recently,the nationwide active surveillance results revealed that a number of H7N9 subtype influenza A viruses have evolved into highly pathogenic strains in China. In order to prevent the potential pandemic caused by the mutant viruses to poultry,two reassortant viruses,tentatively named rS3277 and rGDRZ,were generated using reverse genetics in this study. The rescued viruses contained intact or modif i ed HA and NA genes of currently circulating isolates A/ chicken/SH/3277/2016 and A/chicken/SD/GDRZ/2017,respectively,while containing the internal genes derived from A/ Puerto Rico/8/34(PR8). The novel reassortant candidate vaccine virus,rGDRZ,with deletion of the multi-basic amino acid motif in the HA gene,was served as a vaccine candidate and examined for immune eff i cacy,together with rS3277 and an naturally isolated H7N3 virus,A/duck/ZJ/1208/2009(ZJ1208). The results showed that the HI antibody titers against GDRZ reached 7.2 log2,10.1 log2 and 6.2 log2,respectively,when serum of SPF chickens immunized with rS3277,rGDRZ and ZJ1208 was tested on 21 days post-immunization. Importantly,the results of immune challenge test showed that the rS3277,rGDRZ and ZJ1208 vaccines protected SPF chickens from H7N9 highly pathogenic GDRZ virus infection completely. Taken together,the recombinant candidate vaccine strain constructed by this research can provide strong support for the prevention and control of H7N9 highly pathogenic avian inf l uence.
inf l uenza virus;H7N9;virulent virus;reverse genetics;vaccine
S852.65
:A
:1005-944X(2017)06-0086-04
10.3969/j.issn.1005-944X.2017.06.025
科技部科技基础性工作专项(2012FY111000)
