乌鲁木齐县石人沟地衣物种多样性及群落特征1)
2017-06-05艾尼瓦尔吐米尔夏衣拉肖开提NazarbekGuldan阿不都拉阿巴斯
艾尼瓦尔·吐米尔 夏衣拉·肖开提 Nazarbek Guldan 阿不都拉·阿巴斯
(新疆大学,乌鲁木齐,830046)
乌鲁木齐县石人沟地衣物种多样性及群落特征1)
艾尼瓦尔·吐米尔 夏衣拉·肖开提 Nazarbek Guldan 阿不都拉·阿巴斯
(新疆大学,乌鲁木齐,830046)
为了查明新疆乌鲁木齐县石人沟地区的地衣物种多样性及其区系和群落特征,在石人沟地区的不同植被带设立16个样点,对地衣进行了样方调查、标本采集和标本鉴定,并对地衣区系成分和生长型进行了分析。结果表明,石人沟地区地衣共有30种,隶属于17属,10科,5目,其中叶状地衣和壳状地衣种类较丰富。地衣区系成分划分为世界广布成分、环北极成分、温带成分、中亚物种和中国特有物种等5个区系成分;按照该地区地衣生长的现状,把其划分为岩面生、树生、藓丛生和地面生等4个类型;该地区地衣的生长型主要以壳状地衣为主,其次为叶状地衣,缺乏枝状地衣种类。同时基于30种地衣及盖度数据,应用双向指示种分析方法(TWINSPAN)和除趋势对应分析(DCA)将该地区的地衣划分为3个样点组,并计算了物种多样性和均匀度指数。
地衣多样性;群落分类;生长型;地衣区系
With the lichen of Shirengou at 16 sites in Urumqi County lichens floristic composition, we studied the growth form characteristics according to the spot investigation data. Thirty lichen species in 17 genera and 10 families were found growing on rocks and tree barks. The crustose lichens dominated in this area as well as some foliose lichens species. The lichen floristic consists of such as Cosmopolitan element, Circumpolar arctic element, Temperate zone elements, Middle-Asian species and Species endemic to China. The lichens were divided into four ecological types according to the substrate types and their growth form. With 30 recorded lichen species and their coverage, we applied Two-way Indicator Species Analisis (TWINSPAN) and Detrended Correspondence Analysis (DCA) to classify the lichen species into three groups, and calculate the species diversity and evenness index.
地衣是由真菌和藻类的高度结合而形成的共生复合体,是陆地生态系统生物多样性的主要成员之一。地衣特殊的形态和生理特征有利于它广泛的分布和能在一些极端的环境中生长,因此,从热带到两级各种不同的景观中可以找到地衣的踪迹[1-3]。地衣对生态环境的污染非常敏感,近年来,随着全球气温变暖、环境污染等全球性环境问题影响下地衣的多样性出现减少,分布范围不断的缩小,因此,研究地衣物种多样性及资源分布在今后有效保护生物多样性方面具有重要的意义。
20世纪80年代之前国内外有些研究人员对新疆地衣进行了零散性的报道[4-8]。从20世纪80年代初开始我国地衣学家系统地研究新疆的地衣资源,已在地衣区系、系统分类、地衣生态学、地衣化学成分分析和地衣分子生物学等领域取得了不少成果,促进了新疆地衣学的发展[9-16]。目前,新疆地衣的研究随着我国地衣学研究的迅速和稳定的发展而进入迅速发展的阶段。
乌鲁木齐县石人沟地处深深的山沟和两边陡峭的山坡形成的山沟体系。山沟南坡太阳照射强烈、温度高,而北坡的太阳辐射较弱、没有充足热量、温度较低,因而形成了温度高低差异较特殊的地带性环境。这种特殊的地理环境为地衣的生长提供了有利的生境,因此,丰富了该地区的地衣物种多样性。但随着近年来的旅游事业的发展,导致了该地区的环境污染和生态环境的破坏,影响了该地区生物多样性,特别是该地区的地衣种类多样性及分布区不同程度的受到了人类活动和环境污染的影响。因此,对该地区地衣物种多样性和地衣群落进行研究在地衣资源保护和开发利用方面具有一定意义。
1 研究区概况
石人沟位于天山中段北部,准葛尔盆地南部,地处87°50′N,43°45′E,属于乌鲁木齐县。乌鲁木齐县地处天山北麓,准噶尔盆地南缘,东经86°37′56″~88°58′22″,北纬43°1′8″~44°6′11″。东与达坂城区接壤,南以天山吐格塔格达坂为界与托克逊县、和静县相依,西以头屯河为界与昌吉市为邻,北和米东区毗连。石人沟具有温带大陆性气候,温差大,山区年平均气温为3.5 ℃,年均降水量208.4 mm,年均蒸发量2 616.9 mm,年均无霜期179 d,年均日照时间2 813.5 h[17]。
2 研究方法
2.1 野外调查和标本鉴定
野外调查包括地衣的种类调查及盖度和频度调查、标本采集等。乌鲁木齐县石人沟地区常见的植物种类有雪岭云杉(PiceaschrenkianaFisch. et Mey.)、密叶杨(PopulustalassicaKom.)、白榆(UlmuspumilaL.)、小叶忍冬(LoniceramicrophyllaWilld. ex Roem. et Schult.)和腺毛蔷薇(RosafedtschenkoanaRegel)等。在不同景观随机设置样点16个,在各个样点各选取1~3个大小为50 m×50 m样地进行标本采集和样方调查,共设立37个样地。每个样地中随机设立样方若干个,样方大小为50 cm×50 cm,样方间隔2 m。每个样方中不同种类的盖度用网格法测定,以地衣在样方中的平均盖度作为样点的盖度[15,18]。采用形态观察法、化学显色反应法、地衣微量化学结晶法、薄层色谱法等常用方法对所采集的地衣标本进行种类鉴定[1,12]。研究中所用的标本保存于新疆大学西北干旱区地衣研究中心地衣标本馆。
2.2 数据分析
本研究在野外收集数据的基础上,30个地衣种和16个样点构成30×16的矩阵,应用双向指示种分析方法(TWINSPAN)以样点为对象,以地衣的盖度为指标,对样点进行分组,分析各组地衣总盖度。运用除趋势对应分析法(Detrended Correspondence Analysis,DCA)[15,18],并结合TWINSPAN分析结果进行地衣群落分类。使用WinTWINS 2.3程序和MVSP软件包中的DCA程序进行数据分析[15,18]。
根据地衣的种类和覆盖度,计算样点组的Shannon-Weiner物种多样性指数和Pielou均匀度指数,比较各样点组的物种多样性特点。
Shannon-Weiner多样性计算公式[19-20]:
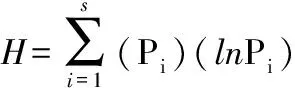
其中:Pi=ni/N,N为样点地衣覆盖度;ni为第i种的覆盖度。
均匀度计算公式为:
J=H/Hmax。
其中:J为均匀度指数;H为实测多样性值;Hmax为理论上群落多样性的最大值,Hmax=lns,其中s为总种数。
3 结果与分析
3.1 石人沟地衣种类组成
文中鉴定石人沟的地衣共有30种,隶属于17属、10科、5目(表1)。

表1 乌鲁木齐县石人沟的地衣名录
从表1可知,石人沟地区茶渍目(Lecanorales)地衣种类占优势,其科、属和种数分别占该地区地衣科、属、种总数的60.00%、58.82%和53.33%。黄枝衣目(Teloschistales)地衣共有5种,2属、1科,分别占该地区地衣科、属、种总数的10.00%、11.76%和16.67%。鸡皮衣目(Pertusariales)地衣共有6种,3属、1科,分别占该地区地衣科、属、种总数的10.00%、17.64%和20.00%。地卷目(Peltigerales)和瓶口衣目(Verrucariales)的地衣种类较少。单种和单属地衣科有3个,分别是胶衣科(Collemataceae)、梅衣科(Parmeliaceae)和地卷科(Peltigeraceae),分别占石人沟地区地衣科、属、种总数的10.00%、5.88%和3.33%。
石人沟地区地衣的生长型主要以壳状和叶状地衣为主,缺乏枝状地衣。其中壳状地衣共有19个,占总种数的63.33%;叶状地衣有11个,占总种数的36.67%(表2)。地衣对生长基物有专一性,大多数的地衣种类在长期的协同进化过程中,通过改变自己的生长型、生活型来适应自己的生长基物,但也有的地衣种类可以生长在2种以上的基物上。本研究结果表明,在石人沟地区生长在1种基物上的地衣共有18个,占地衣总数的60%;生长在2种基物上的地衣有9个,占地衣总数的30%;生长在3种基物上的地衣有3个,占地衣总数的10%(表1)。
3.2 石人沟地衣的地理区系
新疆地衣植物在区系成分上以环北极成分为主,这主要与新疆的地理位置、气候、地质历史变化有关系。本研究根据新疆地衣的地理成分特征[12],参照和综合Thomson[21-22]、Wei[23]、魏江春等[24]、吴征镒等[25-26]的研究资料,将乌鲁木齐石人沟地衣的地理成分划分如下。
世界广布成分:包括皮果衣(D.miniatum(L.) Mann.)、短绒皮果衣(D.vellereumZsch. Rabh.)、石黄衣(X.parietina(L.) Beltr.)、粉芽石黄衣(X.sorediata(Vain.) Poelt)4个种,占总种数的13.33%。
环北极成分:包括丽石黄衣(X.elegans(L.) Beltr.)、蓝灰蜈蚣衣(Ph.caesia(Hoffm.))、破小网衣(L.carpathicaKörb.)、墨绿胶衣(C.fuscovirens(With.) Laund.)、平小网衣(L.sigmatea(Ach.))、C.calcarea(L.) A. Nordin.、北平茶渍(A.contorta(Hoffm.) Körb.)、灰平茶渍(A.cinerea(L.) Körb.)、金黄茶渍(C.aurella(Hoffm.))、被膜微孢衣(A.molybdinaTrevis.)、丛生微孢衣(A.veronensisA. Massal.)、小蜡盘橙衣(C.biatorina(A. Massal.))、粉盘裂片茶渍(L.alphoplaca(Wahlenb.) Hafellner)共13种,占总种数的43.44%。
温带成分:包括裂片石黄衣(X.lobulata(Flörke) B. de Lesd.)、碎茶渍(L.argopholis(Ach.) Ach.)、墙茶渍(L.muralis(Schreb.) Rabenh.)、裂芽地卷(P.praetextata(Flörke ex Sommerf.))、盾脐鳞衣(R.peltata(Ram) Leuck. et Poelt)、翘叶蜈蚣衣(Ph.adscendens(Fr.))、暗裂牙黑蜈蚣衣(Ph.sciastra(Ach.) Moberg)、鳞饼衣(D.oreina(Ach.) Norman.)8个种,占总种数的26.67%。
中亚物种:包括亚洲平茶渍(A.asiatica(H. Magn.))、托敏氏褐梅(M.tominii(Oksner) Essl.)2个种,占总种数的6.67%。
中国特有物种:包括疣微孢衣(A.verruculosaH. Magn.)、油黄茶渍(C.oleiferaH. Magn.)、赭白平茶渍(A.ochraceoalba(H. Magn.))3个种,占总种数的10%。
3.3 石人沟地衣群落结构
对30个地衣物种在16个样点407个样方中的分布以样点为对象、地衣盖度为指标(表2)进行了TWINSPAN和DCA分析(图1、图2)。
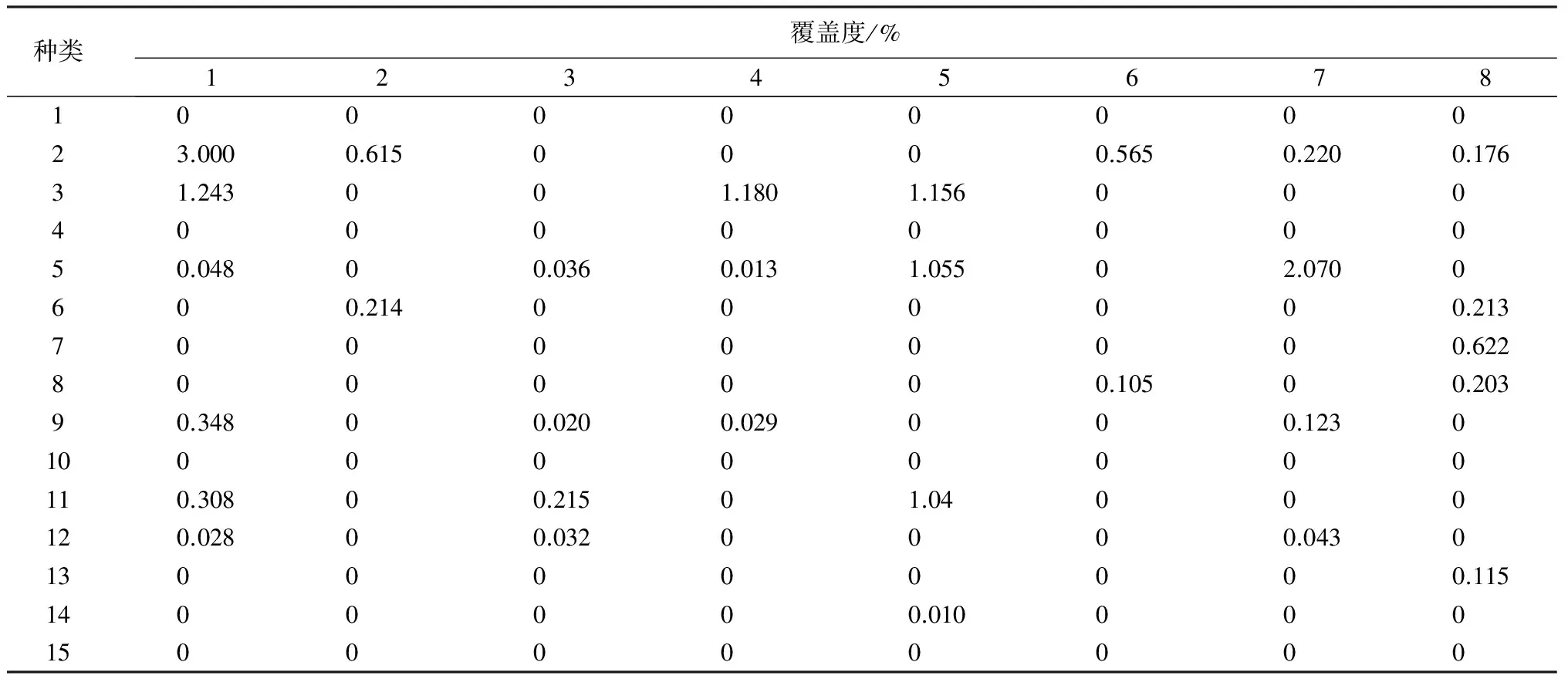
表2 30种地衣在16个样地中的覆盖度
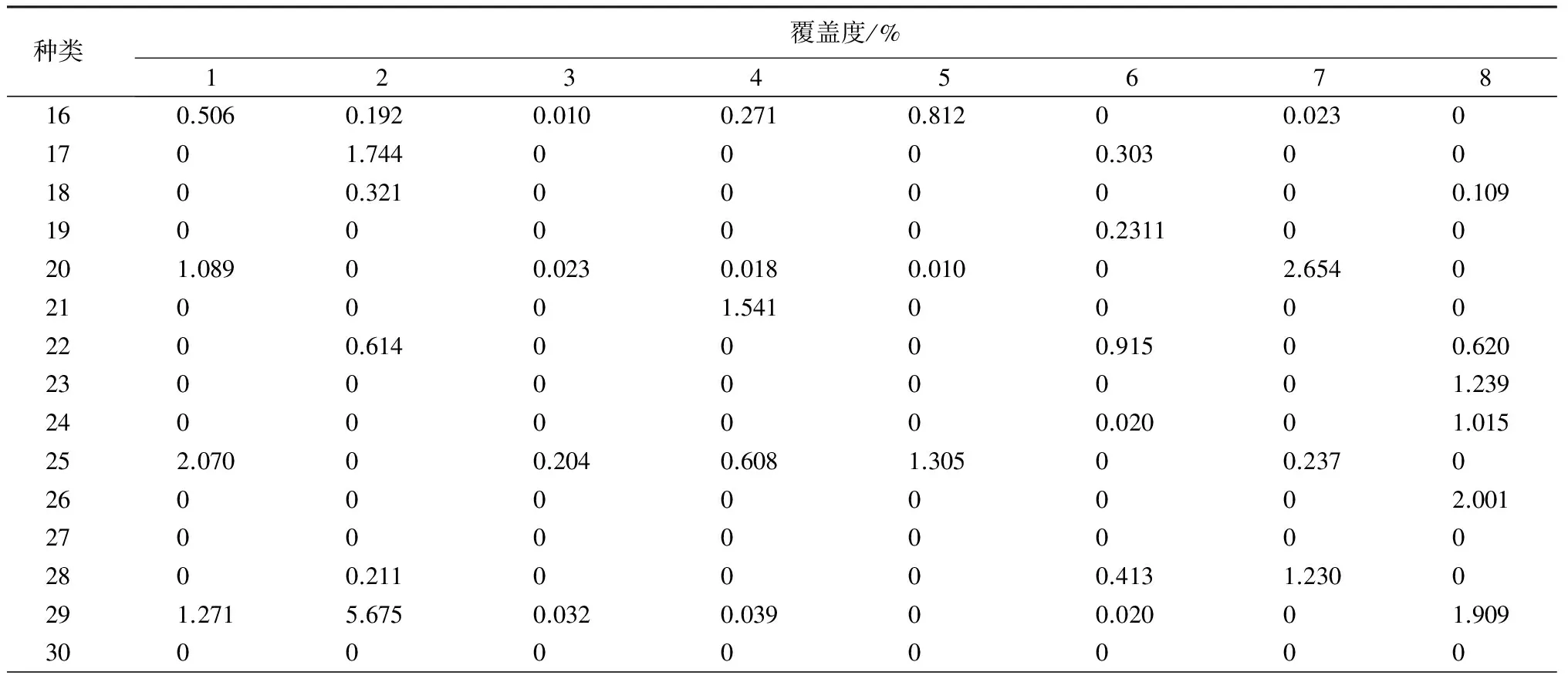
续(表2)

种类覆盖度/%91011121314151610000.3310.21500.0250.31321.2390.20300.243000030000000041.4030.130000.0400.3550.0340500000000.54461.4480.3150.0360.1830000700.022000.31506.0400.32583.0700.014000000.52290000.02900001000.1430.0200000011000000001200000.239000130000.411000.3280140.0230.0100.20300.89202.12301500000.0210.14500.1221600003.07200.1430170.0220.0150000.53200180.0200.016000.524000.21319000.0920.129000.6080.13520000000002100.0260.0230.041004.0100.250220.0300.2130.023000002300.07200.0170.2340.21500.1892402.00100.054000.054025000.0640.562000.6230260.0810.035000000.54127000000.5230.532028000.0540.5410.014000290.0320.043000000.769300000.1140.1330.0320.9263.042
根据TWINSPAN和DCA分析结果,16个样点可分成3个组(表3)。样点组1包括样点1、3、4、5和7共5个样点。主要的地衣种类有丛生微孢衣、疣微孢衣、油黄茶渍、盾脐鳞衣等,共13种。地衣的总盖度为24.063%,其中平均盖度1%以上的地衣有10种,盖度最大的是盾脐鳞衣,为3.794%。
样点组2包括样点2、11、12、13、15、16共6个样点。主要地衣有油黄茶渍、破小网衣、亚洲平茶渍、北平茶渍等24种。地衣的总盖度为41.532%,其中平均盖度1%以上的地衣有10种,盖度最大的是碎茶渍,为6.680%,其次为皮果衣,为6.444%。
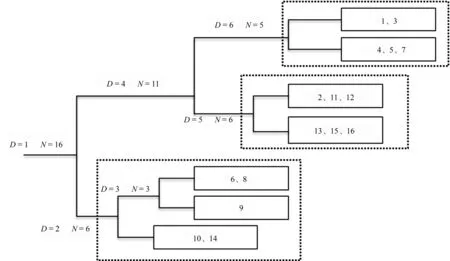
D.分类水平;N.样点总数;框中的数字代表16个样点编号。

图中的数字表示16个样点编号。
样点组3包括样点6、8、9、10和14共5个样点。主要地衣有丛生微孢衣、金黄茶渍、托敏氏褐梅、赭白平茶渍等21种。地衣的总盖度为30.945%,其中平均盖度1%以上的地衣有9种,盖度最大的是暗裂牙黑蜈蚣衣,为4.172%,其次为翘叶蜈蚣衣,为3.888%。
样点组2的多样性最大,为2.871,其次是样点组3,为2.195,样点组1的多样性较低,为1.782。对Pielou均匀度指数的分析中可以看出,样点组3的均匀度最高,为0.903,其次为样点组2的均匀度,为0.721,样点组1的均匀度最低,为0.695。
4 结论与讨论
本研究在实地调查和地衣标本的鉴定整理的基础上,初步确定分布在乌鲁木齐县石人沟地区的地衣共有30种,隶属于17属,10科。研究发现该地区地衣的区系成分主要以环北极和温带成分占优势,占70.11%;同时还包括世界广布成分、中亚物种和中国特有种类。因为该地区的气候干旱,植被以旱生植物为主,长期干旱气候的影响下地衣区系中的热带成分的地衣种类逐渐消失,与此同时,环北极和温带成分地衣逐渐适应该地区取代其他热带地衣种类,从而形成了以温带和环北极地衣种类为主的地衣区系,尤其是耐旱型壳状地衣的优势比较明显,符合该地区高等植物区系的发展和形成趋势。文中对已知地衣种类的生长型和基物种类进行分析并发现,研究地区地衣种类的分布不仅随着植被地带性而变化,与着生的基物也有密切的关系。一般岩面生地衣的种类和分布与岩石的理化性质、岩石的大小、岩石的方向等有关;而树附生地衣的分布主要受到树种、树干和树枝的理化特性、水分含量、表面粗糙度、树干大小、树木年龄等多种因素的限制[27-29]。已有的文献表明,地衣的生长和代谢与水分的关系很大[30-31]。
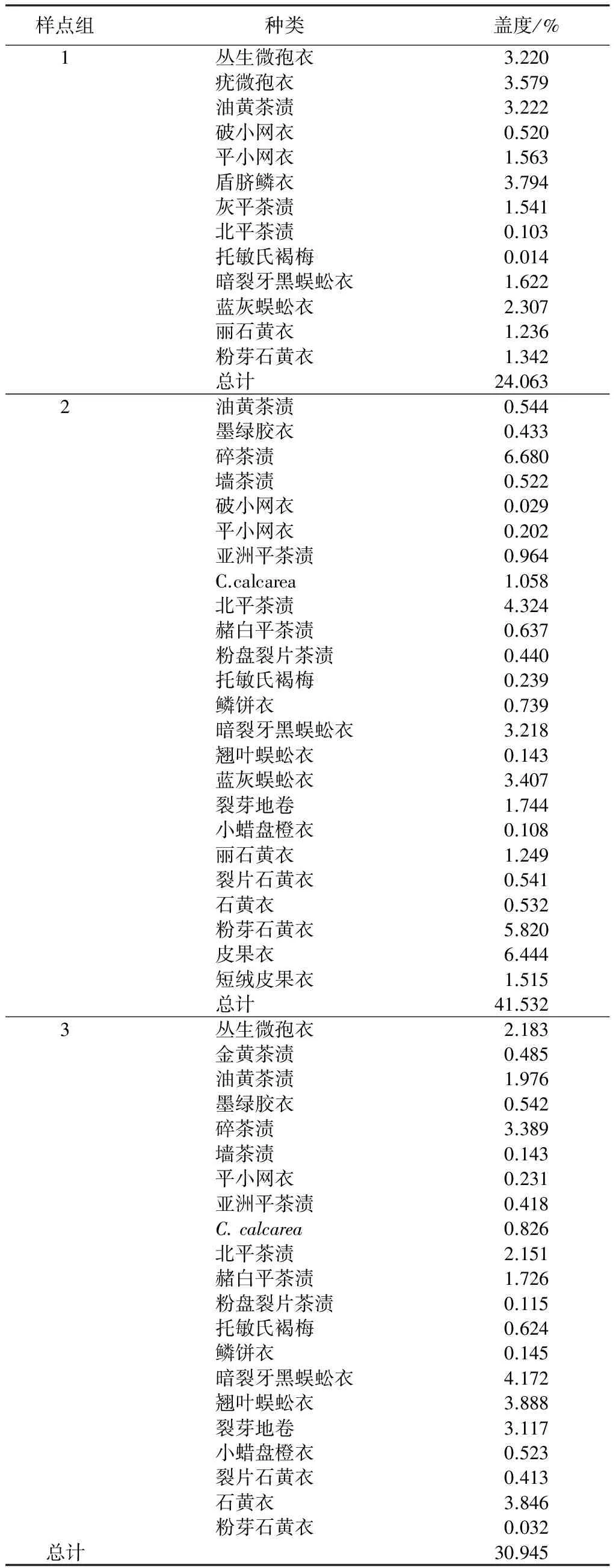
表3 各样点组中地衣的盖度平均值统计
在严酷的干旱条件下,地衣进入休眠状态,降低呼吸和其他生理代谢以适应环境;而待有水分时,很短时间内能够吸水饱和,进行光合作用和呼吸作用,生长速度加快[30-31]。岩石是干旱区壳状地衣种类的主要栖息地和生长基物,在植物群落演替的早期阶段壳状地衣通过分泌次生代谢物质来不断地腐蚀岩石,逐渐改变岩石的理化性质,形成土壤,为枝状和叶状地衣的侵入、定居提供微环境,从而促进地衣群落的演替。本研究发现,在石人沟地区专一的岩面生地衣共有17个,占地衣总数的56.67%,充分体现了干旱地区地衣对环境的适应能力,与阿不都拉·阿巴斯等[12]报道的新疆地衣的生长型和基物类型保持一致。
文中使用植物群落定量研究中常用的TWINSPAN和DCA排序方法对该地区地衣种类进行了群落划分,并分析了个群落的多样性。结果表明,分布在乌鲁木齐县石人沟的地衣可分成3个样点组,各样点组的地衣种类、物种多样性和覆盖度间存在显著性差异。样点组1共有地衣13种。地衣的总盖度为24.063%,其中平均盖度1%以上的地衣有10种,盖度最大的是盾脐鳞衣,为3.794%;样点组2共有地衣24种,地衣的总盖度为41.532%,其中平均盖度1%以上的地衣有10种,其中盖度最大的是碎茶渍,为6.680%,其次为皮果衣,为6.444%。样点组3地衣有9种,盖度最大的是暗裂牙黑蜈蚣衣,为4.172%,其次为翘叶蜈蚣衣,为3.888。已有的研究表明,岩面生地衣的生长和分布与岩石种类,岩石化学成分等因素有密切相关[32-33]。朽木生地衣的多样性受到树种、森林的年龄、森林生态系统的结构和动态、植物群落演替、光照强度、森林郁闭度等多种因素的影响[29]。树附生地衣丰富度和分布主要受到光照、温度、水分、树皮酸碱度、养分、空气污染、林龄、树木连续性、火等,以及附载植物胸径、树高、树种因素的控制[2,34]。艾尼瓦尔·吐米尔等[16]对乌鲁木齐市郊区地衣分布格局与环境关系进行研究时发现,地衣群落种类的分布与郁闭度、人为干扰和湿度等环境因素有一定相关性。本研究发现,石人沟地区的地衣群落结构和地衣物种分布随着海拔、基物种类、离人为干扰的距离和干扰的种类、湿度、光照强度发生变化,有关地衣群落物种分布与环境因素关系的定量研究结果将另文报道。综上所述,乌鲁木齐县石人沟地区的地衣种类主要以耐旱、耐寒的环北极和温带地衣种类组成,地衣种类的分布受到多种自然和人为因素的影响。近年来,该地区旅游事业的发展导致了环境污染的加剧和生态环境的破坏,从而影响了该地区本来较脆弱的生态系统的稳定性,因此,该地区地衣种类的分布和多样性的研究将对该地区环境质量的综合评价具有重大意义。
[1] HALE M E. The biology of lichens[M]. 3 edition. London: Edward Arnold Publisher,1983:84-96.
[2] HAUCK M. Site factors controlling epiphytic lichen abundance in northern coniferous forests[J]. Flora,2011,206(2):81-90.
[3] BRODO I M. Substrate ecology[M]//Ahmadjian V, Hale M E. The lichens. New York: Academic Press,1973:401-441.
[4] ELENKIN A A. Migratory lichens of the deserts and steppes[J]. Bulletin du Jardin Imperial Botanique de St Petersburg,1901,1(1):16-38.
[5] ELENKIN A A. Lichens florae rossiae et regionum confinium orientalium. FasciculusⅠ(Numeris:1~50)[J]. Acta Horti Petropolitani,1901,19(1):1-52.
[6] 刘慎谔.中国北部及西部植物地理概论[J].国立北平研究院植物学研究所丛刊,1934,2(9):423-451.
[7] MOREAU F, MOREAU F. Lichens de China[J]. Revue Bryologique et Lichenologique,1951,20(1/2):183-199.
[8] VAINIO E A. Lichenes ab ove paulsen praecipue in provencia ferghana (Asiamedia) et a boris fedtschendo in tjanschen anno 1898 et 1899 collecti[J]. Botanisk Tidsskrift,1904,26(5):241-250.
[9] 赵继鼎,徐连旺,孙增美.中国地衣初编[M].北京:科学出版社,1982.
[10] 王先叶.天山托木尔峰地区的生物[M].乌鲁木齐:新疆人民出版社,1985:328-353.
[11] 吴金陵.新疆草地地衣[J].植物分类学报,1985,5(1):73-75.
[12] 阿不都拉·阿巴斯,吴继农.新疆地衣[M].乌鲁木齐:新疆科技卫生出版社,1998.
[13] ABBAS A, MIJIT H, TUMUR A. A checklist of lichens of the Xinjiang, China[J]. Harvard Paper in Botany,2001,5(2):359-370.
[14] 努尔巴衣·阿不都沙力克,阿不都拉·阿巴斯.新疆哈纳斯自然保护区地衣地理分布规律的研究[J].吉林农业大学学报,1998,20(增1):231-235.
[15] 艾尼瓦尔·吐米尔,阿不都拉·阿巴斯.新疆托木尔峰国家级自然保护区树附生地衣群落数量分类[J].东北林业大学学报,2015,43(5):97-101.
[16] 艾尼瓦尔·吐米尔,买买提·沙塔尔,阿布都拉·阿巴斯.乌鲁木齐市郊地衣分布格局及其与环境的关系[J].东北林业大学学报,2013,41(4):76-82.
[17] 乌鲁木齐县地方志编写委员会.新疆维吾尔自治区地方志丛书:乌鲁木齐县志[M].乌鲁木齐:新疆人民出版社,2006.
[18] 曹同,陈怡,于晶,等.上海市地面生藓类植物的分布格局分析[J].应用生态学报,2004,15(10):1785-1791.
[19] 王伯荪.植物群落学[M].北京:高等教育出版社,1987:217-233.
[20] 张金屯.植被数量生态学方法[M].北京:北京科学技术出版社,1995.
[21] THOMSON J W. Lichens of alaskan arctic slope[M]. London: Toronto University Buffalo,1979.
[22] THOMSON J W. American arctic lichens 1-The macrolichens[M]. New York: Columbia University Press,1984.
[23] WEI J C. An enumeration of lichens in China[M]. Beijing: International Academic Publishers,1991.
[24] 魏江春,姜玉梅.西藏地衣[M].北京:科学出版社,1986.
[25] 吴征镒.中国植被[M].北京:科学出版社,1980.
[26] 吴征镒,王荷生.中国自然地理:植物地理:上册[M].北京:科学出版社,1983.
[27] McMULLIN R T, DUINKER P N, RICHARDSON D, et al. Relationships between the structural complexity and lichen community in coniferous forest of southwestern Nova Scotia[J]. Forest Ecology and Management,2010,260(5):744-749.
[28] HAUCK M. Site factors controlling epiphytic lichen abundance in northern coniferous forests[J]. Flora,2011,206(2):81-90.
[29] NASCIMBENE J, THOR G, NIMIS P L. Effects of forest management on epiphytic lichens in temperate deciduous forests of Europe-a review[J]. Forest Ecology and Management,2013,298(8):27-38.
[30] HAUCK M, RUNGE M. Stem flow chemistry and epiphytic lichen diversity in dieback-affected spruce forest of the Harz Mountains, Germany[J]. Flora,2002,197(4):250-261.
[31] SCHMULL M, HAUCK M, VANN D R, et al. Site factors determining epiphytic lichen distribution in a dieback-affected spruce-fir forest on Whiteface Mountain, New York: stem flow chemistry[J]. Canadian Journal of Botany,2002,80(11):1131-1140.
[32] CHEN J, HANS-PETER B. Rock-weathering by lichens in Antarctic: patterns and mechanisms[J]. Journal of Geographical Sciences,2002,12(4):387-396.
[33] SHIMIZU A. Community structure of lichens in the volcanic highlands of Mt. Tokachi, Hokkaido, Japan[J]. The Bryologist,2004,107(2):141-151.
[34] RAJAKARUNA N, KNUDSEN K, FRYDAY A M, et al. Investigation of the importance of rock chemistry for saxicolous lichen communities communities of the New Idria serpentinite mass, San Benito County, California, USA[J]. The Lichenologist,2012,44(5):695-714.
Lichen Species Diversity and Community Characteristics of Shirengou in Urumqi County, Xinjiang//
Anwar Tumur, Sayira Xawkat, Nazarbek Guldan, Abdulla Abbas
(Xinjiang University, Urumqi 830046, P. R. China)//
Journal of Northeast Forestry University,2017,45(5):49-55.
Lichen diversity; Community classification; Growth form; Lichen flora
1)国家自然科学基金项目(31660009,31670023)。
艾尼瓦尔·吐米尔,男,1970年12月生,新疆大学生命科学与技术学院,教授。E-mail:anwartumursk@xju.edu.cn。
阿不都拉·阿巴斯,新疆大学生命科学与技术学院,教授。E-mail:abdulla@xju.edu.cn。
2016年12月15日。
Q914.84
责任编辑:任 俐。
