毛竹笋的快速生长对母竹的影响1)
2017-06-05徐超王海湘温国胜胡策梁谢恩黄皓南朱丽娜白尚斌张汝民
徐超 王海湘 温国胜 胡策 梁谢恩 黄皓南 朱丽娜 白尚斌 张汝民
(浙江农林大学,临安,311300)
毛竹笋的快速生长对母竹的影响1)
徐超 王海湘 温国胜 胡策 梁谢恩 黄皓南 朱丽娜 白尚斌 张汝民
(浙江农林大学,临安,311300)
为了探讨竹笋快速高生长与母竹之间的养分输导关系,揭示毛竹快速生长内在规律,试验测定了竹笋快速生长期内不同竹龄器官(叶、枝、秆、蔸根)的主要养分质量分数、冠层(上层、中层和下层)水势、地上器官(叶、枝、秆)的生物量,与同样地同期小年期母竹测得值做比较。结果表明:在竹笋快速生长期,氮元素在母竹叶片中先下降后上升,而蔸根中则先略有升高然后趋于稳定;在叶中,钾元素随竹笋快速生长而迅速升高,而在蔸根中始终保持稳定状态;磷在叶片和蔸根的质量分数均较低。同小年期相比,氮元素在叶、枝、秆、蔸根中的质量分数均较低;磷元素在叶、蔸根的质量分数较低,而在枝、秆中其质量分数要高;钾元素在叶、秆、蔸根中的质量分数高,但在枝中低。在竹笋的快速生长期,各竹龄叶的水势变化总体呈“V”型曲线,从3月中旬到5月中旬,水势有变小趋势,随后有所回升。同小年期相比,林冠各层的水势均大于同期的小年期;在竹笋快速生长期,母竹地上各器官生物量占比相对稳定。同小年期相比,光合器官竹叶生物量占地上器官生物量的7.05%,增加了31.90%。因此,竹笋的快速生长对母竹产生了影响,在快速生长期,母竹通过营养元素的迁移、地上生物量格局的分配,为竹笋的快速生长提供了可能。
快速生长期;幼(母)竹;营养元素;水势;竹笋生物量
毛竹(Phyllostachysedulis)是隶属禾本科刚竹属植物,是我国分布范围最广、面积最大、栽培和利用历史最为悠久、经济价值最高的生态经济竹种,也是森林木竹中用途最多的树种之一,在中国乃至世界竹资源里占据非常重要的地位[1-2]。
毛竹林具有典型的“大小年”之分,即“异龄林”,大年主要完成发笋,小年则主要完成长鞭与换叶[3]。毛竹快速生长期是其重要的生理时期,指从竹笋的出土、长高、抽枝、叶的展开直到高生长的结束、秆的形成这段时间。该过程历时40~60 d,幼竹就可完成高生长,竹林生物量迅速增加。但该时期新竹自身的光合能力比较弱,并且根系系统尚未完全发育成形,其生长发育所需的大部分养分原料来自母竹系统[4]。各龄级母竹为供给幼竹的迅速生长,不断地调整营养元素的流动和生物量分配,使得整个竹林系统物质循环以及能量流动都一直处于不断变化的过程[3-4]。目前,对竹林生态系统的研究主要集中在物质养分的输入和输出[5]、竹林地上各器官的营养元素积累与分布[6-8]、毛竹林的凋落物及其分解[4,9]、各龄竹不同年份地上和地下生物量格局[10-12],以及毛竹林生态系统水汽和碳通量[13-15]的变化。而幼竹的快速生长对母竹营养元素分配、水势变化、生物量格局分配的影响迄今未见报道。因此,探究该时期母竹养分动态特征、水势变化规律及其生物量分配特征,对阐明毛竹快速生长内在规律、调整养分管理措施、提高林分生产力具有重要理论和现实指导意义。
1 试验地概况
本试验地点位于浙江省杭州市临安市青山(30°14′N,119°42′E)的毛竹林生理生态测量站。该地气候类型为亚热带季风。四季分明,雨热同期,年平均气温15.9 ℃,年降水量761~1 780 mm。土壤类型主要为黄壤和黄红壤,试验林为毛竹纯林,林下植物较少,毛竹的年龄为1~6 a,高度9~16 m,胸径8~16 cm,立竹度2 400~3 000株·hm-2,林相较好,大小年明显,经营措施为每年劈灌和小年采伐[16]。
2 材料与方法
2.1 样地设置和采样
在坡度、坡向、立竹度等立地条件基本一致的毛竹林选定3个样地,每个样地面积为50 m×50 m,统计样地毛竹的胸径,并分别统计2龄竹、4龄竹、6龄竹的数量,计算各竹龄的平均胸径,作为标准竹。根据幼竹的生长速度,将该时期分为生长初期(3月20日采样)、高生长开始期(4月1日采样)、高生长期(4月15日、5月3日采样)、高生长结束期(5月20日采样)、展叶期(6月4日采样)、新竹形成期(7月14日采样)7个阶段,分7次采样,并用多露点水势仪(PSYPRO.USA)测定各竹龄不同冠层(上层、中层、下层)的叶片水势,测定方法为:选择完好的叶片,用纸巾轻轻的将叶片的正面和背面擦干净。在擦干净叶片后,选取叶片的待测部位(叶片的背面)用凡士林将探头与叶片密封。依照以上方法将叶片夹好后,在每种竹龄各层毛竹叶片上都连接上3个探头,叶片均随机选取。待其平衡后进行读数,读数时间为06:00—16:00,每两小时测定一次。每次每样地采伐2、4、6龄竹的标准竹各1株,共计63株。
在试验地选取2、4、6龄竹的标准竹砍倒,分秆、枝、叶3个器官称其鲜质量,各器官分别取2 000 g,均分成两份备用;同时将样竹的竹蔸挖起,洗去泥土,取蔸根样品1 000 g备用。然后样品带回实验室,其中一份在80 ℃下烘干至恒质量,测定其含水率,算出不同器官的干质量即生物量。另一份和根的样品在60~70 ℃烘干,粉碎、过60目筛孔,充分拌匀后置于干燥瓶中密封、编号,用于养分元素测定。
研究还分析了该试验林同一样地小年期母竹,为小年8月份采样,方法同上。
2.2 测试方法
含水率采用质量法测定,全氮质量分数采用凯氏定氮蒸馏法测定,全磷质量分数采用磷钒钼黄比色法测定,全钾质量分数采用火焰光度法测定[17]。
2.3 数据处理
使用SPSS16.0进行相关数据分析,采用Sigmaplot 12.5进行作图。
3 结果与分析
3.1 竹笋快速生长期各龄级母竹养分动态变化
3.1.1 各龄级母竹氮、磷、钾质量分数
分析测定了各龄级器官(叶、枝、秆、蔸根)氮、磷、钾三大主要营养元素的质量分数(表1)。结果显示,各器官均表现为氮元素的质量分数最高,钾元素的质量分数次之,磷元素的质量分数最低;同一元素,器官不同,质量分数差异也比较大。具体表现为在叶中氮、钾元素的质量分数较高,分别是21.12、12.29 g·kg-1,远远超过枝、秆、蔸根中氮、钾的质量分数,而磷元素在叶、枝、秆、蔸根的质量分数分别是0.87、0.56、0.55、0.63 g·kg-1,质量分数较氮、钾元素相对稳定;不同器官营养元素质量分数也不同,氮元素、磷元素平均质量分数由大到小的顺序为叶、蔸根、枝、秆,钾元素平均质量分数由大到小的顺序表现为叶、蔸根、秆、枝;竹龄对各个器官的营养元素质量分数有一定的影响。氮元素和钾元素的质量分数由大到小表现为2龄竹、4龄竹、6龄竹,磷元素相对其他两个元素质量分数较低,且在各竹龄间差异较小。
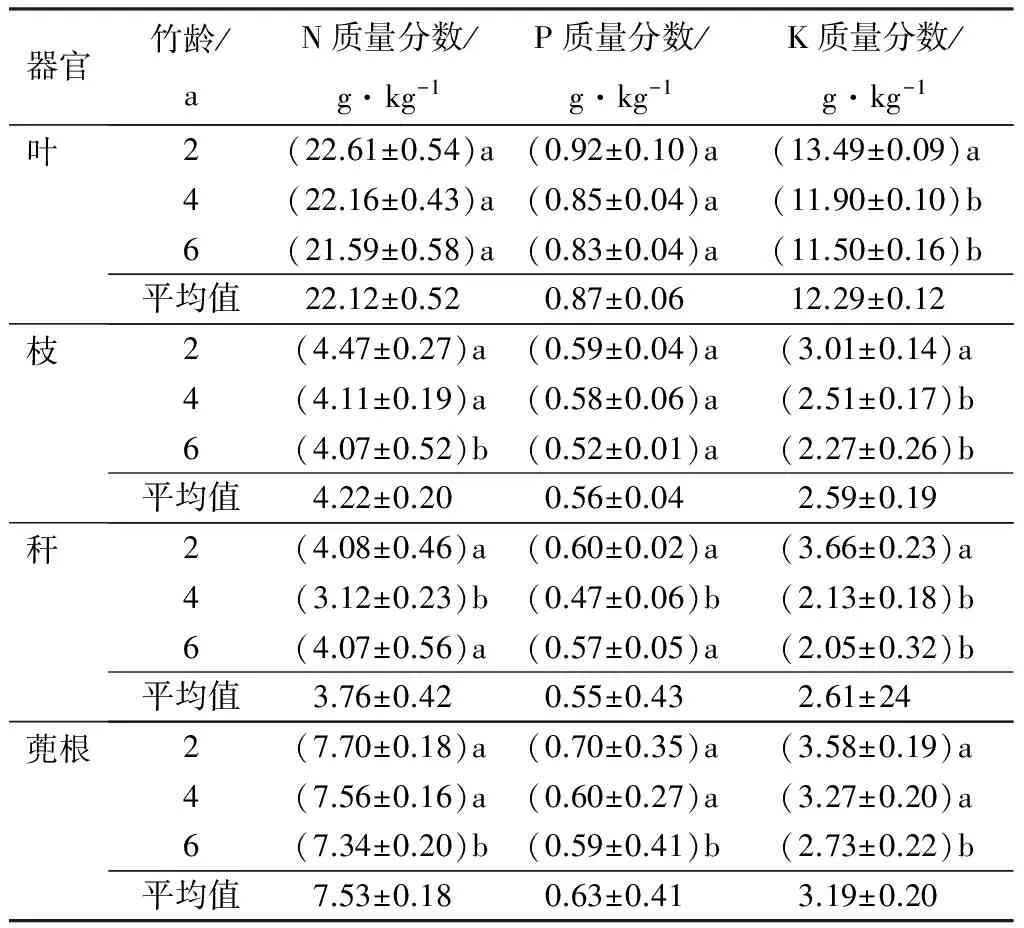
表1 快速生长期母竹各器官氮、磷、钾质量分数
注:表中数据为7次测量的平均值±标准误;同列不同字母表明同一器官不同竹龄的N、P、K质量分数差异性显著(P<0.05)。
3.1.2 叶和蔸根中氮、磷、钾质量分数的动态变化
叶和根分别是光合作用和养分吸收的重要器官,重点分析了叶和蔸根中的氮、磷、钾元素动态变化规律(图1)。从3月中旬到5月中旬,随着幼竹的快速生长,氮的质量分数的在叶中呈现下降趋势,5月中旬达到最小值为20.59 g·kg-1,随后缓慢上升,到6月初达到最大值为24.22 g·kg-1,然后下降;但在同时期的蔸根中,氮元素先呈现上升趋势,随后相对稳定。与此同时,钾元素的质量分数在叶中随幼竹的快速生长总体呈现波动式的逐渐上升趋势;但是在蔸根中钾元素的质量分数在此时期变化不是很大,总体趋于稳定。不同于氮、钾两种元素,磷元素的质量分数在叶和根中质量分数很低,在叶中随着幼竹的快速生长,磷元素总体呈上升的趋势,6月中旬达到最大值为0.87 g·kg-1,然后下降;在蔸根中,磷元素是波动幅度较大,5月份达到最大值。
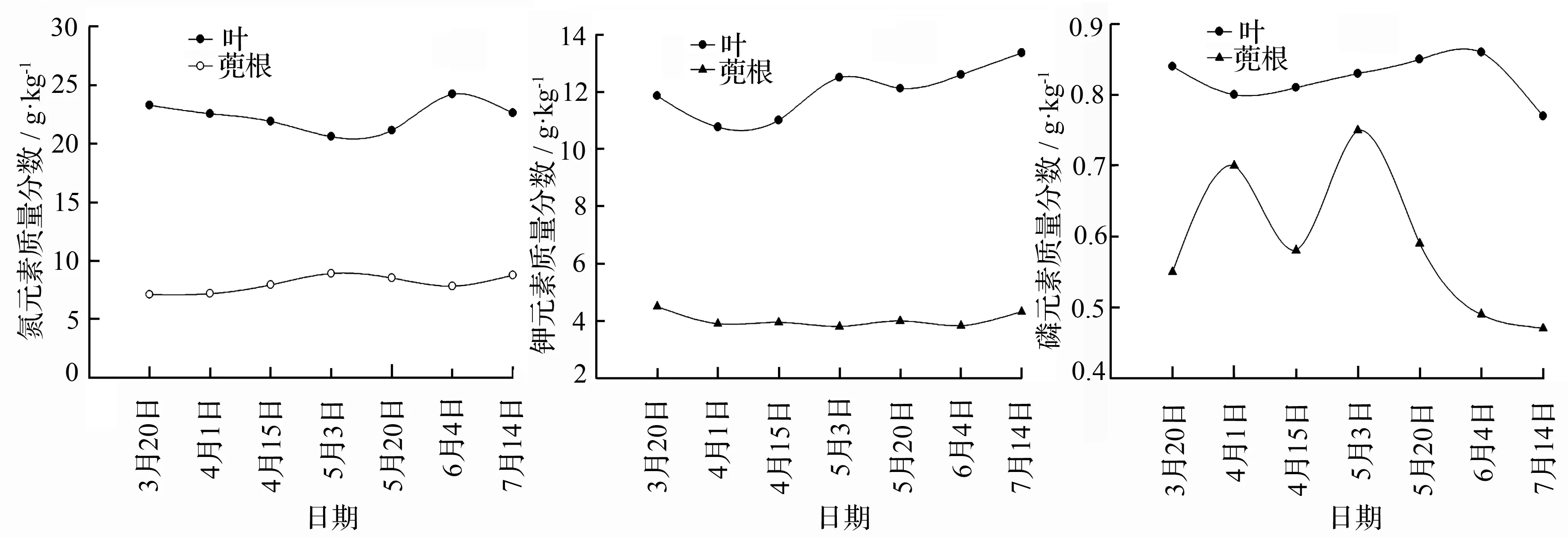
图1 快速生长期母竹叶、蔸根中的养分质量分数动态变化
3.1.3 新竹快速生长对母竹各器官营养元素质量分数的影响
表2分析了小年期和幼竹快速生长期各龄级母竹叶、枝、秆、蔸根中氮、磷、钾3种元素质量分数的差异。结果显示,小年期,氮元素在叶、枝、秆、蔸根中的质量分数比幼竹快速生长期要高;在幼竹快速生长期,叶、蔸根中的磷元素质量分数分别是0.87、0.56 g·kg-1,相对于小年期的质量分数较低,但是在枝、秆中磷元素的质量分数要高于小年期,且叶、枝、秆中磷元素质量分数差异显著;与氮、磷元素不同,在叶、秆、蔸根中钾元素质量分数要高于小年期,分别是12.29、2.61、3.19 g·kg-1,但在枝中钾元素质量分数低于小年期,在叶、枝、蔸根中的钾元素质量分数差异显著。
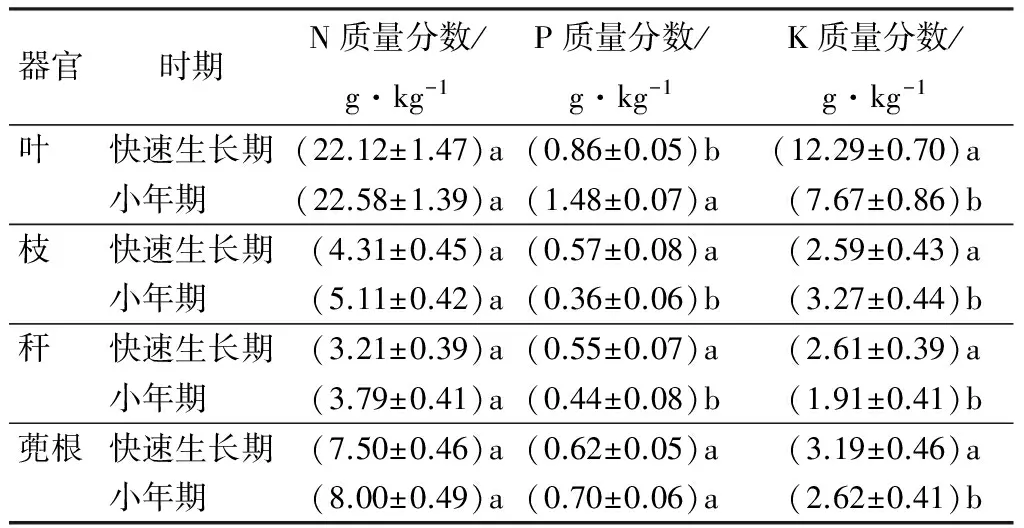
表2 快速生长期和小年期各器官养分元素质量分数的比较
注:表中数据为7次测量平均值±标准误;同列不同字母表示同一器官不同竹龄的N、P、K质量分数差异性显著(P<0.05)。
3.2 春笋快速生长期毛竹叶片水势动态变化规律
3.2.1 各龄级母竹竹叶水势动态变化
测定了各龄竹的不同冠层叶片水势变化(表3),结果显示,从横向水分运输看,各龄竹的叶水势均表现为竹龄越小叶片水势越低,即由小到大表现为2龄竹、4龄竹、6龄竹,根据SPAC(soil-plant-atmosphere continuum)理论,水分运输方向从高水势流向低水势,说明在快速生长期,水分传输方向是从老龄竹流向低龄竹。从垂直方向的水分运输可以得出,相同竹龄的不同冠层的叶片水势由大到小呈现上层水势、中层水势、下层水势的趋势(表3)。各竹龄冠层的水势都呈现出一定的日变化规律并且变幅较大,呈“V”型曲线,在06:00—08:00各竹龄的叶片的水势都很高,随后则渐渐降低,到14:00左右下降至最小值,然后缓慢上升(图2)。在幼竹快速生长期各竹龄的叶水势变化同样呈“V”型曲线,从3月中旬到5月中旬,表现逐渐变小趋势,然后有所回升(图3),此结果与袁佳丽等[18]研究结果一致。

表3 快速生长期各度母竹叶不同冠层的水势
注:表中数据为7次测量的平均值±标准误;同列不同字母表示差异显著(P<0.05)。
3.2.2 新竹快速生长对母竹叶片水势的影响
表4分析了快速生长期和小年期各龄母竹不同冠层(上层、中层、下层)水势的差异。结果表明:毛竹林不同冠层的叶片水势由大到小呈现为下层水势、中层水势、上层水势,在快速增长期,各冠层的叶片水势均大于同期的同冠层的小年期。其中上层差异不显著,中层和下两层差异达到显著水平(P<0.05)。
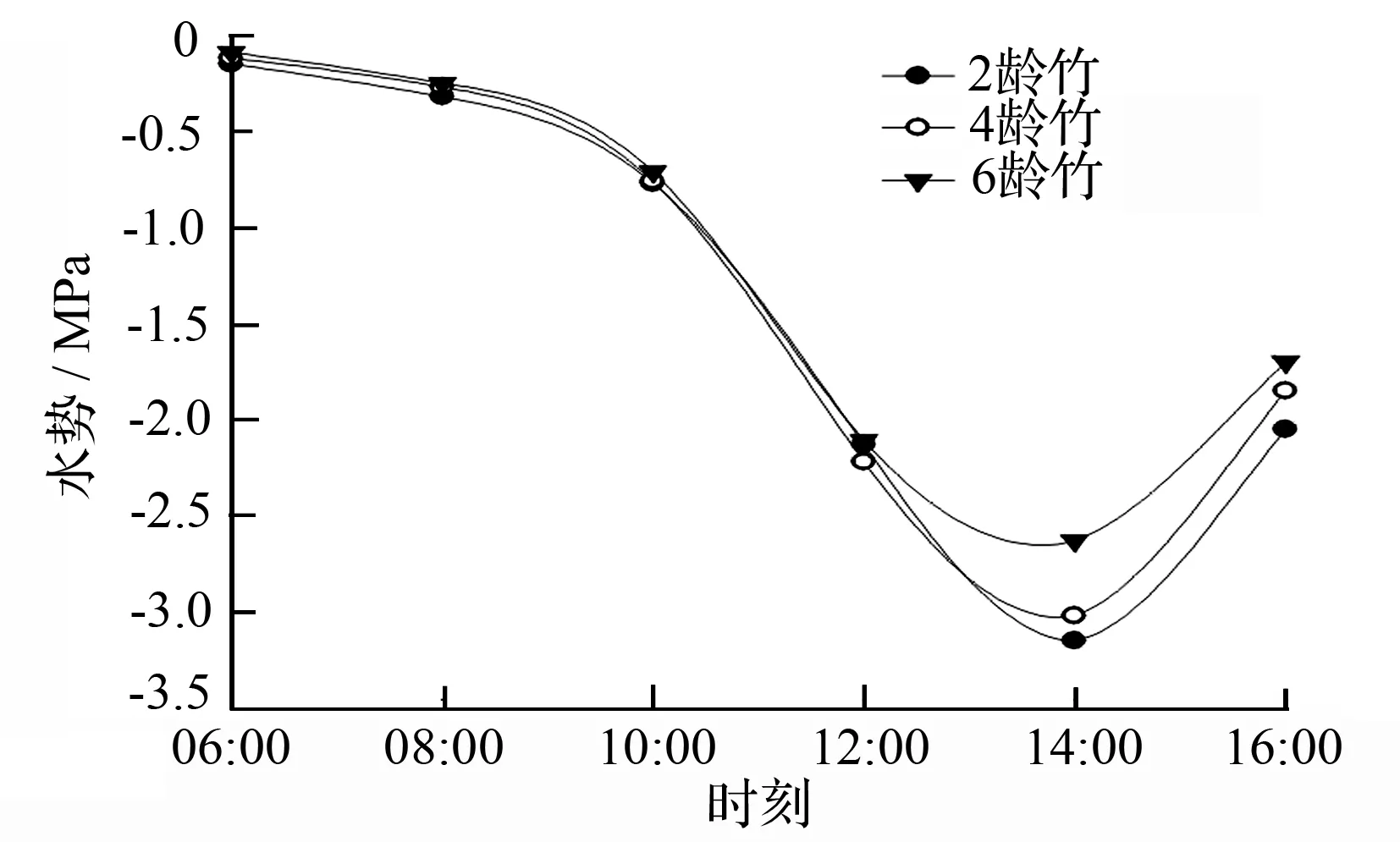
图2 快速生长期各度母竹叶水势日变化
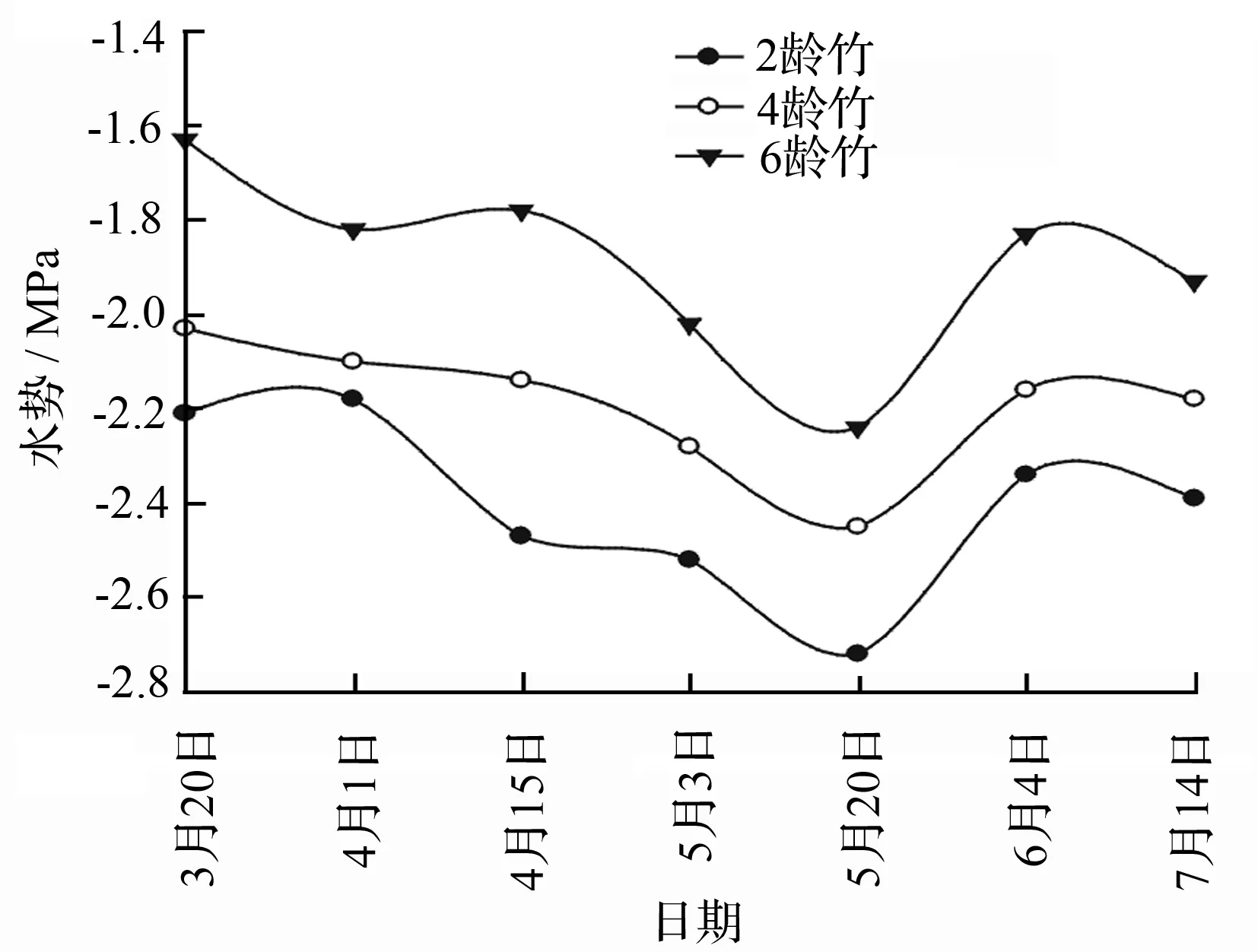
图3 快速生长期母竹叶水势动态变化

表4 快速生长期和小年期母竹叶片不同冠层水势的比较
注:表中数据为7次测量的平均值±标准误;同列不同字母表示差异显著(P<0.05)。
3.3 春笋快速生长期毛竹地上生物量动态变化规律
3.3.1 地上各器官含水率的变化和生物量的动态变化
表5分析了地上部分各龄级不同器官(叶、枝、秆)的含水率。结果表明:不同器官含水率差异较大,各龄级含水率由大到小均表现为叶、秆、枝;同时含水率随着竹龄的不同亦有所不同,2龄、4龄、6龄竹的叶含水率分别是51.1%、54.8%、57.1%,逐渐增大的趋势,而竹枝的含水率是随着竹龄增加而减小。对于竹秆,2龄竹的含水率大于4龄和6龄竹,但4龄、6龄竹的含水率差异不显著。
从母竹的叶、枝、秆的含水率在幼竹快速增长期内的动态变化规律(图4)可以看出,从4月初开始,各竹龄的母竹叶片含水率逐渐下降,直至6月份则降到最小值,这是因为此时期是毛竹的展叶期,竹叶的叶肉组织比较幼嫩,但是此时叶片新陈代谢比较旺盛,耗水较其他时期更多[18],说明新竹迅速生长对各个龄级母竹系统的新陈代谢产生了影响。枝和秆随着幼竹的快速生长其含水率呈现逐渐增大的趋势。
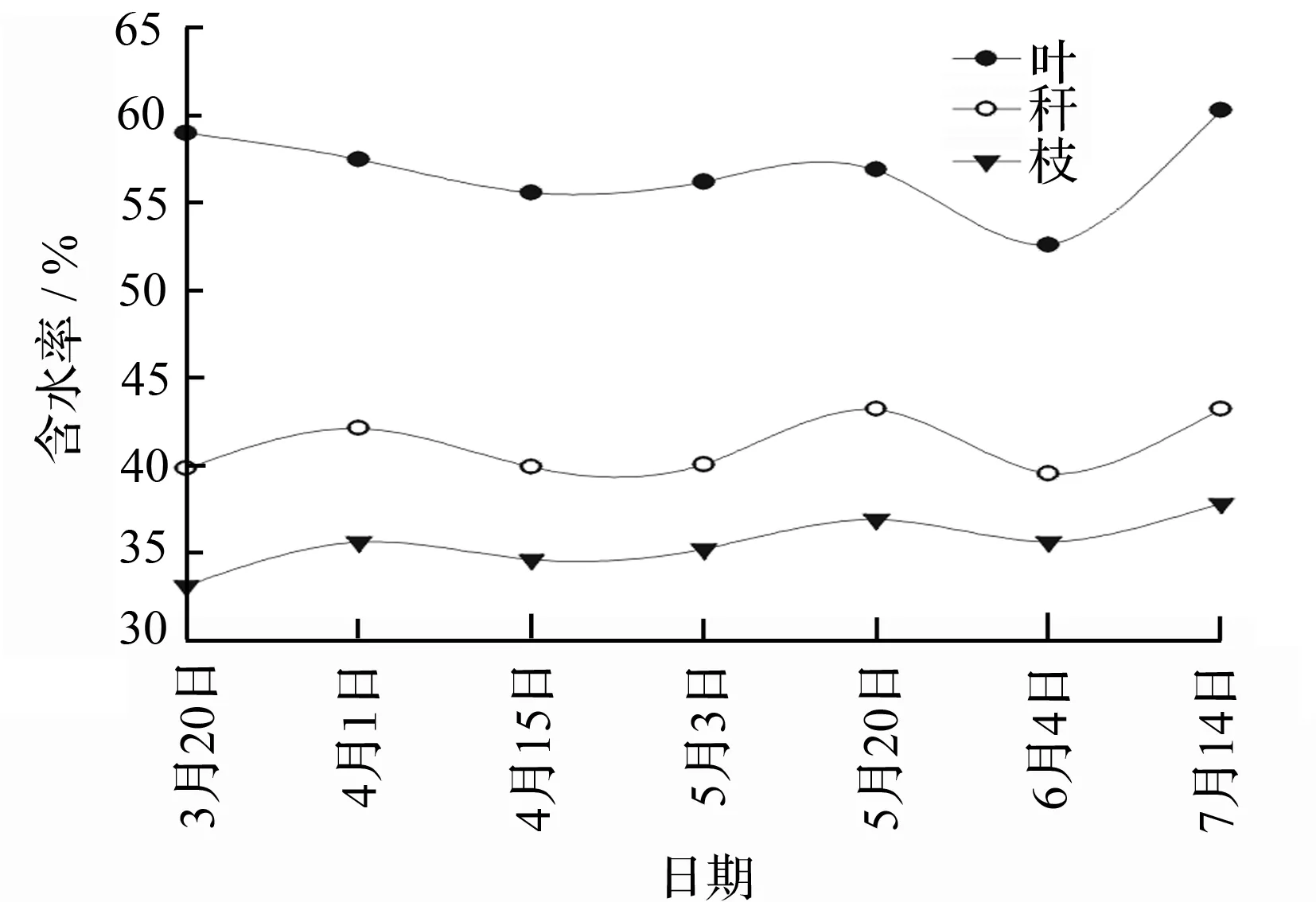
图4 快速生长期母竹器官含水率的动态变化
从幼竹快速生长期,母竹叶、枝、秆的生物量占地上生物量的动态变化趋势(图5)可看出,母竹地上器官生物量格局波动较平缓,没有明显的规律性。
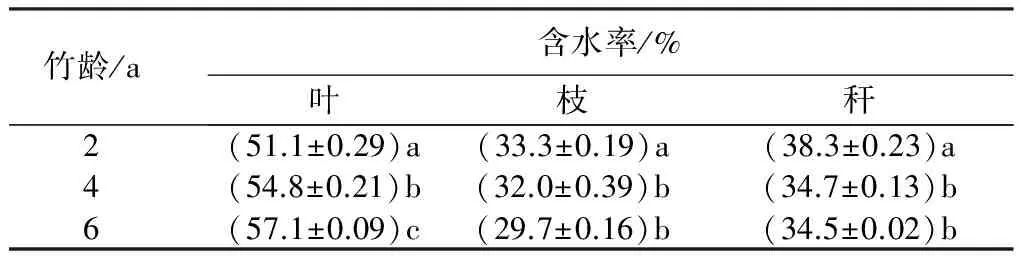
表5 快速生长期母竹器官含水率
注:表中数据为7次测量的平均值±标准误;同列不同字母表示差异显著(P<0.05)。
3.3.2 新竹快速生长对生物量格局的影响
本研究分析了该试验林同一样地新竹快速生长期生物量分配格局,并与小年期进行比较(表6)。结果表明:幼竹快速增长期竹叶生物量占总生物量的百分比是7.05%,而小年期为4.80%,增加了31.90%。方差分析表明,竹叶生物量比例在新竹快速生长期与小年期的差异达到显著水平,而竹秆和竹枝的生物量分配比例无显著差异,为满足该时期新竹生长需求,母竹通过增加竹叶的生物量分配来提高光合作用的利用率。

表6 快速增长期和小年期地上各器官生物量占总生物量百分比
注:表中数据为7次测量的平均值±标准误;同列不同字母表示差异显著(P<0.05)。
4 结论与讨论
营养元素的循环不仅是生态系统最基本功能过程,也是生态系统生产力的决定因素[19]。在毛竹快速生长期,母竹各器官的营养元素质量分数,因年龄不同、器官不同与元素不同而形成差异,不同竹龄各器官均以氮元素的质量分数最高,明显高于磷元素和钾元素,其中竹叶氮元素的质量分数为(20.59~23.27 g·kg-1),高于雷竹(Phyllostachysviolascens)(17.73~22.98 g·kg-1),但低于麻竹(Dendrocalamucatiflorus)(28.6~35.8 g·kg-1)叶中的氮元素质量分数[16],表明毛竹叶片氮的质量分数在竹类植物中位于中间水平,叶中氮、磷、钾三种元素的质量分数在5—6月份迅速上升,可以在毛竹快速生长前期即(3—4月份)进行营养诊断,根据叶片中氮元素质量分数的情况,给予土壤施加氮肥,以供竹笋的生长发育所需,使其健康完成自己的生长过程;氮、磷、钾三种元素的质量分数表现为竹龄越小质量分数越大,研究表明,不同年龄阶段的马占相思(Acaciamangium)有显著的营养元素的转移机制[20],因此,在毛竹快速生长过程中,为满足幼竹的爆发式生长,母竹向幼竹提供大量的营养物质,满足了毛竹快速生长阶段这一重要生理期的养分需求,同时也大大降低了对环境中养分供应的依赖;同小年期相比,质量分数最高的氮元素在叶、枝、秆、蔸根中的质量分数均较低;磷元素在叶、蔸根的质量分数较低,而在枝、秆中其质量分数要高;钾元素在叶、秆、蔸根中的质量分数要高,但在枝中质量分数低,表明在幼竹的快速生长期,幼竹的快速生长影响了母竹各器官的营养物质分配,母竹分配更多的营养元素给予子竹,满足子竹的快速生长所需,使子竹在短短40~60 d迅速完成高生长。
水势是植物体内水分状况的一个重要指标,水势的高低表明,植物从相邻细胞或者土壤中吸收水分用以维持自身正常的生理活动[21]。不同竹龄的冠层叶片水势表现为随竹龄的减小而降低,具有明显的日变化规律,早上水势相对较高,随后水势在一直下降,到14:00左右水势达到最小值,随后又逐渐回升,研究表明,在快速生长期这个重要生理阶段,母竹系统向新竹系统源源不断的输送水分,且随着竹龄增大其输送水分的能力则越强[18],Aster et al.[22]认为,早上植物水势比较高,表明植物此时能很好地从外界环境获取水分以满足自身需求,同时也表明毛竹有较高的水分获取和恢复水分能力,但14:00左右各龄竹的水势下降到最低,甚至已经达到干旱胁迫的程度,则表明此时刻毛竹的蒸腾失水作用较为强烈。同小年期相比,在快速生长阶段,叶片各冠层的水势均大于同期的小年期,表明幼竹的快速生长影响母竹的叶片各冠层的水势。袁佳丽等[18]研究表明,不同竹龄的毛竹在生长中期(5月中旬左右),各竹龄冠层的水势相对于生长前期和生长后期则较低,但是此时期毛竹林土壤水势则较高,这样高的土壤水势和低的叶片水势形成较大的水势梯度差,这样更能满足此时期毛竹自身生长以及蒸腾的耗水。
植物将同化产物分配于各器官形成生物量格局。通过调节不同生理生长阶段各器官的生物量格局来适应外界环境,以保证自身正常生长发育[11-12]。竹龄对地上生物量格局有一定影响。2、4龄竹的竹叶生物量分配比例大于6龄竹,说明2、4龄竹对光竞争的投入相对较大,进而保证了较强的光合能力,这与此前研究的2、4龄竹子的养分积累速度较快的研究结果基本一致[8]。因此,在竹林抚育时,对2、4龄竹的管理应加强;同小年期相比,叶片的生物量在幼竹快速生长期明显增大,表明为了满足新竹生长,竹林的养分更多的投入到叶中,以便提高光合能力。毛竹林一般在小年6月份完成换叶,作为幼竹快速生长期对照的小年期,取样时间均设在8月份,此时间段毛竹林已换叶生长2个月左右,因换叶造成的差异则比较小,因此,幼竹快速生长期,母竹在各器官生物量分配上,竹叶生物量所占比例提升应与满足新竹生长关系密切,为满足新竹生长,在生物量分配上进行了调整与优化。
[1] 江泽慧.世界竹藤[M].沈阳:辽宁科学技术出版社,2002.
[2] 叶松涛,杜旭华,宋帅杰,等.水杨酸对干旱胁迫下毛竹实生苗生理生化特征的影响[J].林业科学,2016,51(11):25-31.
[3] 毛超,漆良华,刘琦蕊,等.毛竹林各器官对N素的吸收和利用率[J].林业科学,2016,52(5):64-70.
[4] TU L H, HU H L, HU T X, et al. Litterfall, litter decomposition, and nutrient dynamics in two subtropical bamboo plantations of China[J]. Pedosphere,2014,24(1):84-97.
[5] 杨清培,欧阳明,杨光耀,等.竹子生态化学计量学研究:从生物学基础到竹林培育学应用[J].植物生态学报,2016,40(3):264-278.
[6] 吴家森.不同年份毛竹营养元素的空间分布及与土壤养分的关系[J].林业科学,2005,41(3):171-173.
[7] 刘广路,范少辉,官凤英,等.不同年龄毛竹营养器官主要养分元素分布及与土壤环境的关系[J].林业科学研究,2010,23(2):252-258.
[8] NIRMALA C, MADHO S B, HAORONGBAM S. Nutritional properties of bamboo shoots potential and prospects for utilization as a health food[J]. Comprehensive Reviews in Food Science and Food Safe,2011,10(3):153-169.
[9] 刘广路,范少辉,官凤英,等.毛竹凋落叶组成对叶凋落物分解的影响[J].生态学杂志,2011,30(8):1598-1603.
[10] ARUNJN,GITASREED,ASHESHKD.AbovegroundstandingbiomassandcarbonstorageinvillagebamboosinNorthEastIndia2009BiomassandBioenergy[J].BiomassandBioenergy,2009,33(9):1188-1196.
[11] CHEN X G, ZHANG X Q, ZHANG Y P, et al. Changes of carbon stocks in bamboo stands in China during 100 years 2009 Forest Ecology and Management[J]. Forest Ecology and Management,2009,258(7):1489-1496.
[12] LEE J M. Comparing aboveground carbon sequestration between moso bamboo (Phyllostachysheterocycla) and China fir (Cunninghamialanceolata) forests based on the alometric model[J]. Forest Ecology and Management,2011,261(6):995-1002.
[13] 刘玉莉,江洪,周国模,等.安吉毛竹林水汽通量变化特征及其与环境因子的关系[J].生态学报,2014,34(17):4900-4901.
[14] 孙成,江洪,陈健,等.亚热带毛竹林生态系统能量通量及平衡分析[J].生态学报,2015,35(12):4128-4136.
[15] 徐超,温国胜,王海湘,等.毛竹快速生长期的高生长与碳通量的变化规律[J].东北林业大学学报,2016,44(11):1-4.
[16] 曾莹莹,王玉魁,蔡先锋,等.毛竹林爆发式生长期立竹器官营养成分的动态变化[J].浙江农林大学学报,2015,32(2):272-277.
[17] 鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000:308-316.
[18] 袁佳丽,温国胜,张明如,等.毛竹快速生长期的水势变化特征[J].浙江农林大学学报,2015,32(5):722-728.
[19] 黄张婷,姜培坤,宋照亮,等.不同竹龄雷竹中硅及其他营养元素吸收和积累特征[J].应用生态学报,2013,24(5):1347-1353.
[20] 何斌,秦武明,余浩光,等.不同年龄阶段马占相思(Acaciamangium)人工林营养元素的生物循环[J].生态学报,2007,27(12):5158-5167.
[21] 林琭,汤昀,张纪涛,等.不同水势对黄瓜花后叶片气体交换及叶绿素荧光参数的影响[J].应用生态学报,2015,26(7):2030-2040.
[22] ASTER G, DEMEL T, MASRESHA F, et al. Adaptation of five co-occurring tree and shrub species to water stress and its implication in restoration of degraded lands[J]. Forest Ecology and Management,2006,229(1/2/3):259-267.
Effects of Fast-growing ofPhyllostachysedulison the Mother Bamboo//
Xu Chao, Wang Haixiang, Wen Guosheng, Hu Ce, Liang Xieen, Huang Haonan, Zhu Lina, Bai Shangbin, Zhang Rumin
(Zhejiang Agriculture and Forestry University, Lin’an 311300, P. R. China)//
Journal of Northeast Forestry University,2017,45(5):11-15,19.
The experiment was conducted to study the relationship between the rapid growth of bamboo shoots and nutrient transport between mother bamboos, and reveal the inherent law of rapid growth of bamboo. The main nutrient content, water potential of canopy and biomass of aboveground organs were measured in different growth stages of bamboo shoots during the fast growth period, and compared with the corresponding value mother bamboo in the same period. In the rapid growth period of bamboo shoots, N content in the leaves of the mother bamboo decreased first and then increased, but in the root, it increased first and then stabilized, K increased rapidly with the rapid growth of bamboo shoots, and remained stable in the roots, P in leaves and roots were low. Compared with the off-year period, the contents of N in leaves, branches, culms and roots were lower, the contents of P in leaves and roots were lower, while those in branches and culms were higher, and K in leaves, culms and roots was higher, but lower in branches. During the rapid growth period of bamboo shoots, the leaf water potential of each bamboo age showed a “V”-shaped curve. From mid-March to mid-May, the water potential became smaller and then picked up. Compared with the off-year period, the water potential of canopy layers was greater than that in the off-year period. During the rapid growth period of bamboo shoots, the biomass of the organs on the ground was relatively stable. Compared with the off-year period, bamboo biomass accounted for 7.05% of the aboveground organs biomass, increased by 31.9%. Therefore, the rapid growth of bamboo shoots had an important impact on the mother bamboo, and the rapid growth of bamboo shoots had an effect on the mother bamboo. In the rapid growth period, the distribution of nutrients and the allocation of aboveground biomass provided the rapid growth of bamboo shoots.
Fast-growing period; Young (mother) bamboo; Nutrient elements; Water potential; Bamboo shoot biomass
1)国家自然科学基金项目(31270497,31570686,31170594)、浙江省与中国林业科学研究院省院合作项目(2014SY16)、浙江省大学生科技创新活动计划暨新苗人才计划项目(2016R412045)。
徐超,男,1986年7月生,浙江农林大学林业与生物技术学院,硕士研究生。E-mail:nmweifan@126.com。
温国胜,浙江农林大学林业与生物技术学院,教授。E-mail:wgs@zafu.edu.cn。
2016年12月20日。
S718.5;Q949.71+4.2
责任编辑:任 俐。
