玉米抗病毒病基因工程研究进展
2017-05-30燕照玲段俊枝冯丽丽陈海燕齐红志杨翠苹施艳任银玲刘毓侠
燕照玲 段俊枝 冯丽丽 陈海燕 齐红志 杨翠苹 施艳 任银玲 刘毓侠
摘要:病毒病是导致玉米产量降低和品质下降的主要原因之一。在我国,玉米矮花叶病和粗缩病发生范围最广、危害最严重。基因工程技术可人为将抗性基因或部分片段定向导人植物获得转基因抗病毒植株,具有速度快、效率高等优点,在玉米抗病毒育种中具有重要的应用价值。文章在总结我国主要玉米病毒病及其病原种类的基础上,论述采用不同策略培育抗病毒玉米植株的研究进展,其中,利用植物病毒基因序列的策略有病毒编码蛋白基因介导的抗病性、RNA干扰(RNAi)介导的抗病性、人工小RNA(amiRNA)介导的抗病性3种,还可利用非植物病毒基因,包括寄主的抗性基因及来自其他植物、动物和微生物的抗病毒基因如核糖体失活蛋白、核酸酶、2-5A体系的基因等;最后分析各种策略的优缺点及抗病毒转基因玉米的安全性问题,为科研工作者优化玉米抗病毒育种工程、培育生产上可推广的抗病毒品种提供参考。
关键词:玉米;病毒病;甘蔗花叶病毒;水稻黑条矮缩病毒;基因工程;抗病育种
中图分类号:S435.131 文献标志码:A 文章编号:2095-1191(2017)12-2136-09低和品质下降的主要原因之一,且呈发病面积逐年扩大、病情逐年加重的趋势,绝产情况时有发生。由于生产上缺乏行之有效的药剂和防治措施,病毒病已成為限制玉米持续增产的主要障碍。培育、推广抗病毒品种并辅以合理的栽培管理措施是目前公认防治病毒病的最佳途径,但玉米种质资源对病毒病抗性的遗传基础较复杂,进而给通过传统杂交利用玉米抗性基因培育抗病品种带来一定难度。随着组织培养和基因工程技术的发展,可人为将抗性基因或部分片段定向导入植物获得转基因抗病毒植株,该技术具有速度快、效率高等优点,在玉米抗病毒育种中的应用研究已取得了重大进展。本文通过介绍我国主要的玉米病毒病种类及其病原,重点论述采用不同策略培育抗病毒玉米植株研究进展,为科研工作者优化玉米抗病毒育种工程提供参考。
1我国玉米病毒病主要种类及其病原
玉米在其整个生长期均可被病毒感染,产生叶片褪绿、枯斑、雌雄穗不育、植株矮化、坏死或直接死亡等症状,最终导致产量降低、品质下降。据不完全统计,全世界有40多种玉米病毒病,在我国发生的主要有粗缩病、矮花叶病、条纹矮缩病、红叶病和鼠耳病等,其中矮花叶病和粗缩病发生范围最广、危害最严重(李秀坤等,2015),因此,这两种病毒病是我国科研工作者研究的重点。
1.1玉米矮花叶病及其病原
矮花叶病是世界玉米产区普遍发生的一种病毒病,1963年首先在美国俄亥俄州被发现。我国于1968年首次在河南新乡和安阳地区发现该病,随后华北各省(市)陆续发生,目前在东北、西南及西北各省(区)均有报道。矮花叶病已成为我国玉米产区的主要病害之一。发病植株通常叶片不均匀褪绿,形成花叶、条纹症状,病情严重时植株矮化,雌雄穗发育受抑制,最终导致玉米产量和质量受到严重影响。此外,玉米矮花叶病明显表现出暴发性、迁移性和间歇性三大特征,每年给玉米生产造成20%~80%的损失(张超等,2017)。
玉米矮花叶病由一至多种病毒系统性侵染引起,国际上已报道的病毒有6种,均为马铃薯Y病毒属(Potyvirus)成员,分别是玉米矮花叶病毒(Maize dwarf mosaic virus,MDMV)、甘蔗花叶病毒(Sugar-cane mosaic virus,SCMV)、玉米属花叶病毒(Zea mosaic virus,ZeMV)、高梁花叶病毒(Sorghum mo-saic virus,SrMV)、约翰逊草花叶病毒(Johnson-grass mosaic virus,JGMV)和白草花叶病毒(Penni-setum mosaic virus,PenMV)。据报道,我国玉米矮花叶病的病原有SCMV(曾被认为是MDMV-B或SC-MV-MDB)(蒋军喜等,2003)和PenMV(曾被命名为MDMV-G)(Fan et al.,2003),目前SCMV发生普遍,而PenMV的发生较少,且多限于山西省和河北省(张超等,2017)。各病毒基因组均为单分子正义单链RNA(ssRNA),全长约10 kb,核酸5′端连接一个基因组连接蛋白(VPg),3′端具有一个poly(A)尾,中间是一个大的开放阅读框,通过多聚蛋白加工和移码翻译策略产生11个具有不同功能的成熟蛋白,从N端到C端依次为第一蛋白(P1)、辅助成分—蛋白酶(HC-Pro)、第三蛋白(P3)、P3N-PIPO(位于P3阅读框内部)、第一个6K蛋白(6K1)、圆柱状内含体蛋白(CI)、第二个6K蛋白(6K2)、VPg、核内含体蛋白a-蛋白酶(NIa-Pro)、核内含体蛋白b(NIb)及外壳蛋白(CP)(Revers and Garcia,2015)。
1.2玉米粗缩病及其病原
粗缩病也是玉米生产上的主要病毒病害,具有暴发性、流行性和毁灭性的特点。该病于1949年首次在意大利种植的美国玉米材料中被发现,目前遍布世界大多数玉米产区。在我国,粗缩病于20世纪50年代首次在新疆和甘肃被发现,70年代和90年代一度在华北和西北等地区暴发。近年来,由于气候变化及种植结构调整,粗缩病在我国的发生逐渐加重,特别是在黄淮海夏玉米区套播或晚春播、早夏播玉米上危害严重,其中山东、江苏、辽宁、安徽和河南等省的玉米生产均因粗缩病暴发而遭受重大损失。玉米幼苗期感病,生长延缓,节问变得粗短,植株严重矮化,同时叶背出现白色蜡泪状脉突,叶片变短、僵直、表现深绿;生育后期植株常不能抽穗,或穗异常、结实少,严重降低产量(张晓婷等,2011)。
目前已报道能引起玉米粗缩病的病毒主要有4种,即玉米粗缩病毒(Maize rough dwarf virus,MRDV)、马德里约祠托病毒(Mal de Rio Cuarto virus,MRCV)、水稻黑条矮缩病毒毒(Rice black-streaked dwarf virus,RBSDV)和南方水稻黑条矮缩病毒(Southern rice black-streaked dwarf virus,SRBSDV),均属于斐济病毒属(Fijivirus)(张晓婷等,2011)。美国和欧洲的玉米粗缩病病原主要是MRDV;MRCV主要分布于南美洲;我国玉米粗缩病的病原主要是RBSDV和SRBSDV,其中RBSDV在全国普遍发生,尤其是北方玉米种植区,其传播介体为灰飞虱,而SRBSDV主要在南方发生,由白背飞虱传播(章松柏等,2013)。RBSDV基因组由10条双链RNA(dsRNA)组成,总长29142 bp,根据片段由大到小依次命名为S1~S10。其中,S1编码RNA依赖RNA聚合酶,S2编码主要核心结构蛋白,S8和s10分别编码核心衣壳蛋白和外层衣壳蛋白(邓金奇和戴良英,2012)。SRB-SDV是周国辉等(2010)在我国南方地区水稻上发现的新病毒,后来发现其也侵染玉米,引发典型的粗缩病症状,其基因组结构与RBSDV相似,但在序列上存在明显差异,各片段核苷酸序列相似性均低于80%。
2玉米抗病毒基因工程
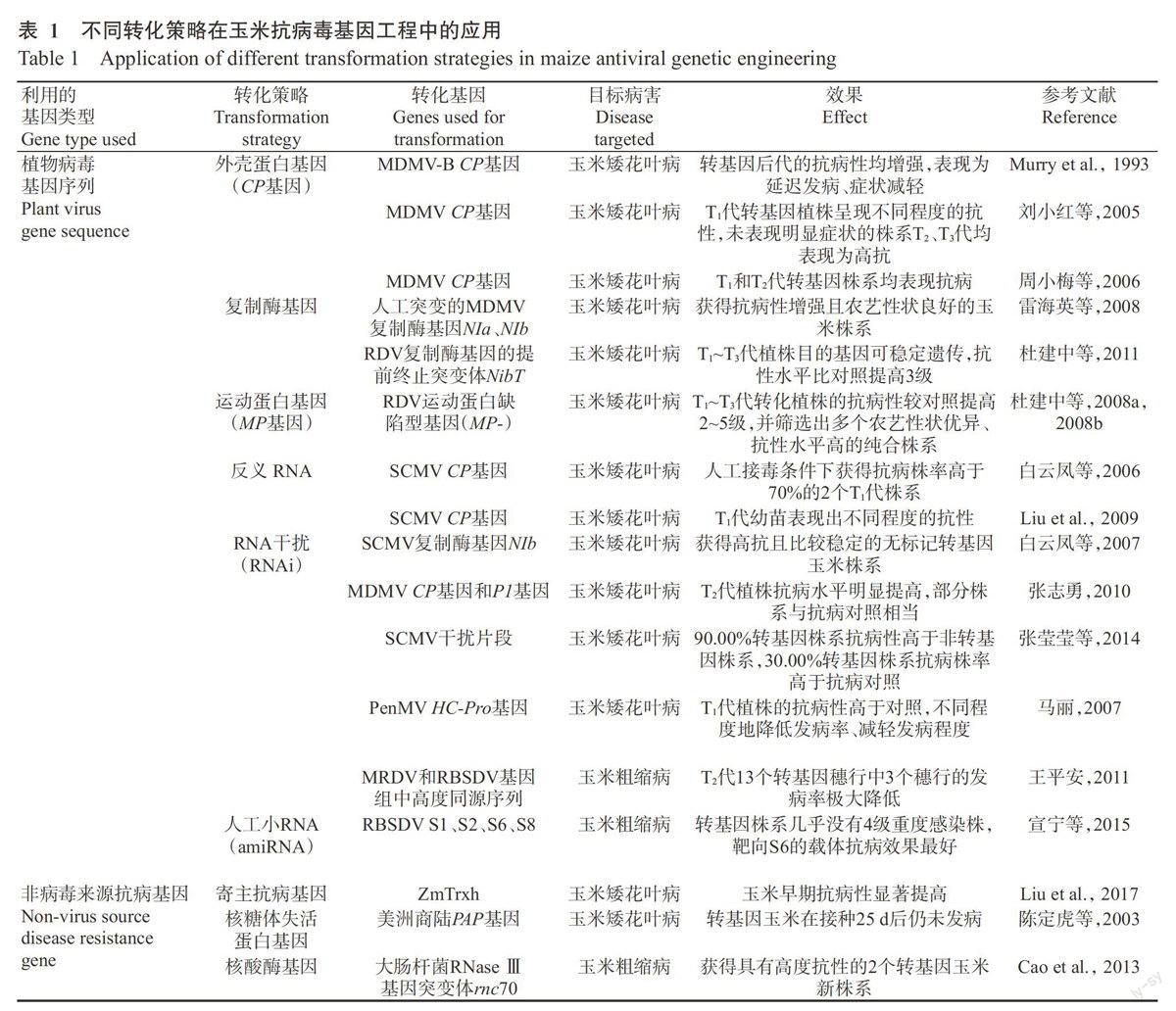
在玉米抗病毒基因工程研究中,用于转化的基因可分为两类:一类是植物病毒的基因序列,如CP基因、复制酶基因、运动蛋白(MP)基因及反义RNA等;另一类是非植物病毒基因,包括来自植物、动物和微生物的抗病毒基因及寄主的抗性基因等。如表1所示,国内外学者通过将两类基因转化玉米获得大量阳性植株,且大多数植株的抗病性提高。
2.1利用植物病毒基因序列
自Sanford和Johnston(1985)提出来源于病原抗性的概念后,利用病毒来源的基因转化植物逐渐成为抗病毒基因工程的重要途径。第1代抗病毒转基因策略主要通过表达病毒自身蛋白基因来实现。第2代抗病毒转基因策略基于RNAi技术,通常是将与病毒基因同源的dsRNA或发卡RNA(Hairpin RNA,hpRNA)转入植物体内,产生病毒特异性的小干扰RNA(Small interfering RNA,siRNA)后,引起入侵的同源病毒基因发生沉默。第3代抗病毒转基因策略是基于amiRNA干扰技术,能有效避免脱靶效应和重组病毒的产生,具有抗性水平高、遗传稳定性强、生物安全性高等优点(FaNm and LarMn,2013)。
2.1.1第1代抗病毒轉基因策略 该策略主要是将植物病毒编码蛋白的基因转入植物细胞,其表达后可使转基因植株获得抗病毒能力。玉米抗病毒基因工程研究中利用的病毒基因主要包括CP基因、复制酶基因、MP基因及反义RNA,转化植株后均取得较好的抗病效果,其中CP基因研究得最早,应用也最多。
(1)CP是形成病毒颗粒的结构蛋白,其主要功能是包被病毒基因组核酸及参与病毒的长距离运输。CP基因介导的抗性机制较复杂,早期研究认为是由于CP蛋白的积累封闭病毒粒子脱壳,从而阻碍病毒侵染,后来逐渐发现存在着不依赖于CP蛋白本身的抗性机制,因而抗性水平也可能发生在RNA水平上(Reimann-Philipp and Beachy,1993)。Murry等(1993)首次报道,通过将MDMV-B的CP基因导入玉米得到抗病植株,人工接种MDMV-A株系、MDMV-B株系鉴定,转基因后代的抗病性均较非转基因对照增强,表现为延迟发病、症状减轻。之后CP基因介导的抗玉米病毒病策略得到科学界的广泛关注,国内也进行了积极探索。刘小红等(2005)采用MD-MV CP基因转化玉米优良自交系18-599红和18-599白,人工接种病毒后,对照植株全部表现出玉米矮花叶病的典型症状,而T1代转基因植株呈现不同程度的抗性,其中有4个植株未表现明显症状;对这4个株系后代进一步接种病毒鉴定发现,T2和T3代均表现为高抗玉米矮花叶病,表明转MDMV CP基因获得的抗病性能稳定遗传。周小梅等(2006)在玉米自交系综3和综31中转入MDMV CP基因,也获得了可育转基因植株。在大田抗病性鉴定试验中,综31的T1和T2代转基因株系均表现出抗玉米矮花叶病,大部分植株的发病率和病情指数低于对照(T1代植株比例分别为83%和67%,T2代比例为100%和90%);温室抗病性鉴定试验中,综3的18个T1代转基因株系有7个抗MDMV侵染,不仅发病延迟,其株高和穗形成率还明显高于对照。接种病毒后,综3对照的可溶性糖含量和超氧化物歧化酶(SOD)、苯丙氨酸解氨酶(PAL)、过氧化物酶(POD)活性均下降,而转基因植株体内的可溶性糖含量有所上升,SOD、PAL活性变化不大,POD活性高于对照,过氧化氢酶(CAT)活性变化也存在差异(韩玉杰,2009)。
(2)复制酶是催化病毒合成正、负链RNA的RNA聚合酶。复制酶基因介导的抗病性机制可能是转基因表达的复制酶反向调控病毒复制,使其复制效率降低,如果表达缺陷型复制酶,其还可能与野生型病毒复制酶产生竞争而干扰病毒复制(Carr et al.,1992;杜建中等,2011)。张志燕(2004)将RBSDVS1部分片段转化玉米,获得了可育再生植株,在T1代植株中检测到外源基因,并且植株形态和农艺性状表现正常,为筛选抗粗缩病玉米株系奠定了基础。雷海英等(2008)将人工突变的MDMV复制酶基因(NIa、NIb)转入4种不同玉米自交系中,对转化的植株连续2代(T1和T2)利用人工摩擦接种病毒的方法进行田问抗病性鉴定,获得了对玉米矮花叶病抗病性增强且农艺性状良好的玉米株系。从T2代整体的发病率来看,4种材料与其相应的对照材料相比均表现出较高的抗性,其中金黄96B、478两种转基因材料较金黄96C、C649两种材料的抗性强,可能与玉米材料、转化基因及外源基因整合位点等不同有关。杜建中等(2011)将水稻矮缩病毒(Rice dwarf virus,RDV)复制酶基因的提前终止突变体(NibT)导人玉米自交系金黄96B,对T1~T3代转基因植株进行鉴定,发现目的基因可稳定遗传,田问对玉米矮花叶病的抗性水平各代基本一致,比对照提高3级,同时穗长、穗粒数、百粒质量等农艺性状也有所改善。而林美娟等(2005)认为复制酶基因介导的抗病性比CP基因介导的抗病性更有效,然而由于该策略对植物病毒种类具有较高的特异性,因此其应用范围可能会受限。
(3)MP介导的病毒抗性策略一般采用缺陷型MP。抗病机理可能表现在蛋白质水平上,转基因所表达的缺陷型MP与入侵病毒的正常MP竞争寄主胞问连丝上的结合位点,阻碍病毒在细胞间的移动(王关林和方洪筠,2002)。Carrington等(1996)研究认为,许多不同病毒的MP可能与相同的胞间连丝成分发生作用,因而该策略的抗病效果具有广谱高效性。杜建中等(2008a,2008b)将RDV运动蛋白缺陷型基因(MP-)导入玉米自交系478,田间接种MDMV进行调查,结果发现转化植株的抗矮花叶病能力与对照植株相比有显著提高,个别株系在T1代的发病率为0,T1~T3代转化植株的抗病性逐渐提高,较对照提高2~5级。结合对优异农艺性状的筛选,获得96C0502、96C0507和96C0513等抗矮花叶病转基因玉米纯合株系,这些株系连续2年的发病率均为0,抗病性比对照提高4级。以上研究表明,利用病毒MP-基因转化玉米是获得抗矮花叶病玉米株系的快捷途径。进一步对RDV MP-转基因玉米的生理特性进行研究,结果发现转基因改变了玉米同工酶的酶谱,第1真叶中POD、CAT和SOD的活性显著高于非转基因玉米,表明该基因导入能够加快受体基因型变异速率,从而增强其抗逆能力和适应性(燕平梅等,2012)。
(4)反义RNA是一类与mRNA互补的ssRNA,其通过与靶向mRNA配对产生dsRNA,阻止mRNA翻译成蛋白质或导致mRNA降解,从而阻断病毒侵染(牙库甫江·阿西木等,2015)。Liu等(2009)将SCMVCP基因以反义方向导人玉米自交系18-599红,对T1代种子长出的幼苗接种SCMV-MDB,发现其对病毒表现出不同程度的抗性。白云凤等(2006)将SCMV反义CP基因导入玉米自交系综3,人工接毒条件下获得抗病株率高于70%的2个T1代株系,与转MD-MV或SCMV CP基因获得的结果相比,转反义CP基因后代的抗病株率高、抗病性强,说明转反义CP基因也是玉米获得矮花叶病抗性的一条有效途径。反义RNA不会翻译成蛋白质与入侵病毒进行异源包装,也不会与入侵病毒发生遗传物质重组,因而该策略与正义RNA相比具有更高的生物安全性。
2.1.2第2代抗病毒转基因策略 RNAi是植物体内天然存在的抗病毒机制。基于RNAi的抗病毒转基因策略是人为强化植物的这种天然机制,选择病毒基因组中的某段序列设计成双链结构转入植物进行表达,其会被植物体内的Dicer(一种RNA酶)降解成21~25 nt的siRNA,然后siRNA与植物的AGO蛋白结合形成RNA诱导的沉默复合体(RNA-induced si-lencing complex,RISC),其在siRNA指导下将互补的病毒RNA降解,从而阻止病毒入侵(牙库甫江·阿西木等,2015)。RNAi介导的抗病性策略特异性强,较易获得抗性高的植株,且抗病性能稳定遗传,同时避免病毒蛋白翻译、病毒RNA重组及异源包装等潜在风险,与传统抗病毒基因工程相比具有更高的安全性(白云凤等,2007)。因此,RNAi技术为培育抗病毒植株提供了一条高效途径,成为玉米抗病毒基因工程研究的重要策略。
白云凤等(2007)构建SCMV复制酶基因(NIb)的反向重复序列表达载体并转化玉米自交系,对T1和T2代植株接种病毒进行鉴定,获得对SCMV高抗且较稳定的无标记转基因玉米株系,同时发现其抗病株率和抗病程度均高于传统的转正义基因和反义基因。张志勇(2010)分别选取MDMV CP基因和P1基因的保守序列构建RNAi表达载体,之后转化玉米自交系18-599,對T2代转基因植株接种病毒发现,其抗病水平明显提高,部分株系与抗病对照H9-21相当。河南农业大学玉米种质资源创新与高产机制研究课题组将人工合成的SCMV干扰片段构建成RNAi载体,以HiⅡ、HiⅡA、HiⅡB、H99的幼胚为受体材料进行转化获得了转基因株系,对T1和T2代接种SCMV进行鉴定发现,90.00%转基因株系抗病性高于非转基因株系,30.00%转基因株系抗病株率高于抗病对照黄早4,转基因株系的抗病性明显提高,说明利用RNAi技术培育抗矮花叶病的转基因玉米株系具有可行性(张莹莹等,2014)。对于PenMV引起的玉米矮花叶病,马丽(2007)也尝试采用RNAi策略进行抗病基因工程研究,其根据PenMV HC-Pro基因保守序列设计引物构建了反向重复序列表达载体并转化玉米,经人工接种PenMV鉴定,转基因T1代植株的抗病性高于对照,表现为发病率降低、发病程度减轻。
抗玉米粗缩病研究中,郭靖(2011)分别选择MRDV S10和RBSDVS10、S8保守区域中的303、452和291 bp片段作为RNAi靶序列,构建成具有反向重复结构的植物表达载体后转化玉米自交系18-599,获得了阳性转化植株。王平安(2011)选取MRDV和RBSDV基因组中高度同源的一段558 bp序列作为干扰序列,转化玉米HiⅡ幼胚后,对T2代采取人工接种饲毒灰飞虱方法进行田间抗性鉴定,结果发现非转基因穗行发病率为57.1%,13个转基因穗行中3个穗行的发病率相对较低(0~15.8%),被初步鉴定为转基因抗粗缩病株系。
为了获得兼抗矮花叶病和粗缩病的转基因玉米植株,可构建同时含有2种病毒干扰片段的复合表达载体。王迅等(2010)分别将RBSDV S8、S10部分片段与SCMV CP基因的一段融合,通过正反向插入植物表达载体构建2种病毒来源基因的RNAi载体,将其导人玉米自交系A188中,部分阳性转基因植株对玉米矮花叶病具有明显的抗性。甘德芳(2011)分别扩增SCMV和MRDV的CP基因(MRDV为S10)特异性干扰片段,构建成反向重复序列并串联在一起得到RNAi复合表达载体,通过转化玉米获得了整合有2种病毒CP基因RNAi片段的转化株。对T2代4叶期玉米接种SCMV后,不同转化株系的抗病性存在差异,但均高于对照。至今,有关二价转基因玉米植株对粗缩病的抗性尚无报道,下一步需要对获得的转基因材料进行粗缩病抗性检测,以验证采用RNAi策略抗多种病毒的效果及可行性。
2.1.3第3代抗病毒转基因策略 amiRNA抗病毒策略是以植物内源前体miRNA(Pre-miRNA)为骨架,将其茎环结构中的miRNA序列替换为与病毒基因序列互补的amiRNA序列形成pre-amiRNA,其转入植物后在内源miRNA合成机制作用下生成靶向病毒mRNA的amiRNA,指导RISC降解病毒mRNA(Tiwari et al.,2014)。利用amiRNA策略培育抗病毒植株已在许多植物和病毒组合中被证明有效,其在玉米抗病育种方面的潜力也引起了广泛的重视。宣宁等(2015)根据玉米zea-miR159a的前体序列和RBSDV基因组序列信息设计引物,构建了用于沉默RBSDV S1、S2、S6、S8的amiRNA载体,之后转化玉米自交系综31,选择miRNA表达量高的纯合体株系进行自然发病试验,结果表明,转基因株系的抗病表现优于野生型玉米,几乎没有4级重度感染株,而野生型玉米全部发病,且4级重度感染株比例占37.5%,4个amiRNA载体中以靶向基因沉默抑制子的S6-miR159转基因玉米抗病表现最好,健株(0级)和轻微感病株(1级)比例达41.5%,说明利用amiRNA技术培育抗粗缩病玉米新品种也可行。
2.2利用非病毒来源的抗病毒基因
尽管研究者采用不同策略转化病毒基因序列均获得了抗甚至高抗病毒病的玉米材料,但将病毒基因序列转入植物,不确定因素较多,生物安全性一直以来备受人们质疑,因此,研究人员多年来致力于发掘不同来源的抗病毒基因。
2.2.1寄主抗病基因 植物在与病原体长期相互作用过程中形成了抗病基因(Resistance gene,R基因)系统,能够对病原体的侵染产生防御反应。至今在植物体内克隆获得具病毒抗性的R基因有40多种,多数基因的抗性具有特异性,人们利用植物R基因控制病毒已取得了很好的效果(牙库甫江·阿西木等,2015)。关于玉米中矮花叶病和粗缩病抗性的数量性状基因座(Quantitative trait locus,QTL)定位研究已有大量报道,但由于玉米对病毒抗性的遗传基础较复杂,从玉米中分离R基因的工作进展缓慢。Shi等(2013)研究表明,ZmeIF4E参与调控防卫基因表达并诱导寄主的局部和系统抗性,认为其可能是玉米粗缩病抗性候选基因。还有研究发现,玉米对SC-MV的抗性涉及Scmvl和Scmv2两个主效QTLs,其分别位于第6和第3染色体上,在抗病早期和后期发挥作用(Liu et al.,2017)。瞿会(2016)利用元分析方法整合玉米Scmv2信息,在“一致性”QTL区段内确定与抗病相关的候选基因,并克隆获得完整的基因编码区。Liu等(2017)针对Scmvl开展精细定位,克隆获得玉米的主效抗病基因ZmTrxh,其编码非典型的H型硫氧还蛋白,将ZmTrxh基因转入HiⅡ中发现,玉米早期对SCMV的抗性显著提高,外源ZmTrxh的表达水平高低与植株的抗性呈高度正相关。
2.2.2核糖体失活蛋白基因 核糖体失活蛋白(Ribosome-inactivating proteins,RIPs)属于N糖苷酶,能够特异性地将28S rRNA位于4324A处的腺嘌呤糖苷键降解,阻断EF2/GTP复合物与核糖体60S大亚基结合,使蛋白质合成受抑,该蛋白能够在病毒侵染细胞后使核糖体失活,导致病毒无法进行复制(牙库甫江·阿西木等,2015)。目前,玉米抗病毒基因工程研究中使用的RIPs主要是美洲商陆抗病毒蛋白(Pokeweed antiviral protein,PAP),其为从美洲商陆植株体内提取的一种碱性毒蛋白,属于Ⅰ型RIPs,具有对危害动物、植物及人体的病原真菌、细菌和病毒的广谱抗性。陈定虎等(2003)在转PAP基因玉米幼苗叶片上接种SCMV,至25 d植株没有发病,而对照植株在接种后7 d表现出花叶症状,表明PAP基因对SCMV侵染具有显著抑制作用。由于PAP对植物病原具有广谱抗性,其在植物保护领域具有广阔的应用前景。
2.2.3核酸酶基因 RNaseⅢ家族是一类依赖于dsRNA的特异性内切核酸酶,在真核生物和原核生物中均有存在,植物病毒基因组多为ssRNA,其复制时形成dsRNA中间体,正好是RNaseⅢ家族的靶向底物(Zhang et al.,2001)。为了降低野生型RNaseⅢ对植物正常生长产生的危害,可通过突变使其只能结合而不能切割dsRNA。Cao等(2013)将来源于大肠杆菌的RNaseⅢ基因突变体rnc70转化玉米综31,然后将得到的转基因植株种植于粗缩病重病田,经多代筛选和自交,获得对RBSDV具有高度抗性的2个转基因玉米新株系。田间鉴定发现,中等发病条件下,转化植株发病率和发病程度极低,出现免疫植株;严重发生年份,部分植株(其中一个株系中有6/7的植株)也显示出良好的抗性,而非转基因植株高度感病。人工接种条件下,转基因植株不仅发病率明显降低,病株中病毒的含量也明显下降。由于rnc70基因介导的抗性对能形成dsRNA结构的病毒均有效,因此,转rnc70基因的玉米材料可能对粗缩病、矮花叶病等多种玉米病毒病都表现出抗性,但需进一步验证,以充分发挥其应用潜力。
2.2.4其他抗病毒基因 dsRNA依赖性蛋白激酶R(PKR)具有多重功能,在寄主防御病毒侵染中发挥重要作用。PKR被细胞内的dsRNA或病毒复制中间体激活后,能使真核生物翻译起始因子elF-2的α亚单位磷酸化,导致病毒蛋白不能合成,从而抑制病毒繁殖(Sadler and Williams,2007)。PKR基因对RNA病毒具有广谱抗性,其在玉米抗病毒基因工程中的功效值得探索。尹祥佳(2011)应用In-Fusion克隆技术将人源PKR基因和抗草甘NEPSPS基因连接到载体上得到了双价植物表达载体,转化玉米自交系掖478和昌7-2后均获得阳性转化植株,为获得兼具病毒病抗性和除草剂抗性的转基因玉米材料打下了基础。
2-5A体系主要存在于高等脊椎动物体内,是一个由干扰素(1FN)诱导的降解单链RNA的抗病毒系统,之后的研究发现其对dsRNA动物病毒及植物病毒也表现出有效抗性,推测2-5A体系是具有应用前景的广谱抗病毒系统(Mitra et al.,1996)。张志燕(2004)将动物2-5A体系的RNase L基因和2-5A基因同时转入玉米自交系Z3和z31,均获得双阳性玉米再生植株,为培育具有广谱病毒病抗性的玉米品系打下了基础。
3展望
植物抗病毒基因工程研究所用的基因序列分为病毒来源的基因序列和非病毒来源的基因序列。由于病毒基因组小、序列易测定、基因易克隆和操作,利用病毒来源基因序列的抗病毒基因工程研究取得長足进展,采用的相关策略可分为病毒编码蛋白基因介导的抗病性、RNAi介导的抗病性和amiR-NA介导的抗病性3种。在玉米抗病毒基因工程研究中,利用前两种策略均得到了不同抗病水平的转基因植株,第3种策略的应用刚起步,与前2种策略相比,具有精确、高效、可控且操作简便的优点,在培育抗病毒植物中具有广阔的应用前景,因而有必要加强其在玉米抗病毒基因工程中的应用。利用病毒基因序列通常只抗1种或与其相关的几种病毒,目前提高矮花叶病抗性水平的研究相对较多,也获得了大量抗性种质材料,而抗粗缩病和兼抗2种病毒病的研究比较滞后,需要加强探索,以便尽快解决我国玉米生产上病毒病防治的难题。相对于病毒基因序列,将非病毒来源的抗性基因导入玉米生物安全性会更高,但从玉米中分离抗病基因的工作进展缓慢,还需要加快研究。此外,来自其他物种的核糖体失活蛋白、核酸酶、2-5A体系等的基因理论上均能促使受体植物获得广谱抗性,目前通过转化这些基因也得到了较多的玉米材料,但其广谱抗病毒效果有待进一步验证。
玉米抗病毒基因工程的研究主要集中在采用不同策略获得抗病毒植株及不同因素对抗病毒效果的影响方面,随着技术的发展和抗病性的提高,人们开始关注其安全性问题。目前已有研究表明,转基因抗矮花叶病玉米具有与常规玉米相同的生物学营养等价性,长期饲喂对大鼠的生长发育、生殖功能均无明显影响,其对大鼠也不存在亚慢性毒性和遗传毒性,但对大鼠的学习与记忆能力有明显降低作用,推测转基因玉米可能影响了神经细胞的正常活性(周文丽等,2014)。由于相关研究较少,关于转基因抗病毒玉米的食用安全性、生物安全性和生态安全性,还需进行全面评价。转基因玉米中关于抗虫、抗除草剂玉米的研究较成熟并已进入商品化生产阶段,而抗病毒玉米的研究与发展远远落后,只有经过抗病性和安全性的双重验证,转基因抗病毒玉米才能投入大田生产,真正发挥其抗病作用。
