长白山臭冷杉种群结构与动态
2017-05-21亢新刚杨英军王全军
滕 毅 ,张 青,亢新刚 ,杨英军 ,王全军
(1.北京林业大学,北京 100083;2.汪清林业局,吉林 延吉 133200)
种群动态是用来研究数量在时间和空间上的变化规律的一种手段[1-4]。生命表以及根据生命表绘制的存活曲线可以用来对种群动态变化进行研究或者作为种群统计的工具,根据生命表可以分析种群过去生长状态,而且还可以分析种群结构现存状态及更新策略以及预测将来的种群动态[5]。4个生存分析函数对表达种群的生存规律起到了辅助的作用[6]。不同龄级的株数组成形成了种群的天然更新动态,谱分析作为数学工具,可以用来分析复杂的种群天然更新数量的波动性和年龄更替的周期性[7]。通过对种群不同生长阶段的空间格局分析,了解其个体在水平空间结构上的分布变化规律[8]。
云冷杉天然针阔混交林作为世界重要森林类型之一,具有很高的经济、社会和生态效益[9]。长白山地区金沟岭林场的云冷杉针阔混交林不仅是我国东北地区主要林型,更是重要的风景林和用材林。臭冷杉是该林分类型的主要优势种和建群种。近些年来,已有不同地区冷杉种群结构动态的相关报道[10-13],与已有研究不同的是,本文通过对不同郁闭度条件下的臭冷杉种群进行调查,运用种群生命表法、谱分析法与不同发育阶段空间结构动态特征相结合,综合分析臭冷杉种群径级和空间分布结构和特征,呈现其不同生长阶段数量动态及变化规律,为合理经营臭冷杉种群提供理论依据。
1 研究区概况
研究地位于吉林省汪清县境内,金沟岭林场四面环山,有丰富的树木资源和动物资源,生态状况保护良好,地理位置坐标是(130°10′E、43°22′N)。林区属于低山丘陵地貌,海拔分布范围是300~1 200 m不等,坡度基本上都是在5°~25°之间,极少数也达到了35°比较陡的坡度。根据历年的土壤情况调查,发现本实验区的土壤区位于中低山灰化土壤区,属于玄武岩河谷地区大多为冲击土、沼泽土或草甸土等,土壤呈粘性、颗粒状,有湿松、根系多等特点,土壤大多40 cm左右厚。本实验区是季风型气候,一月份温度最低,平均气温零下32 ℃,最高气温出现在7月,平均气温22 ℃,全年平均温度3.9 ℃,积温2 144℃,霜期从9月中旬到翌年5月末,积雪平均50 cm厚,全年生长期有120 d。实验区区主要森林类型以针阔混交林为主,植被以云冷杉为主。乔木主要以臭冷杉Abies nephrolepis、红皮云杉Picea koraiensis、长白鱼鳞云杉Picea jezoensis为主,其他的伴生树种有红松Pinus koraiensis、色木槭Acer mono、白桦Betula platyphylla、青楷槭Acer tegmentosum、山杨Populus davidiana、花楷槭Acer ukurunduense、水曲柳Fraxinus mandschurica、榆树Ulmus pumila、黄菠萝Phellodendron amurense、胡桃楸Juglans manshurica等。主要灌木包括柳叶绣线菊Spiraea salicifolia、胡枝子Lespedeza bicolor、忍冬Lonicera japonica和刺五加Acanthopanax senticosus等。主要草本植物有唐松草Thalictrum aquilegifolium、小叶芹Aegopodium alpestre、酢浆草Oxalis acetosella、舞鹤草Maianthemum bifolium等。
2 研究方法
2.1 样地设置
通过在金沟岭林场检查法1大区的2、4、5这3个小区中,通过机械布设,共设有70个10 m×10 m的更新样地,经过2014年复测,已有三次测量数据。林木平均年龄为85 a,平均胸径为22.1 cm,林分平均密度为874株/hm2。将不同郁闭度样地分级,郁闭度0.5~0.6为较低郁闭度等级,Ⅰ级;0.7~0.8为中等郁闭度等级,Ⅱ级;0.9~1为较高郁闭度等级,Ⅲ级。每个等级随机选取10个10 m×10 m样地,每个等级样地面积为1 000 m2。在每个样地内进行每木检尺(DBH≥5 cm),记录树种名称、树高、胸径;记录臭冷杉(DBH<5 cm且H≥30 cm)的株数,并记录样地植被状况和生境条件。
2.2 年龄结构分析
根据样地内臭冷杉的最大胸径计算出所对应的年龄,对臭冷杉种群进行径级分组,用以代替龄级。臭冷杉种群径级分组:第1径级为H≥0.3 m,DBH<1 cm,第2径级为1 cm≤DBH<5 cm,其他情况按照每隔5 cm为一个径级进行划分。把树木径级从小到大的顺序看作是时间顺序关系,第1径级对应第1龄级,第2径级对应第2龄级,以此类推一一对应,统计各龄级株数。[14]。
为了分析臭冷杉种群不同生长阶段的空间分布格局,根据其生长特性,将臭冷杉按照胸径树高分成4个生长阶段,H≥30 cm,DBH<1 cm为下层幼苗阶段,1 cm≤DBH<5 cm为林分下层幼树阶段,5 cm≤DBH<25 cm为林分中层中树阶段,DBH≥25 cm为林分上层大树阶段[15]。
2.3 生命表
静态生命表又被称作特定时间生命表,常被应用在长寿命的木本植物种群的统计分析中。静态生命表是一种可以有效统计种群死亡过程的一种方法,不仅可以直观地体现出臭冷杉种群内不同龄级的存活个体数、种群各龄级间隔的树木死亡率和种群区间寿命等指数,同时还反映了种群在特定时期内的动态变化规律[16]。根据3种不同郁闭度臭冷杉种群不同龄级的个体数,编制静态生命表。在特定时间生命表中,可以通过实测值求得以下关系式:

在生命表中有可能高龄级的树比低龄级的树多的现象发生,由于死亡率对于年龄来说应该是连续变化的,因而应该具有某种光滑性,所以采取匀滑技术对臭冷杉种群数据进行修匀,修匀可以针对一个参数或者是一个参数系列。
为了更好的阐明臭冷杉种群的生存规律,揭示其种群动态,本文引入用来进行生存分析的4个函数,生存率函数、累积死亡率函数、死亡密度函数和危险率函数[17]。式中:Si是存活率,qi是死亡率,hi是龄级长度。
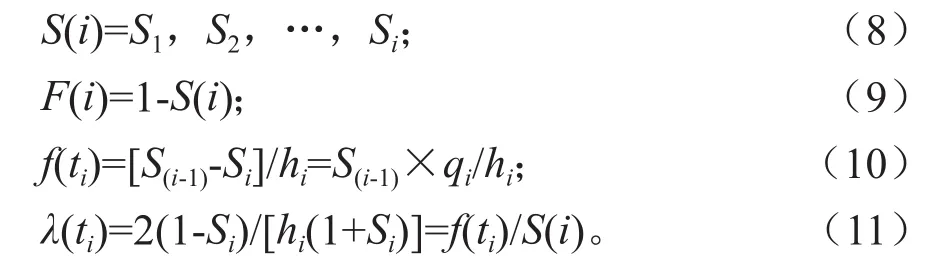
2.4 谱分析法
本文运用的谱分析方法是用来揭示臭冷杉种群数量的周期性波动,探讨目标林分分布波动性和年龄更替过程的周期性的数学工具。臭冷杉种群的天然更新过程可以通过不同龄级株数分布波动表现出来。谱分析法的正弦波形式是:
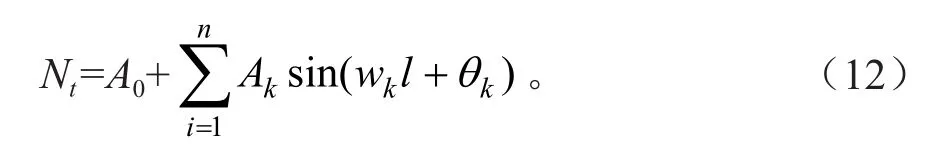
将臭冷杉种群各年龄阶段个体分布视为一个时间系列t,用Xt表示t年龄序列时臭冷杉个体数,n为系列总长度;p=n/2为谐波的总个体数;T为正弦波的基本周期,即时间系列t的最长周期,Fourier分解中的各个参数可以用下式来估计:

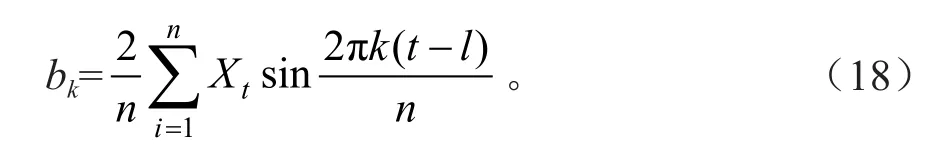
式中,ak、bk为参数估计值。通过对臭冷杉各龄级个体数量波动较大的实际情况判断后决定,在计算时进行对数化处理,用Xt′=ln(Xt+1)代替Xt,然后利用谱分析中的公式分别计算各种情况下各个波形的振幅。A1为基波,A2,…,An为谐波。
2.5 种群分布格局
为了分析臭冷杉种群不同生长阶段的空间分布格局,根据其生长特性,将臭冷杉按照胸径树高分成4个生长阶段。对于不同发育本文通过应用方差/均值比率、平均拥挤度、聚块性指标、负二项指数(K)、丛生指标(I)、Cassie指标(CA)这些方法来判断臭冷杉种群的空间分布格局[18-19]。
3 结果与分析
3.1 种群生命表
3种郁闭度条件下,幼苗幼树数量在整个种群中占比都是最高的,分别为73.28%、73.39%和83.83%。较高生命期望均出现在高死亡率后一个龄级,表示臭冷杉通过强烈的环境筛选后,可以表现出较高的生命期望,此阶段的臭冷杉生存质量最高。
死亡率曲线反映了不同郁闭度条件下死亡率的动态变化,从图1中可以看出,臭冷杉种群的死亡率和亏损度变化基本一致,不同郁闭度条件下死亡率曲线存在明显的差异:在较低郁闭度等级时,死亡率有2个高峰,分别是第1、8龄级;中等郁闭度等级时,死亡率有个高峰,分别是第1、4、9龄级;较高郁闭度等级时,死亡率有3个高峰,分别是第1、2、10龄级。从图1(a)中可以看出,臭冷杉种群在较低郁闭度等级时,1龄级到2龄级死亡率最高,说明幼苗生存压力较大,因为样地郁闭度低,导致灌木疯长,对幼苗的压迫比较严重,中龄级存活个体数量较为稳定,经过生长竞争,高龄级臭冷杉进入衰老期存活个体数较少;从图1(b)中可以看出,第1龄级存活数量远多于其它龄级,说明林下更新幼苗状态较好,但在幼苗阶段死亡率较高,第2~4龄级呈缓慢下降趋势,第4龄级死亡率显著提高,在这个阶段,臭冷杉的生长对环境资源的需求增大,种内竞争加剧,出现死亡高峰;从图1(c)中可以看出,在整个龄级序列中,第2龄级的死亡率最高,说明在此郁闭度下,由于上层林木郁闭度较大,臭冷杉个体在从更新层生长到进阶的过程中生存压力大,臭冷杉个体很难进阶。以上说明虽然都是臭冷杉种群,但是在不同的郁闭度条件下,臭冷杉种群的生命过程是不同的,所以在对臭冷杉种群进行经营管理的时候要具体情况具体分析。
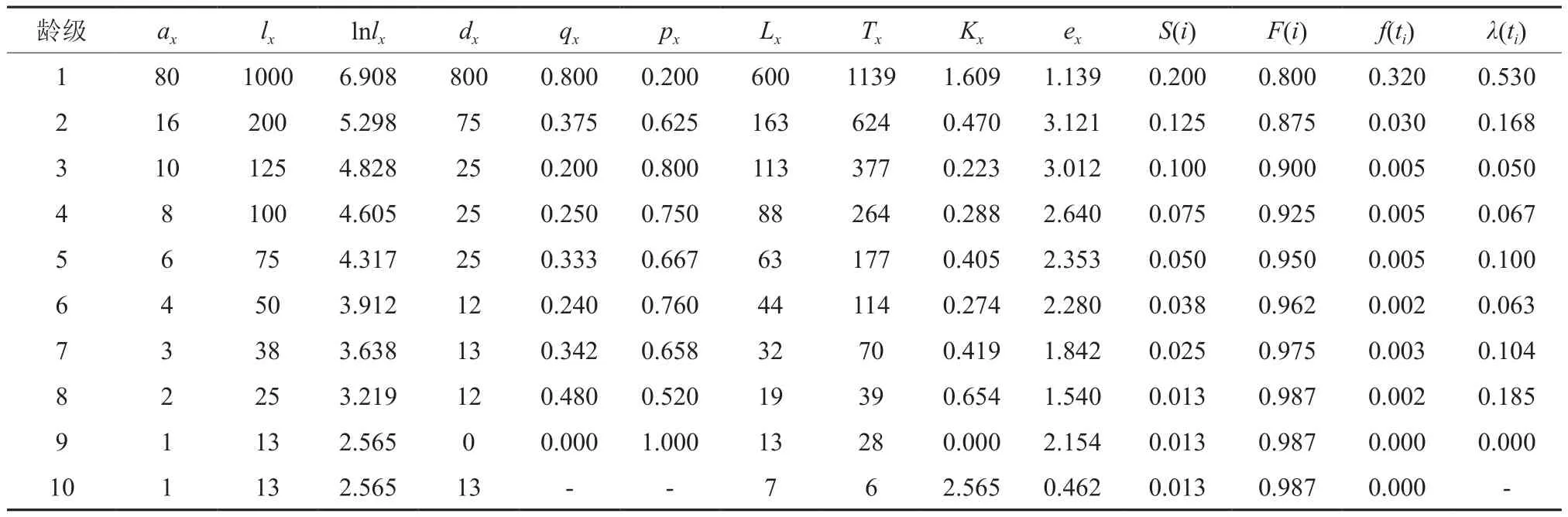
表1 较低郁闭度等级臭冷杉种群静态生命表Table 1 Crown density level to Ⅰ Abies nephrolepis population static life
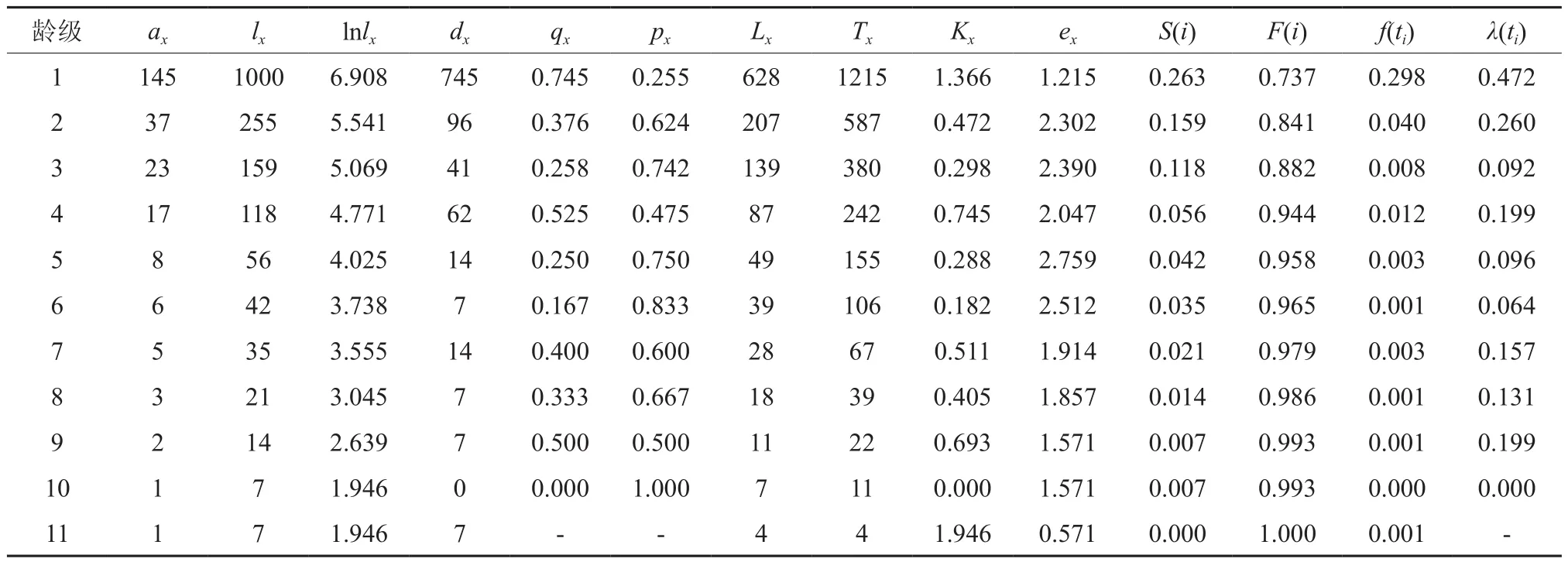
表2 郁闭度等级为Ⅱ臭冷杉种群静态生命表Table 2 Crown density level to Ⅱ Abies nephrolepis population static life
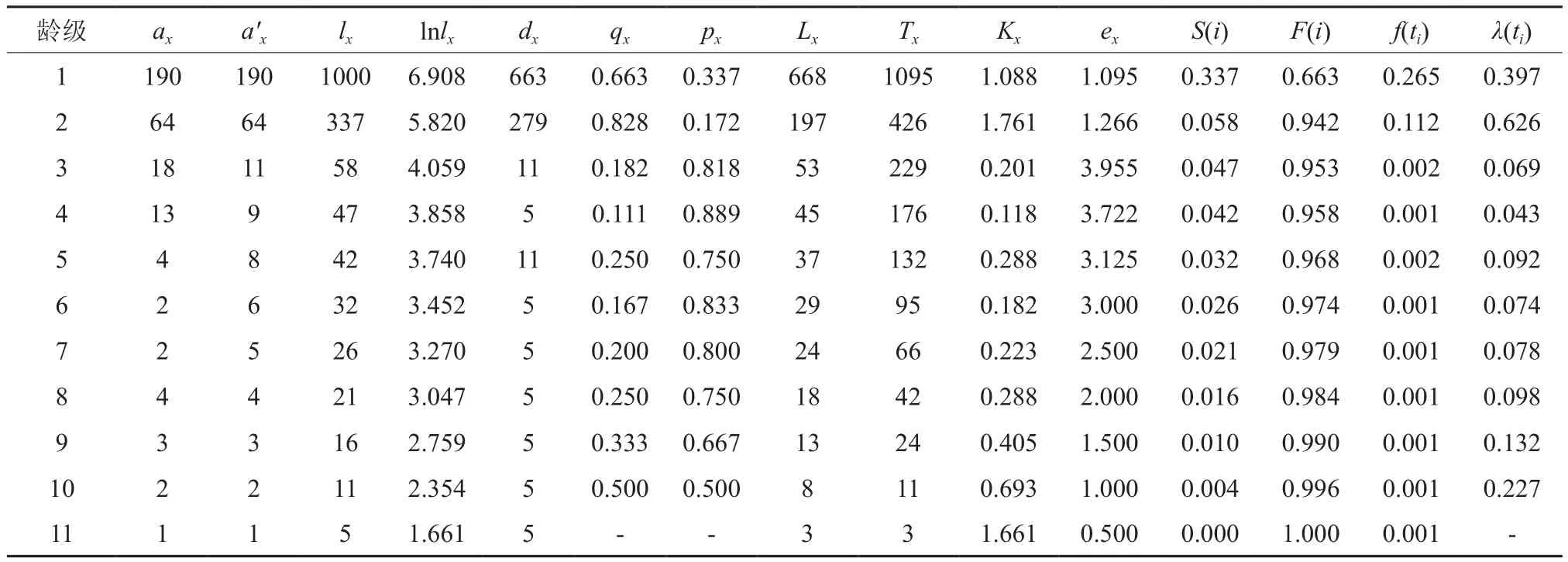
表3 郁闭度等级为Ⅲ臭冷杉种群静态生命表Table 3 Crown density level to Ⅲ Abies nephrolepis population static life
3.2 存活曲线
存活曲线是用各龄级存活个体数量相对时间来描述特定年龄的死亡率,表达种群动态的一种手段。以与径级相对应的龄级为横坐标,以存活株数的自然对数为纵坐标,绘制三种不同郁闭度条件下的臭冷杉种群存活曲线。这是生物物种经过长时间的自然选择而形成的。Deevey把存活曲线定义个3种不同类型,Ⅰ型存活曲线是形状为凸型的曲线,表示早期死亡率低,大多数种群个体能够活到平均的生理年龄,但是在达到平均年龄时,较短时间内死亡率急剧升高,几乎全部死亡;Ⅱ型存活曲线是形状为对角线型的曲线,表示各个龄级都具有几近相同的死亡率;Ⅲ型存活曲线是形状为凹型的曲线,表示种群内个体早期死亡率高,此后的死亡率较低且趋于稳定。臭冷杉种群的存活曲线是用标准化存活数的对数表达的,以龄级为横坐标,以标准化存货量的对数为纵坐标,不同郁闭度条件下存活曲线如图2所示。

图1 不同郁闭度条件下臭冷杉种群死亡率和亏损度曲线Fig.1 Mortality rate and vanishing curve of Abies nephrolepis populations under different canopy densities
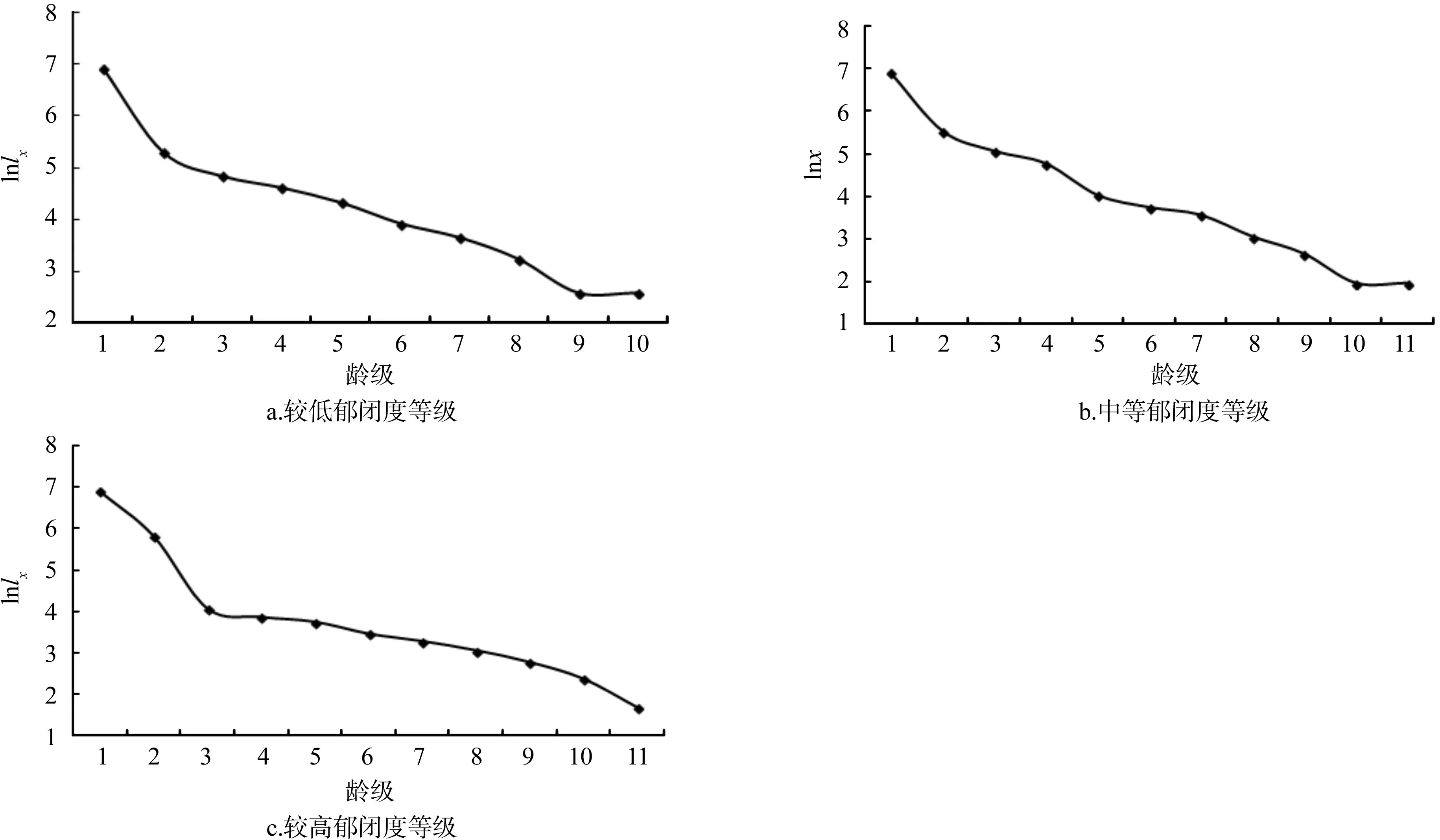
图2 臭冷杉种群存活曲线Fig.2 Survival curve of Abies nephrolepis populations
总体上来说,3种郁闭度条件下的臭冷杉种群的存活曲线介于Deevey-Ⅱ型和Deevey-Ⅲ型之间。由于不能确定是哪种模型,本文选用两种数学模型来检验,指数方程来描述Deevey-Ⅱ型曲线,用幂函数来描述Deevey-Ⅲ型曲线[17],建立相应模型得到:
郁闭度为Ⅰ级时:
Nx=7.016e-0.102x(F=310.319,R2=0.964,Sig <0.001)
Nx=7.41x-0.408(F=62.941,R2=0.887,Sig <0.001)
郁闭度为Ⅱ级时:
Nx=7.611e-0.123x(F=340.032,R2=0.987,Sig <0.001)
Nx=8.247x-0.513(F=55.168,R2=0.860,Sig <0.001)
郁闭度为Ⅲ级时:
Nx=6.863e-0.114x(F=101.459,R2=0.919,Sig <0.001)
Nx=7.633x-0.496(F=66.491,R2=0.881,Sig <0.001)
根据检验结果可以判断臭冷杉种群在3种郁闭度条件下,存活曲线均更趋于Deevey-Ⅱ型,都属于稳定型种群。
3.3 生存分析
臭冷杉种群的生存率函数曲线随着龄级的增加而下降,累积死亡率函数曲线与之相反。在较低郁闭度等级时,臭冷杉种群在第1龄级时,危险率最高,死亡密度也最大,说明臭冷杉幼苗的生长过程中竞争十分激烈,原因是因为林分郁闭度低,导致灌木草本疯长,营养空间竞争激烈;第8龄级也出现了高危险率,但是死亡密度很小,是由于臭冷杉种群在高龄级时出现的生理衰老导致的。在中等郁闭度等级时,臭冷杉种群在第1、4、9龄级出现了高危险率,第1龄级个体数量最多种内竞争激烈,死亡密度最大,危险率也是最高的;在第4龄级出现了较高危险率,但是死亡密度并不是太大,出现的原因是因为臭冷杉生长到中龄级阶段,对空间和自然条件需求量加大,种内竞争加剧,从而出现了较高危险率;第9龄级虽然危险率高但是死亡密度并不大,是由于生理衰老。在较高郁闭度等级时,臭冷杉种群在第1、2、10龄级表现出较高危险率,第1、2龄级死亡密度很大,第9龄级较小,与其它两个郁闭度等级不同的是,臭冷杉种群在第二龄级也表现出很高的危险率,因为林分郁闭度较大,上层生长空间封闭,导致幼树到进阶的阶段出现大量死亡。整体上来说,臭冷杉种群幼苗阶段个体数量大幅度减少,中期表现较稳定,后期出现衰退现象。
3.4 波谱分析
臭冷杉种群数量动态是通过不同龄级个体数量来体现的。本文以5 cm为级差进行波普分析,根据3种郁闭度条件下,不同龄级个体数量进行谱分析,结果如表4所示。
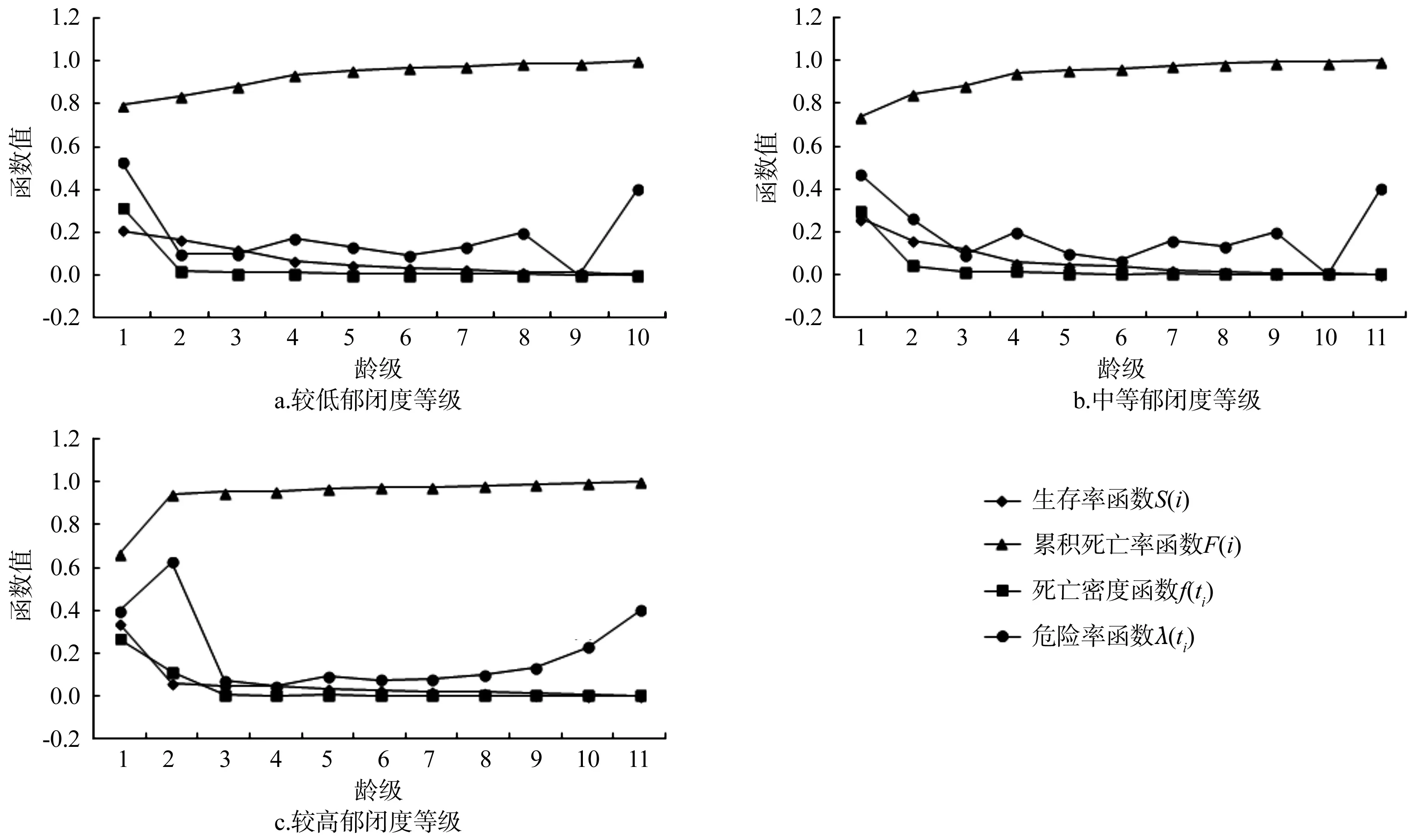
图3 不同郁闭度条件下臭冷杉种群的4种函数曲线Fig.3 Four survival function curves of Abies nephrolepis populations under different canopy densities
3种郁闭度条件下臭冷杉种群的整个生长过程受基波的影响是非常显著,3种情况下基波A1值都是最大的,证明臭冷杉种群整个生命周期生物学特性控制着其生长过程,而且趋向性非常明显,且表现出来的波动不是单一的周期,与天然更新的周期波动相类似,整个生长动态表现出大周期内有小周期的现象。但可能由于所调查的臭冷杉种群年龄不够大,最大径级为45~50 cm,林分还没有达到成熟阶段,时间序列长度不足以表达出基本周期,也没表现出明显的固有周期长度,但其存在性是肯定的。
从表2中还可以发现,臭冷杉种群数量动态不仅受基波影响,而且还存在小周期波动。较低郁闭度等级和中等郁闭度等级时,有A4和A 52个小周期,A4的波动对应径级30~35 cm处,对应第8龄级,臭冷杉种群在此过程发生强烈的竞争后,存活的个体达到主林层高度,产生林木分化、数量调节现象,这有利于种群的发展,A5的波动对应径级35~40 cm,在此阶段,臭冷杉种群逐渐进入生理衰退期,种群数量发生了波动。较高郁闭度等级时,只有A 5发生波动,说明臭冷杉种群在高郁闭度条件下,数量调节能力较差,种群更新状态一般。
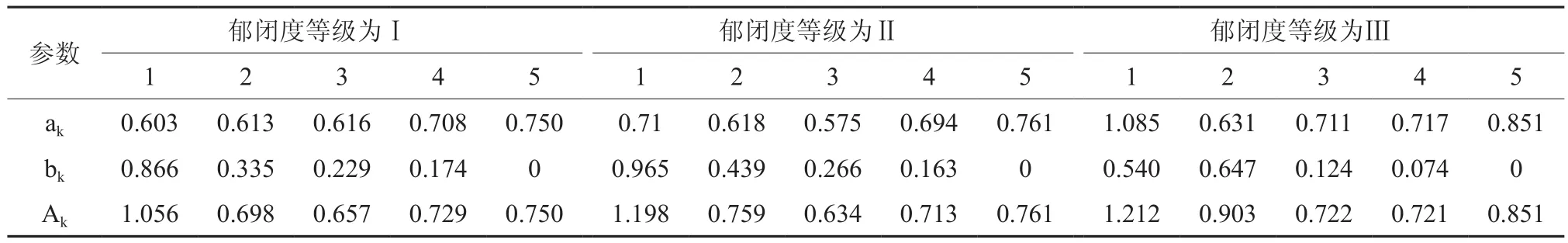
表4 不同郁闭度臭冷杉种群的周期性波动Table 4 Periodic fluctuation of Abies nephrolepis population under different canopy densities
3.5 空间分布格局
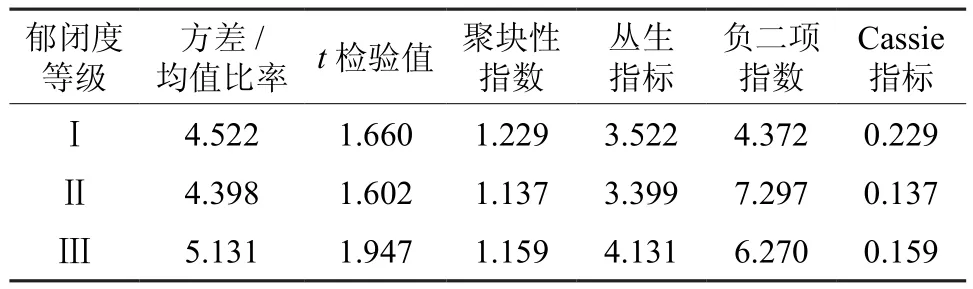
表5 臭冷杉种群整体分布格局Table 5 Distribution pattern of Abies nephrolepis population
由表5可知,不同郁闭度等级的臭冷杉种群分布格局有一定相似性。t检验表明:3种郁闭度条件下臭冷杉种群方差均值比都大于1,说明3种郁闭度条件下臭冷杉种群均呈集群分布;聚块性指标大于1也说明臭冷杉种群趋于集群分布;但是3种郁闭度条件下聚集程度各不相同,方差均值比率法和丛生指标分析结果一致,聚集强度随郁闭度的减小而减小,聚块性指标、负二项指数和Cassie指标反映出的结果一致,聚集强度由强到弱分别是,郁闭度Ⅰ、郁闭度Ⅲ和郁闭度Ⅱ。
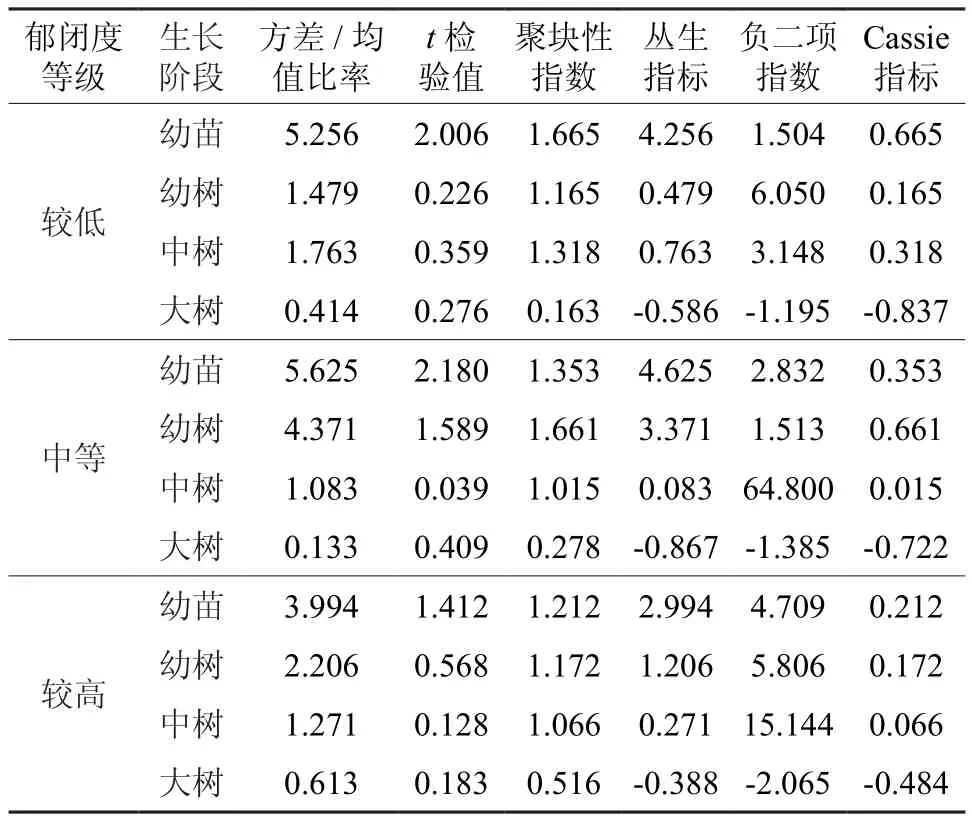
表6 不同郁闭度臭冷杉种群不同发育阶段分布格局Table 6 Different densities of Abies nephrolepis population distribution pattern in different developmental stages
较低郁闭度等级时,臭冷杉在幼苗、幼树和中树阶段都呈集群分布,幼树和中树的聚集程度很弱,大树阶段呈均匀分布;中等郁闭度等级时,幼苗和幼树阶段呈集群分布,幼树阶段聚集程度较高,中树阶段趋近于随机分布,大树阶段为均匀分布;较高郁闭度等级时,幼苗和幼树阶段为集群分布,幼苗阶段聚集程度较高,中树向随机分布过度,大树阶段呈均匀分布。整体上来看,臭冷杉在幼苗幼树阶段呈集群分布,中树阶段向随机分布发展最终在大树阶段形成均匀分布。
4 结论与讨论
对臭冷杉种群生命表的分析表明,3种郁闭度条件下,幼苗幼树数量在整个种群中占比都是最高的,分别为73.28%、73.39%和83.83%,更新苗数量充足。臭冷杉种群的死亡率和亏损度变化基本一致,不同郁闭度条件下死亡率曲线存在明显的差异:较低郁闭度等级时,死亡率有2个高峰,分别是第1、8龄级;中等郁闭度等级时,死亡率有个高峰,分别是第1、4、9龄级;较高郁闭度等级时,死亡率有3个高峰,分别是第1、2、10龄级。臭冷杉种群最高生命期望值出现在较高死亡率之后,说明在经过强烈的环境筛选之后,存活下来的臭冷杉在当时有很强的生命力。
经过指数方程和幂函数的检验,发现臭冷杉种群在3种郁闭度条件下,存活曲线均更趋于Deevey-Ⅱ型,都属于稳定型种群。
本文运用4个生存函数,通过对3种郁闭度条件下臭冷杉种群进行分析,很好的说明了种群的结构和动态,整体上来说,臭冷杉种群幼苗阶段个体数量大幅度减少,中期表现较稳定,后期出现衰退现象。与死亡率曲线变化基本一致。
谱分析结果显示,3种郁闭度条件下臭冷杉种群存在着明显的周期性,反映了臭冷杉种群动态的周期波动特征。出现波动的径级与死亡率曲线和生存分析曲线结果相呼应。
臭冷杉种群在不同郁闭度条件下整体的分布格局均呈集群分布,聚集强度各不相同,集群强度由强到弱分别是,郁闭度Ⅰ、郁闭度Ⅲ和郁闭度Ⅱ。臭冷杉种群在不同发育阶段上水平分布格局有差异由集群分布向随机分布转变最终达到均匀分布。在幼苗、幼树阶段表现为集群分布,在中树阶段向随机分布转变,最终在大树阶段达到均匀分布,这个结果与年龄结构及数量动态结论相匹配。
参考文献:
[1]岳春雷,江洪,朱荫湄.濒危植物南川升麻种群数量动态的分析[J].生态学报,2002,22(5):793-796.
[2]郭 微,上官铁梁,工志明,等.灵空山油松种群年龄结构与动态分析[J].植物科学学报,2013,31(2):130-135.
[3]刘贵峰,臧润国,丁 易,等.天山云杉种群数量动态研究[J].林业科学研究,2011,24 (5):659-662.
[4]刘国军,张希明,朱军涛,等.准噶尔盆地东南缘梭梭种群结构与动态研究[J].西北植物学报,2011,31(6):1250-1256.
[5]洪 伟,王新功,吴承祯,等.濒危植物南方红豆杉种群生命表及谱分析[J].应用生态学报,2004,15(6):1109-1112.
[6]刘海洋,金晓玲,沈守云,等.湖南珍稀濒危植物珙桐种群数量动态[J].生态学报,2012,32(24):7738-7746.
[7]陈远征,马祥庆,冯丽贞,等.濒危植物沉水樟的种群生命表和谱分析[J].生态学报,2004,26(12):4267-4272.
[8]刘普幸,鹿晨昱,姚晓军,等.敦煌绿洲不同生境胡杨种群结构与空间格局研究[J].北京林业大学学报,2011,33(2):48-52.
[9]李 冰,樊金拴,车小强.我国天然云冷杉针阔混交林结构特征、更新特点及经营管理 [J].世界林业研究,2012,25(3):43-48.
[10]党海山,张燕君,张克荣,等.秦岭巴山冷杉(Abies fargesii)种群结构与动态[J].生态学杂志,2009,28(8):1456-1461.
[11]李先琨,向悟生,唐润琴.濒危植物元宝山冷杉种群生命表分析[J].热带亚热带植物学报,2002,10(1):9-14.
[12]卢 杰,潘 刚,罗大庆,等.濒危植物急尖长苞冷杉种群生命表分析[J].水土保持研究,2010,17(5):212-216.
[13]史小华,刘 毅,彭佳龙,等.秦岭冷杉和巴山冷杉种群年龄结构及动态的比较分析[J].东北林业大学学报,2009,37(1):10-14.
[14]李艳丽,杨 华,亢新刚,等.长白山云冷杉种群结构和动态分析[J].北京林业大学学报,2014,36(3):18-25.
[15]张金屯,植物种群空间分布的点格局分析[J].植物生态学报,1998,22(4):344-349.
[16]黎颖芳,周江雄,刘建华.生命表的构造理论[M].天津:南开大学出版社,2001.
[17]李建贵,潘存德,彭世揆,等.天山云杉种群统计与生存分析[J].北京林业大学学报,2001,23(1):84-86.
[18]李 俊,郑小贤,刘东兰,等.金沟岭林场落叶松人工林的空间结构特征[J].中南林业科技大学学报,2014,43(1):60-63.
[19]于大炮,周 莉,董百丽,等.长白山北坡岳桦种群结构及动态分析[J].生态学杂志,2004,23(5):30-34.
