侧柏母树大小对种子特性和幼苗生长的影响
2019-07-13张金颢曹帮华毛培利山东英才学院山东济南5004
张金颢 曹帮华 毛培利(.山东英才学院 山东 济南 5004;
2.山东农业大学林学院/黄河下游森林培育国家林业局重点实验室/山东泰山森林生态系统国家定位观测研究站, 山东 泰安 271018;3. 山东省林木种苗和花卉站, 山东 济南 250014)
侧柏(Platycladusorientalis)是我国重要乡土树种,具有适应性强、抗严寒、耐干旱瘠薄等特性,是我国荒山造林和水土保持的先锋树种。随着侧柏人工林的不断发育,其天然更新逐渐受到了人们的关注,尤其是对于二十世纪五六十年代营造的林分。这是因为成功的天然更新对侧柏人工林的稳定和生态功能持续发挥具有重要的意义。通过对侧柏人工林林下幼苗分布的调查,认为郁闭度是影响侧柏林下幼树生存、生长的首要因子[1]。王斐和臧丽鹏认为,除了充足的光照,较高的土壤含水量也是侧柏幼苗更新的重要限制因子[2]。除了更新的环境因子,种子质量也是侧柏人工林天然更新的关键因子。张梦颖等发现,随海拔增加,侧柏人工林种子活力先增加再降低,在海拔300~500 m最高,种子发芽率超过60%[3]。但是关于母树大小对侧柏种子质量及幼苗生长的影响目前还没有系统研究。
林木种子的质量直接影响着人工林的生长发育[4]和森林天然更新能否成功[5]。树木个体大小对种子质量有着重要的影响。随着母树胸径的增加,黑松种子的发芽率逐渐增加[6]。太白红杉种子的千粒重随母树基径的增加逐渐增加,但南坡是先增加再降低[7]。对于水青树[8]、胖大海[9]、四合木[10]、Pinusechinata[11]和Sorbustorminalis[12]则是随着母树胸径或基径的增加,种子质量先增加再降低,在中间胸径达到最大值。也有研究表明,香果树[13]、Piceamariana[14]的种子发芽率不受母树胸径的影响。以上研究结果表明,随母树个体大小的增加,林木种子质量有逐渐增加、先增加再降低和变化不显著3种情况。对不同胸径黑松母树种子特性的研究发现,胸径大的母树不仅种子质量好,而且形成的幼苗有着高的生物量和生长速率,这对早期幼苗建立有着重要意义[6]。
侧柏林是泰山主要人工林之一,也是主要的风景林之一。大部分是上二十世纪五十年代营造,以其为主的林分总面积约为1 080 hm2,主要分布在海拔800 m以下的山坡上。由于是风景林,该林分的改造要求严格,极少进行抚育更新,造成林分结构不合理,生态稳定性和功能的发挥受到一定的影响。天然更新对林分结构改造效果好、成本低。本文以泰山20世纪50年代营造的侧柏纯林中不同胸径母树的种子为对象,研究种子萌发和幼苗生长动态,阐明母树大小对幼苗建立阶段的影响机制,为侧柏人工林的天然更新管理提供理论依据。
1 材料与方法
1.1 研究区域概况
泰山位于山东省的中部,横亘于济南、泰安两市之间。研究区属于暖温带大陆性季风气候区,多年平均气温为12.6 ℃,无霜期196 d,≥10 ℃的活动积温3 821 ℃,多年平均降水量758 mm,降水集中在6—9月。土壤类型主要是棕壤土,pH值为6.0左右,土层厚度15~60 cm。泰山植被大部分为人工林,森林总面积9 490 hm2左右,森林覆盖率81.5%。主要的树种有侧柏(P.orientalis)、麻栎(Quercusacutissima)、刺槐(Robiniapseudoacacia)、黑松(Pinusthunbergii)、赤松(Pinusdensiflora)、油松(Pinustabuliformis)等。
1.2 研究方法
1.2.1种子采集
2017年10月,根据调查结果[3],在海拔为200 m的泰山普照寺周边的侧柏林内采集了3个胸径径级的侧柏母树种子,每个径级采集母树5~10株。径级Ⅰ的胸径为9.2~11.8 cm,径级Ⅱ为14.2~16 cm,径级Ⅲ为18.8~21.2 cm。通过自然风干待球果开裂后收集饱满种子用于实验。每个径级种子用百粒法测定千粒重,并分别选择30粒种子用游标卡尺测定最大长度、最大宽度。
1.2.2种子萌发试验
种子萌发试验在山东农业大学林学试验站内进行。2018年4月24日,将取自侧柏林内的土壤过筛、去除杂质,放入直径20 cm、高20 cm的白色塑料盆内至预先标定的深度。浇水至盆底有水流出后,每个花盆摆20粒侧柏种子于表面,再覆盖0.5 cm土于表面盖住种子。按照采种的3个母树径级水平(Ⅰ、Ⅱ、Ⅲ),每个水平9个重复,共计27盆,在室外进行萌发实验。种子突破表土后视作萌发。种子萌发后,每天记录萌发的种子数。试验期间,定时浇水,使土壤保持湿润。6月13日,萌发试验结束观察。统计发芽率(Germination rate,Gr)、发芽指数(Germination index,Gi)和平均发芽时间(Mean germination time,MGT)。

图1 不同径级侧柏母树种子大小特征

图2 母树径级对侧柏种子萌发的影响
Gr(%)=G1/N×100%
式中:G1为萌发数,N为供试种子总数。
MGT=∑(D×n) /∑n
式中:n为对应各日正常发芽的粒数,D为种子置床起的天数。
Gi=∑(Gt/Dt)
式中:Gt为各日净发芽数;Dt为相应发芽日。
1.2.3幼苗生长试验
种子萌发试验结束后,每盆选取大小一致的幼苗3株,分别于7月12日、8月23日、9月23日和10月26日测定幼苗的高度和地径。试验期间,定期浇水。2018年10月26日,幼苗生长试验结束,取出幼苗,并用清水冲洗干净,分为地上部和根系。用HP Scanjet 8200扫描仪扫描根系,再用Delta-T Area Meter Type AMB 2根系参数分析软件分析扫描的图片获取根系长度和根系表面积。然后将地上部和根系在120 ℃的烘箱中杀青30 min,再在80 ℃下烘干直至恒重,称量干重并记录。计算幼苗干重(Biomass,地上部重+根重)、根冠比(Root/shoot ratio,根重/生物量)、比根长(Specific root length,SRL,根长度/根系干重)和比根表面积(Specific root area,SRA,根表面积/根系干重)。计算相对苗高生长速率(Relative height growth rate, RGRH)和相对地径生长速率(Relative ground diameter growth rate, RGRD)。
RGRH= (lnH2-lnH1)/(T2-T1)
RGRD= (lnD2-lnD1)/(T2-T1)
式中:H为幼苗高度,D为幼苗地径,T1代表试验开始时间,T2代表试验结束时间。
1.3 统计分析
所有数据采用SPSS 17.0软件(SPSS,Chicago,IL,USA)进行分析。采用单因子方差分析法(One-Way ANOVA)检验了不同母树径级之间在千粒重、种子最大长、种子最大宽、发芽率、发芽指数、平均发芽时间、幼苗干重、根冠比、比根长和比根表面积。用双因子方差分析法(Two-Way ANOVA)比较了幼苗苗高、地径生长指标。若差异显著,再用最小显著差异法(LSD)来进行多重比较。显著性差异检验水平是p<0.05。
2 结果与分析
2.1 不同径级母树种子大小
随母树径级的增加,千粒重、种子最大长和种子最大宽逐渐增加(图1)。对于千粒重,径级Ⅰ的种子变化幅度为21.31%,径级Ⅱ的为30.49%,径级Ⅲ的为49.49%。而方差分析结果表明,千粒重(F=0.30,p=0.75)、种子最大长(F=0.76,p=0.50)和种子最大宽(F=0.36,p=0.71)在不同径级之间差异均不显著。

图3 母树径级对侧柏幼苗生长的影响

图4 母树径级对侧柏幼苗相对生长速率的影响
2.2 不同径级母树种子的萌发
随母树径级的增加,发芽率、发芽指数逐渐增加,平均发芽时间逐渐降低(图2)。方差分析结果表明,发芽率(F=9.49,p<0.01)和发芽指数(F=11.76,p<0.01)在不同径级之间差异极显著,平均发芽时间(F=3.58,p<0.05)在不同径级之间差异显著。发芽率和发芽指数都是在径级Ⅰ和径级Ⅱ之间差异不显著(p>0.05),极显著小于径级Ⅲ(p<0.01);对于平均发芽时间,则是在径级Ⅰ和径级Ⅱ之间差异不显著(p>0.05),显著大于径级Ⅲ(p<0.05)。
2.3 不同径级母树幼苗的生长动态
2.3.1苗高地径生长动态
随生长月份的增加,不同径级侧柏的幼苗苗高和地径逐渐增加(图3)。方差分析结果表明,母树径级大小对苗高影响不显著(F=2.50,p=0.10),对地径影响显著(F=4.10,p<0.05);生长月份对苗高(F=128.42,p<0.01)和地径(F=150.07,p<0.01)都影响极显著;但交互作用在苗高(F=1.24,p=0.31)和地径(F=2.03,p=0.0.09)上都影响不显著。不同径级之间,径级Ⅰ的地径极显著低于径级Ⅲ(p<0.01),径级Ⅰ与径级Ⅱ、径级Ⅱ与径级Ⅲ之间均差异不显著(p>0.05)。在生长月份之间,对于苗高是7月<8月<9月和10月(p<0.01),而9月和10月之间差异不显著(p>0.05);对于地径则是7月<8月<9月<10月(p<0.01)。
2.3.2相对苗高和地径生长速率动态
随生长月份的增加,径级Ⅰ和径级Ⅲ的相对苗高生长速率与相对地径生长速率都是先增加再降低,而径级Ⅱ的则是逐渐降低(图4)。方差分析结果表明,母树径级大小对相对苗高生长速率影响显著(F=3.90,p<0.05),对相对地径生长速率影响不显著(F=0.31,p=0.74);生长月份对相对高度生长速率(F=82.22,p<0.01)和相对地径生长速率(F=9.28,p<0.01)都影响极显著;二者交互作用对相对苗高生长速率影响极显著(F=6.54,p<0.01),对相对地径生长速率影响不显著(F=2.45,p=0.70)。不同径级之间,径级Ⅰ的相对苗高生长速率显著低于径级Ⅲ(p<0.05),径级Ⅰ与径级Ⅱ、径级Ⅱ与径级Ⅲ之间均差异不显著(p>0.05)。生长月份之间,相对苗高生长速率和相对地径生长速率都是7—8月与8—9月之间差异不显著(p>0.05),二者极显著高于9—10月(p<0.01)。
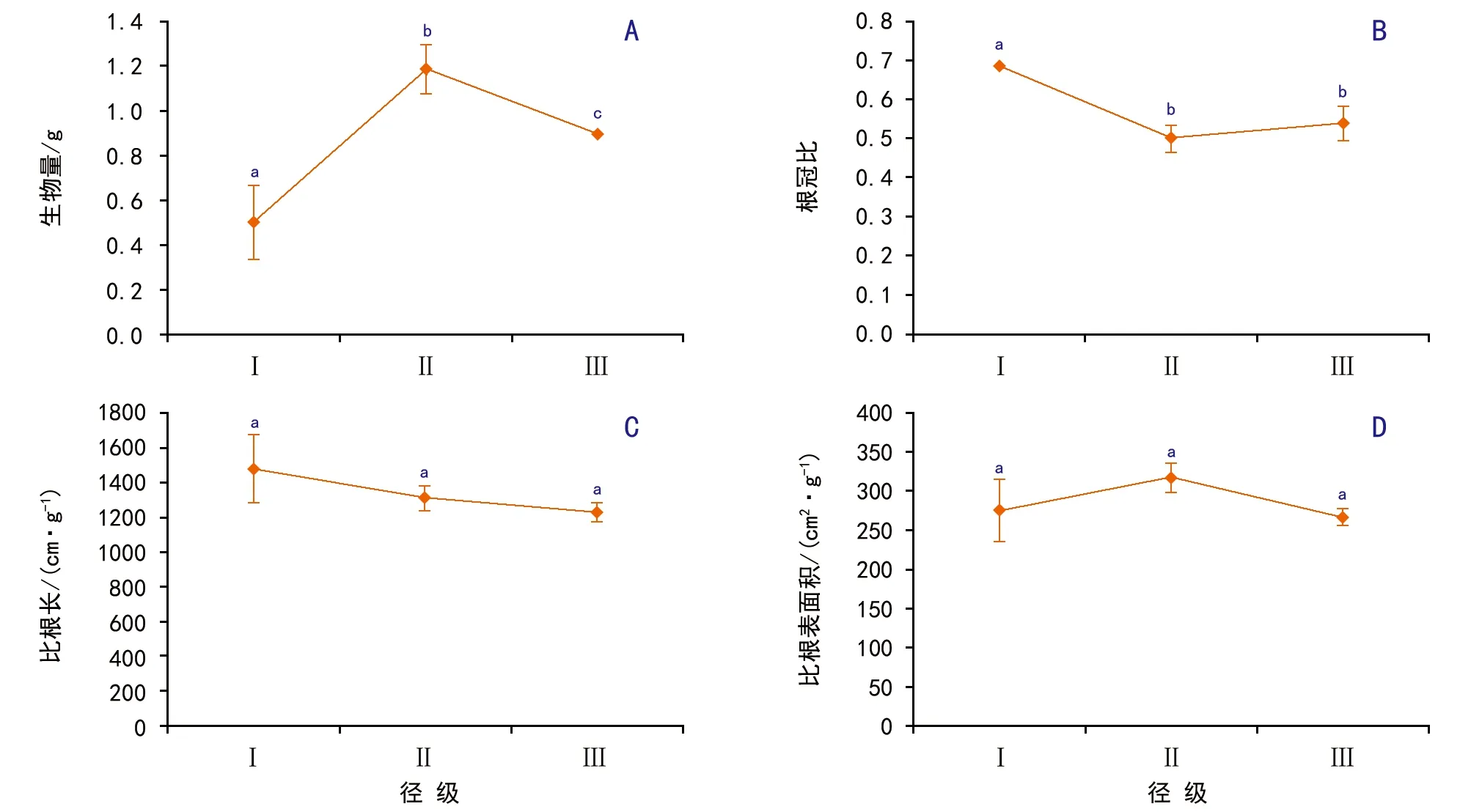
图5 母树径级对侧柏幼苗的生物量及其分配的影响
2.4 不同径级母树幼苗的生物量及分配
随母树径级的增加,幼苗干重和比根表面积先增加再降低,根冠比和比根长则是逐渐降低(图5)。方差分析结果表明,幼苗干重(F=27.00,p<0.01)和根冠比(F=27.30,p<0.01)在不同径级之间差异极显著,而比根长(F=3.20,p=0.11)差异不显著。对于幼苗干重是径级Ⅰ<径级Ⅲ<径级Ⅱ(p<0.05);对于根冠比,径级Ⅰ极显著大于径级Ⅱ与径级Ⅲ(p<0.01),而径级Ⅱ与径级Ⅲ差异不显著(p>0.05)。
3 结论与讨论
种子大小是反映其质量的重要特征。本研究发现,随母树径级的增加,侧柏种子千粒重逐渐增加,但变化不显著。马绍宾等也发现,多年生草本植物桃儿七在同一种群内单粒种子重量变化不大[15]。但水青树种子的干重[8]和四合木果实的重量[10]则随母树胸径(或基径)增加先增加再降低。太白红杉种子千粒重随随母树胸径增加而增加,王孝安等认为,与植株用于生殖的投资增多有关,而随后的降低则是因为植株变大引起树木间相互干扰加大导致生殖投资减少造成的[7]。本文中侧柏种子的千粒重在不同径级母树之间变异很大,在径级Ⅲ达到了49.49%,而种子萌发和幼苗生长中依然是径级Ⅲ的最高。吕文芳等发现,种子千粒重并不能代表种子的发芽能力和随后的幼苗生长水平[16]。而李怀春等也发现,水青树种子千粒重和发芽能力最大在母树胸径30~40 cm最大,而种子的长、宽、厚在母树胸径40~50 cm最大,二者表现不一致[8]。种子大小的变化可能主要与母树生长的环境[17]和生长状况[18]有关。而种子质量可能受到其它因素的影响,如母树遗传因素,所以有时种子大小与种子质量不一定完全一致。
种子萌发是植物开始天然更新的关键阶段之一。本文发现,随母树径级的增加,侧柏种子的发芽率、发芽指数逐渐增加,平均发芽时间逐渐降低,表明种子质量逐渐提升,这与Mao等[6]对黑松种子的研究结果一致。发芽时间的增加会使种子储藏养分过多消耗,导致不能萌发或幼苗细弱,影响幼苗成活率[19]。因此,侧柏大径级的母树形成的种子在萌发阶段具有明显的优势。但水青树[8]、胖大海[9]、Pinusechinata[11]和Sorbustorminalis[12]的种子发芽率则是在中间胸径的母树达到最大值。这可能与本文侧柏母树胸径的范围(9.2~21.2 cm)较小有关。对于林木种子发芽率的降低可能与母树对生殖分配减少有关[8-9]。香果树[13]、Piceamariana[14]的种子发芽率不受母树胸径的影响。侯聪等认为,不同水平密度疏伐的红松母树林内种子生活力没有显著差异,与遗传控制有关[20]。因此,树木在种子萌发上的差异除了与生长环境密切相关,也与母树相关的遗传因素有关。
生长是植物对外界不同胁迫的综合体现,反映了植物的适应能力[21]。本研究发现,不同径级侧柏的幼苗中,径级Ⅰ的地径、相对苗高生长速率和幼苗干重都显著低于径级Ⅲ。Mao等也发现,母树径级最大的黑松种子形成的幼苗有着最大的生物量和相对生长速率最大[6]。高的相对苗高生长速率有利于幼苗快速高生长,获得更多的光资源,提高与周边植物竞争的能力[22]。侧柏母树径级Ⅰ的根冠比极显著大于径级Ⅱ与径级Ⅲ。减少对地上部的投入,尤其是对叶片的投入,会降低植物的光合能力、生长速率[23]及在林下的光截获能力[24]。侧柏是喜光植物,快速的高生长有利于其在林窗或林缘快速生长进入冠层。因此,生长速率低、根冠比高不利于侧柏径级Ⅰ种子形成的幼苗在林下的建立。
母树大小对种子的质量有着显著的影响。本文研究随着径级的增加,侧柏种子的质量逐渐提升,发芽率和发芽指数逐渐增加,平均发芽时间逐渐降低,形成幼苗的生物量和生长速率也是大径级母树的最高,这对侧柏林下的幼苗建立具有重要意义。虽然侧柏不同径级母树之间种子的大小变异较大,但并没有影响种子的萌发和幼苗生长,说明种子的大小不能完全反映种子质量。这可能与种子在形成过程中受到母树生长环境、自身营养状况、遗传因素等多方面的共同影响有关。因此,在今后的研究中,深入开展侧柏母树生长环境和本身的生长状况对种子形成的影响,有助于准确判断种子质量,有利于提出合理有效的人工促进天然更新措施。
