失叶率对山乌桕幼苗生长和叶绿素含量的影响
2017-05-21周新华厉月桥罗嘉东肖智勇颜送宝
周新华,厉月桥,黄 拯,罗嘉东,肖智勇,颜送宝
(1.中国林业科学研究院 亚热带林业实验中心,江西 分宜 336600;2.宜春市林业科学研究所,江西 宜春 336000)
植物生长在开放的自然环境系统之中,它的生存环境并不总是适宜的,经常受到复杂多变的逆境胁迫,这对植物的生长发育将会造成显著的不利影响,为此,在长期的逆境生长过程中,植物通过组成型表达或损伤后诱导产生许多抵抗逆境的策略[1-3]。在植物和逆境的关系中,一方面逆境会对植物的生长产生不良的影响,另一方面植物也会对逆境采取相应的应对措施[4]。植物抗逆机制作用的发挥能够使植物在极端致死环境下保持存活,但这可能导致作物产量或生产力的降低[5]。食叶害虫以叶片为食,在食取叶片的时候可导致叶片破损或脱落,降低叶片的光合同化作用,从而导致植物光合生产力的下降,影响植物的生长量,国内外学者在食叶害虫方面做了许多相关研究[6-11]。
植物的生物量是单位面积植物积累物质的数量(干重:kg/hm2、g/m2,或能量:kJ/m2),植物生物量是生态系统的功能指标和获取能量能力的集中表现,它对生态系统机构和功能的形成具有十分重要的意义[12]。植物的光合作用是其产量的基础,林木90%以上的干物质来自光合作用,植物的叶片是植物进行光合作用来积累生物量的主要同化器官,叶绿素是叶片进行光合作用进行第一性生产的重要物质[13-15]。目前,国内许多学者在植物逆境生长和生物量的已经进行了相关研究报道[16-23],这些报道多是孤立地研究植物的抗逆生理机制或某一树种的生物量,而系统地开展某一逆境危害程度对植物生长量和生物量的研究未见报道。食叶害虫能够显著影响林木的初级生产力、能量流和养分循环,失叶是食叶害虫对林木造成危害的直接表现,准确定量地测定林木失叶率对林木生长的影响,可为区域病虫害诊断、森林灾害评估和病虫害控制提供重要依据。本文通过模拟食叶害虫取食叶片,研究了山乌桕Sapium discolor(Champ.ex Benth) Muell.Arg.在不同失叶率情况下,其生长和叶绿素含量的变化,为准确评估食叶害虫不同危害程度对山乌桕林生长影响、森林灾害评估和病虫害控制提供依据。
1 材料与方法
1.1 试验地概况
试验地点设置在中国林业科学研究院亚热带林业实验中心国家重点油茶良种繁育基地(新余市分宜县分宜镇,27°49′N,114°39′E)。基地年平均降水量1 643.6mm,年平均蒸发量1 503mm,年平均光照时数年光照时数1 535.3 h,年平均无霜期270 d,年平均气温17.2 ℃,最热月份7月的平均气温为28.8 ℃,极端最高温度39.9 ℃,最冷月份1月平均气温为5.5 ℃,极端最低温度-8.3 ℃,光热充足,冬寒期短,气候温和,属南亚热带湿润大陆性季风气候。
1.2 试验材料及处理方法
于4月上旬在中国林科院亚林中心大院内的国家重点油茶良种繁育基地内选择1块苗圃地作为试验开展的样地,用容器规格为12 cm×18 cm的无网袋轻基质栽植山乌桕当年种子萌发出来的实生苗,每个无网袋轻基质栽植1株,定期进行水、肥、光等的育苗管理。在山乌桕苗木生长3个月后,于7月上旬,分别选择生长良好、高矮一致、长势一致的幼苗进行试验布设,分为4组,每组100株,重复3次,对其中3组进行一次性剪叶处理,剪叶量分别为全株的25%、50%、75%,以不剪叶植株作为对照,方法参照高瑞桐等[24-25]、纪淑仁等[26]、姚双艳等[27]。山乌桕苗木失叶的处理方法为:失叶75%为自上而下每4片叶留1片叶,失叶50%为自上而下每2片叶留1片叶,失叶25%为自上而下每4片叶剪掉1片叶,摘除叶仅留叶柄,对照苗木不作任何处理,用于测量失叶率对山乌桕幼苗生长量和叶绿素含量的影响。
1.3 测定项目与方法
1.3.1 生长形态指标的测定
2015年9月上旬,在为期2个月的试验处理结束后,于上午8:00利用卷尺对苗木的株高进行测量,利用数显游标卡尺对苗木的地径进行测量,并记录数据;测量结束后,每一个处理随机选择10株长势良好、无病虫害的苗木,利用数显游标卡尺对苗木的叶长、叶宽进行测量,记录数据,并计算出相应的单叶面积、全株叶面积,每个试验处理重复3次。计算单叶面积和全株叶面积时,首先将叶片放入带有坐标纸的透明夹板上,利用电子数码相机拍照记录影像,用Photoshop CS5软件计算叶片的单叶面积和全株叶面积[28]。
1.3.2 生物量指标的测定
于试验结束后(2015年9月上旬),每个试验处理随机采集5株幼苗,重复3次,总共60株样品,将样品带回实验室,测量其株高、地径、根长;以根茎交界处为分界线,分为地上和地下部分,分别称量鲜质量,然后将样品材料剪碎处理后放入80 ℃恒温烘箱,烘干48 h后至恒质量,称量其干质量。
1.3.3 叶绿素含量SPAD的测定
使用便携式日产叶绿素测定仪:SPAD-502对各处理山乌桕苗叶绿素相对含量进行测定。在剪叶处理后的第15、30、45和60天取样,每个处理均选取健康完整的苗木,测定其成熟完全展开叶片的叶绿素相对含量,每处理一次测定10株,重复3次,取所测数据的平均值作为叶绿素相对含量数据。
1.4 数据处理
试验获得的数据全部采用平均数±标准误(mean±SE)来表示。数据的处理与统计分析采用Microsoft Office Excel 2003和SPSS 18.0社会统计分析软件来进行,使用显著性为0.05水平上的Duncan多范围检验来进行多重比较分析,所有数据在分析时均要统一量纲,确保分析的准确性。
2 结果与分析
2.1 不同失叶率对山乌桕生长形态指标的影响
由表1可以看出,不同程度的失叶率对山乌桕苗木生长形态的影响显著。随着失叶率的增加,山乌桕苗木的苗高表现出先增加后降低的趋势,具体的变化趋势为,失叶25%>CK>失叶50%>失叶75%,经过2个月的失叶处理,山乌桕失叶25%处理的苗木苗高比CK增加了1.59%,失叶50%和75%处理的苗木苗高分别比CK降低了7.90%和12.15%。
与苗高对各失叶处理的响应有所不同,山乌桕苗木的全株叶面积和地径生长量均随着失叶率的增加出现总体趋势下降的的情况(见表1),总的变化趋势为,CK>失叶25%>失叶50%>失叶75%,不同失叶率的山乌桕苗经过2个月的生长后,失叶25%、失叶50%和失叶75%处理的山乌桕植株苗木的地径和全株叶面积均显著低于对照CK植株。就地径和全株叶面积的减小幅度而言,在失叶率超过50%之后,其幅度随失叶程度的增加表现出减小的规律。
随着失叶程度的逐渐增加,山乌桕苗木的叶长、叶宽和单叶面积均表现出逐渐升高的趋势(见表1)。在模拟失叶处理经过2个月生长后,山乌桕失叶25%、50%和75%处理的植株的叶长分别比CK增加了4.03%、6.96%和6.65%,植株的叶宽分别比CK增加了4.34%、9.74%和9.29%,植株单叶面积分别比CK增加了0.65%、17.35%和16.53%。各失叶处理山乌桕苗木的叶长、叶宽和对照CK差异显著,各失叶处理山乌桕苗木的单叶面积,除失叶25%与对照CK差异不显著外,其它失叶处理均差异显著。
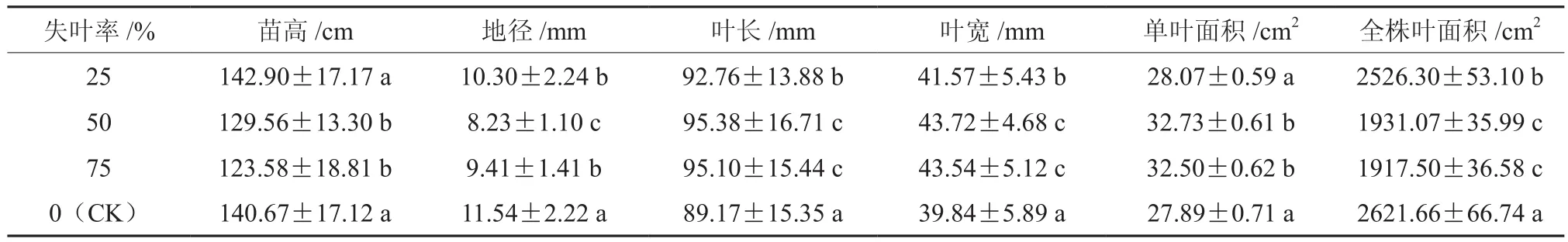
表1 不同失叶率情况下山乌桕苗木生长形态指标的差异†Table 1 Effects of different leaf area removal on the morphologic indexes in Sapium discolor
2.2 不同失叶率对山乌桕生物量的影响
由表2可以看出,不同程度的失叶率对山乌桕苗木生物量累积的影响显著。随着失叶率的增加,山乌桕苗木在叶鲜质量、茎鲜质量、根鲜质量、全株鲜质量、叶干质量、茎干质量、根干质量和全株干质量等8个指标方面均表现出降低趋势。经过2个月的失叶处理,失叶25%、失叶50%和失叶75%的植株叶鲜质量比CK减少了20.16%、31.68%和39.26%,植株茎鲜质量比CK分别减少了9.84%、26.40%和31.21%,植株根鲜质量比CK分别减少了21.32%、26.70%和17.85%,植株全株鲜质量比CK分别减少了16.30%、28.87%和33.63%,植株叶干质量比CK分别减少了17.26%、19.42%和25.12%,植株茎干质量比CK分别减少了22.71%、25.89%和24.27%,植株根干质量比CK分别减少了21.78%、24.75%和13.40%,植株全株干质量比CK分别减少了19.59%、22.18%和23.13%。就减少幅度而言,叶鲜质量、叶干质量、茎鲜质量、茎干质量、全株鲜质量和全株干质量随着失叶程度的增加而变大,而根鲜质量和根干质量在失叶率超过50%时却随着失叶程度的增加相应变小,这说明当失叶率超过一定程度时,植株通过自我调整生物量的分配,来消除或减小外界环境对其生长的不利影响,以便在逆境中更好的生存。
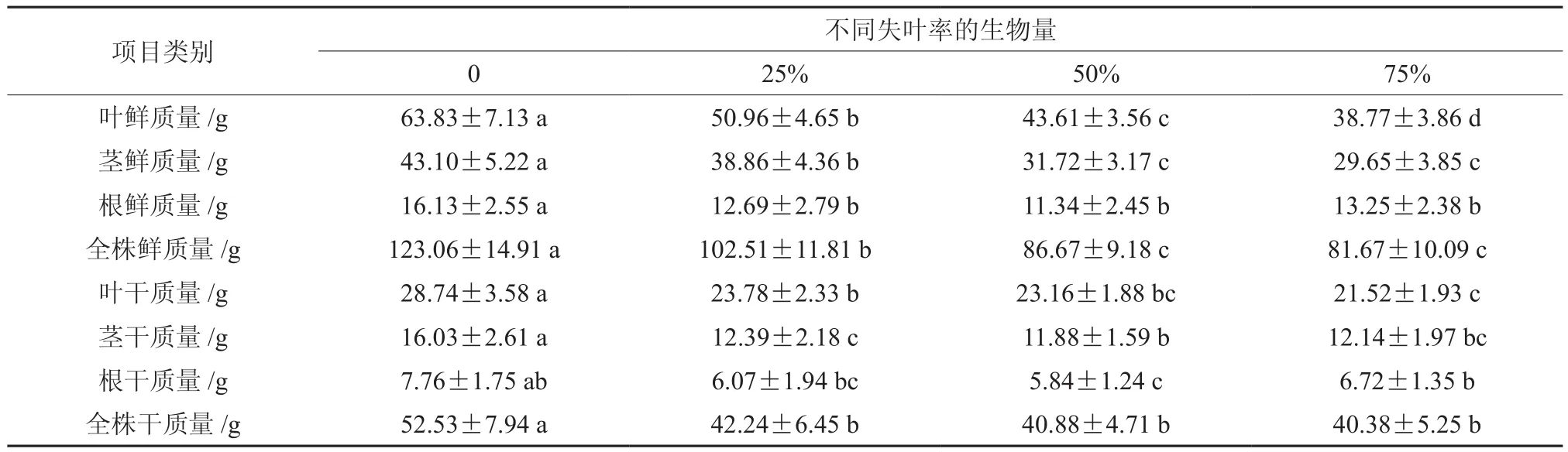
表2 不同失叶率情况下山乌桕苗木生物量的变化Table 2 Effects of different leaf area removal on the biomass in Sapium discolor g
2.3 不同失叶率对山乌桕叶绿素相对含量的影响
由表3可以看出,山乌桕苗木叶片中的叶绿素含量a要高于叶绿素含量b。失叶处理15d后,处理植株与CK植株相比叶绿素含量增多,失叶25%、50%、75%处理的植株叶绿素含量a和叶绿素含量a+b都显著高于CK,且失叶75%植株的叶绿素含量b显著高于CK及失叶25%和50%处理的植株;处理30d后,各失叶处理植株的叶绿素含量a、叶绿素含量b、叶绿素含量a+b均显著高于CK植株,且失叶75%植株的叶绿素含量b与其它失叶处理之间差异显著,此外,其它各处理之间差异不显著;处理45d后,失叶50%、失叶75%处理植株的叶绿素含量b显著高于CK和失叶25%,失叶75%处理的植株叶绿素含量a+b显著高于其它各处理;处理60d后,失叶50%、失叶75%处理植株的叶绿素含量a显著低于CK和失叶25%,失叶50%处理植株的叶绿素含量a+b显著低于CK,各处理之间叶绿素含量b差异不显著。随着处理时间的增加,各处理叶绿素含量与CK比较的变化情况表现为均表现为低-高-低。这说明植物为了维持自身生长发育的营养需求,在叶面积减少后会通过调整叶片中的叶绿素含量,来弥补光合面积的损失,以满足光合作用的需要。

表3 不同失叶率情况下山乌桕幼苗叶绿素指标的变化Table 3 Effects of different leaf area removal on the content of chlorophyll a,b and a+b in Sapium discolor
3 结论与讨论
(1)植物生长在开放的自然生态系统之中,但植物生长的最适环境往往很少存在,植物在其生长史中经常会遇到各种类型灾害的胁迫,而生长于各种不适宜的自然环境中,当环境胁迫解除之后,植物又逐渐恢复其生长,通过其自身的适应机制减少或消除灾害带来的不利影响。国内外学者在该机制下开展了不少植物在各种胁迫生境下的生长研究:杨志敏等[29]报道了泡桐在失叶10%时,其次年材积生长量超过对照;何学友等[30]报道了油茶失叶量超过25%时,对当年产量的影响不仅损失惨重,甚至绝产,而且可能导致树体死亡,对来年的产量影响也相当大。本研究结果表明,失叶对山乌桕生长形态指标具有较大影响:苗高随着失叶程度的增加表现出先增加和降低的趋势,除失叶25%的苗高与对照差异不显著外,其它处理均显著低于对照;全株叶面积和地径生长量均随着失叶程度的增加表现出总体下降的趋势,全株叶面积和地径生长量均显著低于对照;而叶长、叶宽和单叶面积均随着失叶程度的增加表现出上升的趋势,叶长、叶宽和单叶面积均显著高于对照。生长形态指标是植物生长过程中同化过程的外在体现,表现了植物适应胁迫环境的能力。山乌桕苗木在失叶后苗高、地径和全株叶面积等生长形态指标,并未按照失叶处理的比例小于对照,而是通过叶长、叶宽和单叶面积的补偿作用,来弥补光合面积的损失,从而降低苗木苗高、地径和全株叶面积等生长形态指标的减小幅度。
(2)食叶害虫的危害会影响苗木的生物量。本研究结果表明:各失叶处理植株的生物量均显著低于CK,但就减少幅度而言,山乌桕苗木的生物量与失叶程度之间并未呈现出相关性变化,这说明山乌桕苗木在受到伤害后,能够通过自身的补偿机制作用,来消除或减小不良环境对生物量造成的影响。
(3)食叶害虫的危害会影响苗木的叶绿素含量。本研究结果表明:山乌桕苗木的叶绿素含量与叶片受害程度呈现正相关,山乌桕在失叶后增加了叶绿素的含量,进而单位面积的光合作用得到加强,补偿了因失叶对光合作用的影响,弥补失叶对山乌桕生长和生物量造成的影响。山乌桕失叶后15~30 d,所有失叶处理植株的叶绿素含量均高于CK,光合能力提高,使得各失叶处理的苗木之间生物量无明显差异,亦使得光合产物与失叶程度之间未呈现负相关性变化。山乌桕失叶后45~60 d,失叶75%处理植株的叶绿素含量显著高于CK,光合能力得到提高,使其在长期的生长过程之中生物量与25%和50%的植株无显著差异。
(4)本研究结果表明,林木在受到食叶害虫的危害后,一方面通过调整叶长、叶宽和单叶面积等生长形态指标,来弥补光合面积的损失;另一方面又能通过增加叶片中的叶绿素含量,提高单位面积的光合效率,来消除光合面积减小的影响,但是对失叶的耐受能力和补偿能力是有限的,且不同树种应对失叶的补偿能力存在巨大差异。在山乌桕失叶超过25%时,其苗高生长受到明显遏制,必需采取有效措施防止食叶害虫的进一步危害。本研究只是针对不同失叶率对山乌桕苗木生长形态和生物量造成的影响开展的,没有深入模拟研究食叶害虫取食过程中的植株变化,这有待于未来进一步研究。
参考文献:
[1]徐慧妮,王秀峰.植物生长发育逆境胁迫及其研究方法[C]//中国园艺学会.中国园艺学会第七届青年学术讨论会论文集,中国园艺学会:2006:7.
[2]Kempel A,Schadler M,Chrobock T,et al.Tradeoffs associated with constitutive and induced plant resistance against herbivory[J].Proceedings of the National Academy of Sciences,2011,108(14):5685-5689.
[3]张亚军,王丽学,陈 超,等.植物对逆境的响应机制研究进展[J].江西农业大学学报,2011,23(9):60-65.
[4]李媛媛,张 凯,李霜雯,等.失叶率对小黑杨和兴安落叶松幼苗生物量和叶绿素含量的影响[J].林业科学,2015,51(3):93-101.
[5]杨广新,李 胜,李 唯,等.几种外源含氮化合物对草麻黄愈伤组织增殖和麻黄碱含量的影响[J].植物生理学通讯,2007,43(1):49-52.
[6]林丽静,陈志云,李奕震.我国桉树主要食叶害虫的危害及防治[J].桉树科技,2006,23(1):45-48.
[7]孙志强,杜红岩,李芳东.杜仲集约化栽培潜在的病虫灾害及其应对策略[J].经济林研究,2011,19(4):70-76.
[8]赵 星,周艳初,喻传明.竹类食叶害虫的研究及进展[J].经济林研究,1992,10(2):79-81.
[9]袁 菲,刘 曦,赵佳丽,等.汪清林区天然林食叶害虫危害与林分的关系[J].林业科技开发,2014,28(1):55-58.
[10]胡春祥,胡 扬,谢淑萍,等.12种生物无公害药剂对3种杨树食叶害虫的毒力测定及防治[J].东北林业大学学报,2007,35(5):42-45.
[11]蔡金峰,曹福亮,张往祥.淹水胁迫对乌桕幼苗生长及根系无氧呼吸酶活性的影响[J].中南林业科技大学学报,2013,33(9):5-10.
[12]郭 娜,刘剑秋.植物生物量研究概述[J].亚热带植物科学,2011,40(2):83-88.
[13]李启明.植物光合效率与产量的关系及影响光合效率的内在因子[J].植物生理学通讯,1982,(4):34-38.
[14]周新华,朱宜春,潘文婷,等.不同油茶无性系苗期光合效率的研究[J].江西农业大学学报,2015,37(3):461-465.
[15]许晨璐,孙晓梅,张守攻.3种落叶松苗期光能利用效率的比较[J].林业科学研究,2014,27(4):565-569.
[16]刘 杰,崔保山,杨志峰,等.纵向岭谷区高速公路建设对沿线植物生物量的影响[J].生态学报,2006,26(1):83-90.
[17]陈建明,俞晓平,程家安,等.植物耐虫性研究进展[J].昆虫学报,2005,48(2):262-272.
[18]徐呈祥.等渗透式干旱、盐、碱胁迫下5个枣品种及酸枣的生物学相应与抗逆性[J].中南林业科技大学学报,2012,32(12):139-146.
[19]侯晓龙,刘爱琴,蔡丽平,等.异质Pb胁迫对超富集植物金丝草根系生长及生物量分配的影响[J].农业环境科学学报,2012,32(11):2145-2150.
[20]童 琳,唐旭利,张 静,等.菌根真菌侵染对植物生物量累积的影响[J].生态环境学报,2014,23(9):1520-1525.
[21]姜卫兵,郝胜大,翁忙玲,等.逆境对木兰科树种伤害机理研究进展[J].生态环境,2008,17(1):439-466.
[22]李玉梅,郭修武,代汉萍,等.盐碱胁迫对牛叠肚幼苗渗透调节物质及叶绿素含量的影响[J].经济林研究,2015,33(1):9-16.
[23]龚洪恩,吕芳德,陈传松,等.离子注入对伞树幼苗叶绿素含量的影响[J].经济林研究,2014,32(1):140-143.
[24]高瑞桐,秦锡祥,陈德钧,等.摘叶模拟食叶害虫危害对杨树生长的影响[J].林业科学研究,1994(6):709-712.
[25]高瑞桐,秦锡祥,李吉震,等.食叶害虫的食叶量与树木生长的关系[J].林业科学,1985,21(2):199-205.
[26]纪淑仁,夏 槃.夏大豆食叶害虫的为害程度与产量损失关系的模拟研究[J].安徽农业科学,1986(4):64-68.
[27]姚双艳,毛 培,韩 笑,等.几种杀虫剂对4种鳞翅目食叶害虫的毒力和模拟田间防效[J].农药,2015(7):533-536.
[28]刘泽彬,程瑞梅,肖文发,等.遮荫对中华蚊母树苗期生长及光合特性的影响[J].林业科学,2015,51(2):129-136.
[29]杨志敏,俞志顺,杨焕金.大袋蛾危害泡桐的经济阀值研究[J].山东林业科技,1991(3):47-50.
[30]何学友,蔡守平,谢一青,等.不同叶面积损失对油茶产量及品质的影响[J].林业科学,2013,49(5):85-91.
