稻米垩白性状对高温耐性的QTL分析
2017-05-19张桂莲廖斌唐文帮陈立云肖应辉
张桂莲 廖斌 唐文帮 陈立云肖应辉
(湖南农业大学农学院,长沙410128;*通讯联系人,E-mail:chenliyun996@163.com,xiao_yh@163.com)
稻米垩白性状对高温耐性的QTL分析
张桂莲 廖斌 唐文帮 陈立云*肖应辉*
(湖南农业大学农学院,长沙410128;*通讯联系人,E-mail:chenliyun996@163.com,xiao_yh@163.com)
【目的】本研究旨在筛选与稻米外观品质高温耐性连锁的分子标记,为稻米品质育种提供参考。【方法】以耐热水稻品系996和热敏感水稻品系4628为亲本构建的重组自交系为材料,采用垩白粒率耐热指数、垩白大小耐热指数和垩白度耐热指数为评价指标,对水稻垩白性状的高温耐性QTL进行检测。【结果】采用复合区间作图法两年共检测到垩白性状高温耐性QTL 24个,包括垩白粒率高温耐性QTL 8个,垩白大小高温耐性QTL 12个,垩白度高温耐性QTL 4个。其中,第6染色体上的2个垩白粒率高温耐性QTL和第7染色体上的2个垩白度高温耐性QTL在两年中重复检测到,且这2个稳定表达的垩白度位点与2015年检测到的第7染色体上的垩白粒率位点重合。另外,发现有4个QTL一因多效,同时影响垩白粒率、垩白大小及垩白度。【结论】控制垩白粒率耐热指数的qHTCGR6.1和控制垩白度耐热指数的qHTCD7.1是新的QTL。
稻米;垩白性状;高温耐性;数量性状基因座位
水稻是我国主要粮食作物,在保障国家粮食安全中占有举重轻重的地位。随着人们生活水平提高,对稻米品质的要求也越来越高。虽然近几年我国水稻品种的稻米品质有较大提高,但还远满足不了市场的需求。如林海等[1]对2011-2015年通过审定水稻品种主要米质指标分析发现,粳稻品质提高幅度较大,而籼型两系杂交水稻和籼型三系杂交稻米质总体表现较差,优质达标率5年平均分别为39.10%和32.65%,垩白粒率、垩白度优质达标率5年平均值分别为38.36%和6.6%。同样,超级杂交稻的垩白粒率及垩白度优质达标率仍然较低,仅为34.7%和51.0%[2]。垩白粒率、垩白度仍是制约我国杂交水稻品质提高的主要因素。因此,减少垩白、改善稻米的外观品质,增加稻米的商品价值,是我国稻米品质改良中亟待解决的一个关键问题。
近年来,全球气候变暖,地球平均气温升高,极端高温天气发生的频率增大,在世界上许多地区高温已成为影响作物生长的主要因素[3]。我国长江以南双季早稻的开花结实期及中稻开花期经常遭遇异常高温天气,不仅影响水稻的受精结实,导致大幅度减产[4,5],而且稻米品质显著降低,主要表现为外观品质变差,即垩白粒率增加、垩白度增大。大量的研究表明,灌浆成熟期的气温对稻米品质影响极大[6-8]。高温热害已成为限制我国南方稻区水稻优质安全生产的瓶颈。因此,培育和推广对环境温度弱感或钝感的广适性水稻品种,是克服高温胁迫的最有效措施。已有研究表明,稻米品质形成对环境温度的敏感性存在基因型差异,但由于缺乏对其遗传基础的了解,有关稻米品质对温度耐性的遗传改良报道很少。因此,本研究利用水稻耐热品系996和热敏感品系4628为亲本构建的重组自交系群体为材料,对垩白性状进行QTL定位分析,旨在揭示稻米外观品质的遗传机理,筛选与稻米外观品质高温耐性连锁的分子标记,以应用于稻米品质育种。
1 材料与方法
1.1 供试材料
供试材料为耐热水稻品系996、热敏感水稻品系4628及其124个重组自交系群体。该重组自交系群体在以996/4628 F1自交分离获得F2群体的基础上,通过单粒传法构建而成。
1.2 材料种植与温度处理
2014年和2015年分别在湖南农业大学耘园试验地种植耐热品系996、热敏感品系4628、996/4628重组自交系群体。2014年3月27日播种,5月1日移栽,采用单本移植,每一株系种植30株,移栽密度为17 cm×20 cm;2015年3月27日播种,5月4日移栽,采用单本移植,每一株系种植30株,移栽密度为17 cm×20 cm。按照常规水稻栽培管理技术进行田间管理,在水稻幼穗分化第5期左右,选取生长进程和分蘖数目基本一致水稻植株(边行不取样)移入盆钵中(盆钵内径266 mm,高190 mm),移栽时植株根部带土(根部带土量基本一致)移入,每盆3株,每株系4盆,盆内加水让其恢复生长。始穗当日上午7:00-8:00标记抽穗分蘖,在抽穗后8 d移入湖南农业大学水稻研究所人工气候室进行高温(8:00-17:00,37℃,17:00-8:00,30℃)和适温(8:00-17:00,30℃,17:00-8:00,25℃)处理。处理期间人工气候室相对湿度为75%,植株冠层光通量密度为850µmol/(m2·s)。处理20 d后移出人工气候室,在室外恢复生长3 d,至成熟期收获标记穗,脱粒,于阴凉通风处自然干燥3个月后进行垩白性状考察。每株系每处理处理两盆,每1盆为1次重复,共2次重复。
1.2 表型测定项目及方法
1.2.1 垩白粒率、垩白大小和垩白度的测定
从平均样品中随机数取试样100粒,3次重复。逐粒目测,拣出试样中具有明显的、白色透明的垩白米粒,然后统计垩白米粒数目,计算垩白粒率(垩白米粒占试样总粒数的百分率),从中选取100粒采用平面方格法,逐粒目测垩白面积占整粒米平面投影面积的百分率,然后用加权法计算试样平均垩白大小。然后,根据公式计算垩白度(垩白度=垩白粒率×垩白大小×100)。
1.2.2 垩白性状耐热指数的计算
垩白性状高温耐性以耐热指数表示,计算公式如下:
垩白粒率耐热指数=[(高温下垩白粒率-适温下垩白粒率)/垩白粒率转换指数]×100;垩白粒率转换指数=[(亲本高温下垩白粒率-亲本适温下垩白粒率)/亲本高温下垩白粒率]×100;
垩白大小耐热指数=[(高温下垩白大小-适温下垩白大小)/垩白大小转换指数]×100;垩白大小转换指数=(亲本高温下垩白大小-亲本适温下垩白大小)/亲本高温下垩白大小×100;
垩白度耐热指数=[(高温下垩白度-适温下垩白度)/垩白度转换指数)×100;垩白度转换指数=[(亲本高温下垩白度-亲本适温下垩白度)/亲本高温下垩白度]×100。
以2个重复(盆钵)耐热指数的平均值作为高温耐性的评价指标。
1.3 DNA提取及基因型分析方法
1.3.1 DNA样品制备和SSR分析
在水稻分蘖期对耐热品系996、热敏感品系4628及996/4628重组自交系群体各株系取3~4片新鲜幼嫩叶片,液氮速冻后保存在-80℃低温冰箱中备用。DNA样品的制备参照Saghai等[9]的方法。提取的DNA溶解于TE缓冲液(10 mmol/L Tris, 0.1 mmol/L EDTA)。每份DNA统一用ddH2O稀释成20 ng/L的浓度,作为PCR分析的模板。
根据已有文献报道的引物序列[10-14],共计选取了991对SSR引物和66对SFP引物对亲本的多态性进行了筛选。SSR和SFP分析参照Chen等[10]的程序,10 μL反应体系包含10 mmol/L Tris-HCl(pH 8.3),50 mmol/L KCl,1.5 mmol/L MgCl2,50 μmol/LdNTPs,0.2 μmol/L引物,0.5 U Taq聚合酶(天根生化科技有限公司生产)和20 ng DNA模板。扩增反应在美国ABI-2700 PCR扩增仪上进行:94℃下预变性5 min;94℃下30 s;55℃下30 s,72℃下1 min,共35个循环;最后72℃下延伸7 min。扩增产物用8%的非变性聚丙烯酰胺凝胶分离,银染显色。DNA条带利用装有荧光灯的灯箱进行观察。
1.3.2 遗传连锁图构建及QTL分析
根据两个群体中各株系PCR产物的电泳带型,将与996带型相同的个体记为“1”,与4628带型相同的个体记为“2”,表现双亲带型的杂合个体记为“3”,空缺的条带记为“-”,并进行数据记录。运用MapMaker 3.0软件[15]对整理后的数据进行分析,计算分子标记间的遗传距离,并绘制分子标记遗传连锁图谱(LOD阈值取3.0)。利用已构建的分子标记遗传图谱[16],结合各耐热表型数据,使用WinQTL Cartographer 2.5软件[17],用复合区间作图法(CIM)进行QTL分析。并分析每个QTL可解释表型变异的百分率。将LOD值2.5作为阈值,若标记区间LOD≥2.5,则认为该区间LOD值的最高处对应的位点为该性状的1个QTL,并分析各个QTL的贡献率。
2 结果与分析
2.1 亲本与RIL群体的表型
从表1和图1可看出,在垩白粒率、垩白大小、垩白度性状方面,亲本996和4628耐热指数差异大,且不同年份表现基本一致。而重组自交系群体各株系的垩白粒率、垩白度、垩白大小的耐热指数均表现为正态连续性分布,同一性状两年间的频率分布特征基本一致,但2015年各性状值多大于2014年,这可能是不同的环境条件能够影响水稻垩白性状的分布。
2.2 垩白粒率高温耐性QTL定位
2014年在重组自交群体中检测到垩白粒率高温耐性QTL共6个,分别位于第1、第3、第6和第7染色体上(表2)。QTL的贡献率变化范围为7.91%~17.89%,加性效应值变幅为0.02~0.06;2015年在重组自交系群体中检测到垩白粒率高温耐性QTL共4个,分别位于第6、第7染色体上,QTL的贡献率的变化范围为7.66%~14.25%,加性效应值变幅为0.03~0.06。位于第6染色体上的RM3353-RM1369标记区间内的qHTCGR6.1及RM1369-RM190标记区间内的qHTCGR6.2在2014与2015年间均被检测到。qHTCGR6.1、qHTCGR6.2的LOD值分别为4.44和7.24;贡献率分别为14.24%和17.89%。
2.3 垩白大小高温耐性QTL定位
2014年在重组自交群体中检测到垩白大小高温耐性QTL共7个,分别位于第1、第2、第10染色体上,其中第1染色体上分布1个QTL,第2染色体上分布2个QTL,第10染色体上存在4个QTL(表2)。QTL的贡献率变化范围为10.30%~15.08%,加性效应值变幅为-0.14~0.17。2015年在重组自交系群体中检测到垩白大小高温耐性QTL共5个,分别位于第4、第6、第7染色体上,第4染色体上分布1个QTL,其中第6和第7染色体上各分布2个QTL,这5个QTL的贡献率的变化范围在6.91%~18.76%,其加性效应值变幅为-0.25~0.17。所有垩白大小高温耐性QTL均只被检测出一次,两年间没有相同的QTL被检测出来。位于第6染色体上的qHTCS6.1、qHTCS6.2和第7染色体上的qHTCS7.1、qHTCS7.2与垩白粒率高温耐性的位点重合。
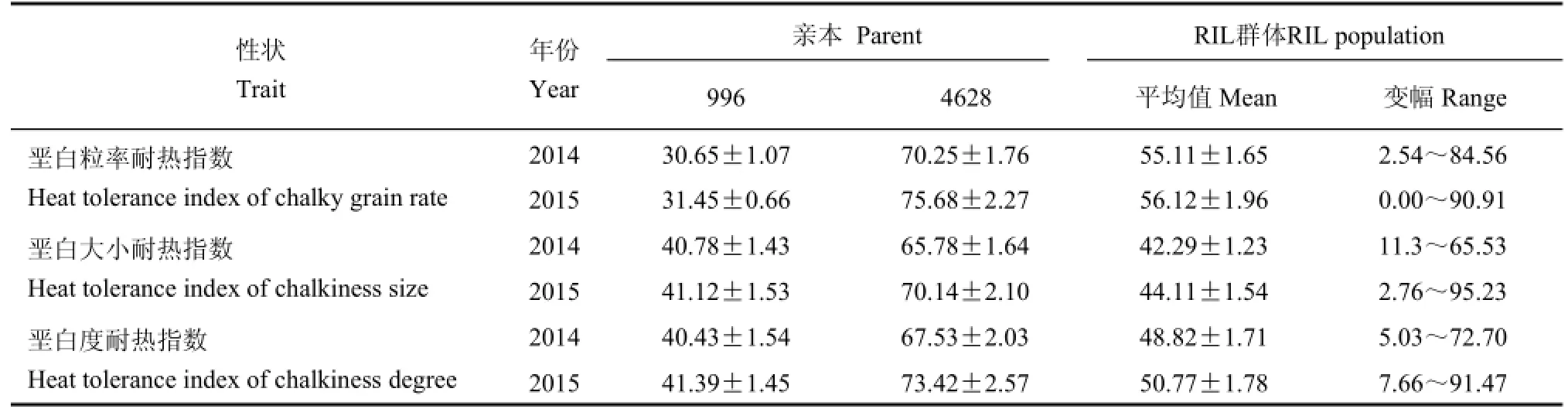
表1 双亲及RIL群体各株系垩白粒率、垩白大小和垩白度耐热指数Table 1.Heat tolerance index of chalky grain rate,chalkiness size and chalkiness degree of two parents and their RIL population.

图1 2014年和2015年RIL群体垩白粒率、垩白大小、垩白度耐热指数分布Fig.1.Frequency distribution of heat tolerance index of chalky grain rate,chalkiness size and chalkiness degree in RIL population in 2014 and 2015.
2.4 垩白度高温耐性QTL定位
2014年在重组自交群体中检测到4个垩白度高温耐性QTL,分别位于第1、第3、第7染色体上,其中2个QTL位于第7染色体上(表2),QTL的贡献率变化范围为11.54%~36.34%,加性效应值变幅为-0.17~-0.10;2015年在重组自交系群体中检测到2个垩白度高温耐性QTL,分布在第7染色体上,贡献率分别为17.44%、29.52%,加性效应值变幅为-0.14~-0.17。位于第7染色体上RM21327 -RM21364标记区间内的qHTCD7.1和RM21364 -RM3859标记区间内的qHTCD7.2在两年均被重复检测出来,2014年贡献率分别为13.27%、16.32%,2015年贡献率分别为17.44%、29.52%。而且这两个位点与垩白粒率、垩白大小的位点重合,表明垩白度与垩白粒率、垩白大小间存在高度相关的内在分子基础。
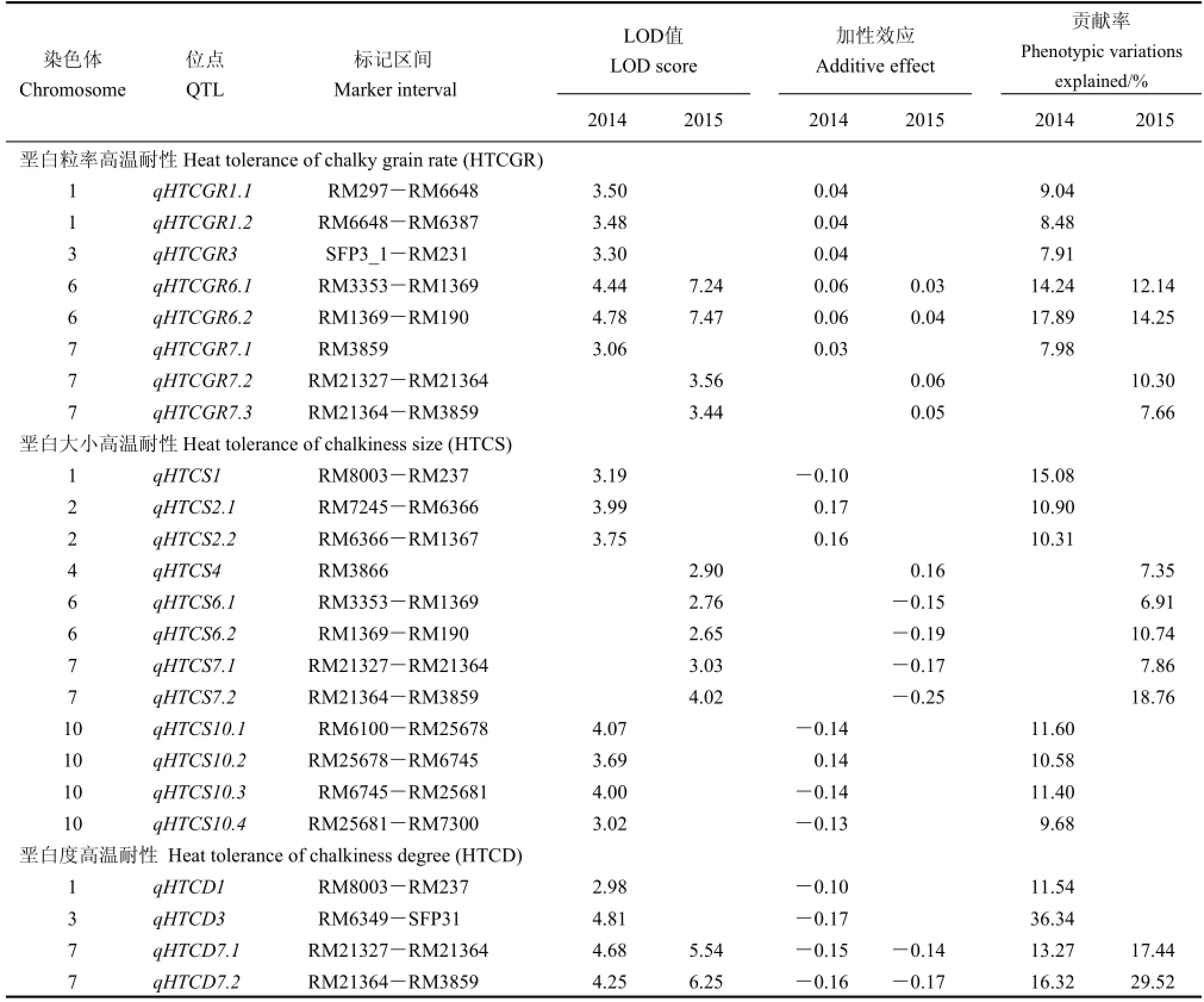
表2 垩白性状高温耐性QTL分析Table 2.Mapping of heat tolerance QTL associated with grain chalkiness traits.
3 讨论
水稻垩白性状包括垩白粒率、垩白面积、垩白度,垩白性状属于数量性状,有多个QTL共同调控垩白性状的表达,并且与环境存在明显的互作效应[18-24]。高温是影响水稻垩白形成的最主要气候因素,灌浆结实期高温胁迫明显导致水稻籽粒垩白粒率上升,垩白面积增大,垩白度增加[25-28]。高温胁迫已成为水稻优质的重要限制因素。本研究以996/4628 RIL为供试材料,对水稻籽粒垩白粒率高温耐性QTL、垩白大小高温耐性QTL、垩白度高温耐性QTL进行分析。研究结果显示,在2014-2015年2年内共检测到垩白性状高温耐性QTL 24个,其中垩白粒率高温耐性QTL 8个,垩白大小高温耐性QTL 12个,垩白度高温耐性QTL 4个。在2014 -2015年,2个垩白粒率高温耐性QTL qHTCGR6.1,qHTCGR6.2和2个垩白度高温耐性QTL qHTCD7.1,qHTCD7.2在两年均被重复检测到,这表明这4个QTL位点在不同年份环境中能稳定表达。其余控制垩白性状的QTL位点只能在一个年份中检测到,属于对环境敏感型。其次,位于第7染色体上的2个垩白度位点与垩白粒率的位点重合,表明这2个位点不仅能调控垩白粒率,而且对垩白度也有一定的影响,这2个性状可能存在相同的遗传基础[29-30]。
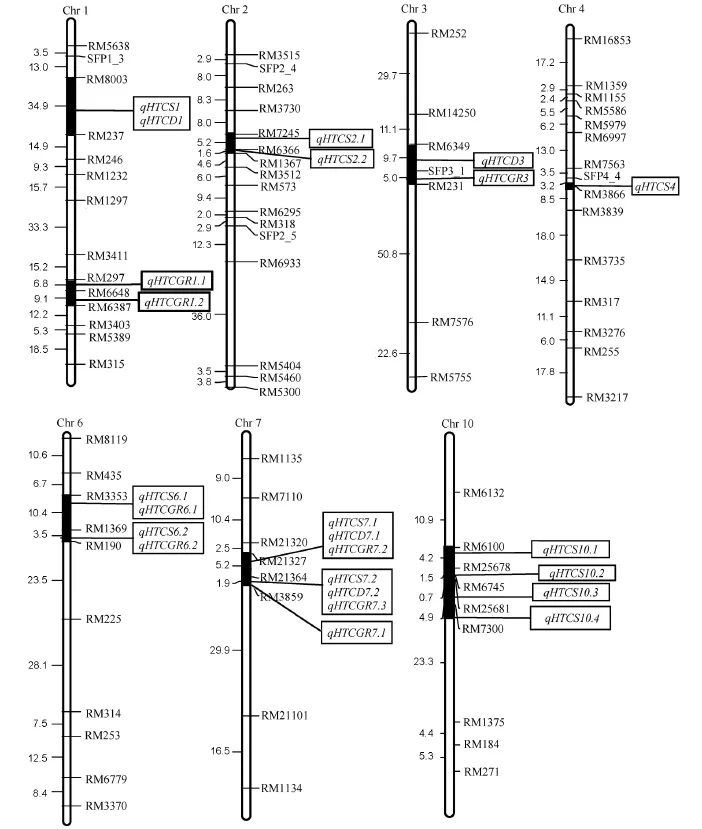
图2 检测到的垩白性状高温耐性QTL在染色体上的分布Fig.2.Chromosome location of putative QTL for heat tolerance of grain chalkiness traits.
同一性状的检测结果在不同群体中可能不同;同一群体的同一性状QTL检测结果在不同年份或地点中可能不同;同一群体的,同一年份的,同一地点的同一性状QTL检测结果可能因不同播始历期而不同。因此,重复性高,贡献率大的QTL往往存在较大的挖掘空间。在本研究中,位于第6染色体RM1369-RM190标记区间的qHTCGR6.2与盘毅等[31]利用本研究的同一个群体在第6染色体上检测到控制花粉育性高温耐性QTL(标记区间为RM190-RM225)和Yamakawa等[32]定位的位点qPGWC-6相邻,与高方远等[33]定位的控制垩白粒率性状表达的基因位点重合。这说明标记RM190附近可能存在同时控制垩白粒率、垩白粒率高温耐性及花粉育性高温耐性基因。位于第7染色体RM21364-RM3859标记区间的qHTCD7.2与杨亚春等[22]定位的垩白粒率QTL qCR7-1标记区间RM481-RM3859相邻,这说明标记RM3859附近可能存在同时控制垩白性状及垩白性状高温耐性基因。位于第6染色体RM3353-RM1369标记区间内的qHTCA6.1和位于第7染色RM21327-RM21364标记区间内的qHTCD7.1在两年中稳定表达,目前未有相关报道,可能为新的QTL位点,可为分子标记辅助选择培育耐热优质水稻品种提供新的基因资源。
[1]林海,庞乾林,王志刚,鄂志国.2015年我国审定的水稻品种基本特性分析.中国稻米,2016,22(6):4-9. Lin H,Pang G L,Wang Z G,E Z G.Analysis on characteristics of rice varieties registered in China in 2015.China Rice,2016,22(6):4-9.(in Chinese)
[2]闵捷,朱智伟,章林平,陈能,许立,牟仁祥.中国超级杂交稻组合的稻米品质分析.中国水稻科学,2014, 28(2):206-210 Min J,Zhu Z W,Zhang L P,Chen N,Xu L,Mou R X. Analysis on milled rice quality of super hybrid rice combinations in China.Chin J Rice Sci,2014,28(2): 206-210.(in Chinese with English abstract)
[3]IPCC.Climate Change 2001:Scientific Basis.New York, USA:Cambridge University Press,2001.
[4]夏明元,戚华雄.高温热害对四个不育系配制的杂交组合结实率的影响.湖北农业科学,2004(2):21-22. Xia M Y,Qi H X.Effect of high temperature heat stress on seed setting rate of hybrid combinations from four sterile lines.Hubei Agric Sci,2004(2):21-22.(in Chinese)
[5]杨惠成,黄仲青,蒋之埙,王相文.2003年安徽早中稻花期热害及防御技术.安徽农业科学,2004,32(1):3-4. Yang H C,Huang Z Q,Jiang Z X,Wang X W.Heat damage and defense technology of early and middle rice at flowering stage in Anhui in 2003.Anhui Agric Sci, 2004,32(1):3-4.(in Chinese with English abstract)
[6]郑建初,张彬,陈留根,杜群,秦永生,宋健,张卫建.抽穗期高温对水稻产量构成要素和稻米品质的影响及其基因型差异.江苏农业学报,2005,21(4):249-254. Zheng J C,Zhang B,Chen L G,Du Q,Qin Y S,Song J, Zhang W J.Genotypic differences in effects of high temperature in field on rice yield components and grain quality during heading stage.Jiangsu J Agric Sci,2005, 21(4):249-254.(in Chinese with English abstract)
[7]谢晓金,李秉柏,李映雪,李昊宇,赵小艳,杨沈斌,王志明.抽穗期高温胁迫对水稻产量构成要素和品质的影响.中国农业气象,2010,31(3):411-415. Xie X J,Li B B,Li Y X,Li H Y,Zhao X Y,Yang S B, Wang Z M.Effects of high temperature on rice yield components and grain quality during heading stage.Chin Agric Agrometeorol,2010,31(3):411-415.(in Chinese with English abstract)
[8]石军,褚旭东,王志,黄廷友,李春财.自然高温对15个籼稻杂交组合稻米品质的影响.湖北农业科学,2011, 50(5):897-899. Shi J,Chu X D,Wang Z,Huang T Y,Li C C.Effect of natural high temperature on rice quality of 15 indica hybrid combinations.Hubei Agric Sci,2011,50(5): 897-899.(in Chinese)
[9]Saghai-Maroof M A,Biyashev R M,Yang G P,Zhang Q. ExtraordinarilypolymorphicmicrosatelliteDNAin barley:Species diversity,chromosomal locations,and population dynamics.Proc Natl Acad Sci USA,1994, 91(12):5466-5470.
[10]Chen X,Temnyk H S,Xu Y,Cho Y G,McCouch S R. Developmentofamicrosatelliteframeworkmap providing genome-wide coverage in rice(Oryza sativa L.). Theor Appl Genet,1997,95:553-567.
[11]Temnykh S,Park W D,Ayres N,Sam Cartinhour,Hauck N,Lipovich L,Cho Y G,Ishii T,McCouch S R.Mapping and genome organization of microsatellite sequences in rice(Oryza sativa L.).Theor Appl Genet,2000,100: 697-712.
[12]McCouch S R,Teytelman L,Xu Y B,Lobos K B,Clare K,Walton M,Fu B Y,Maghirang R,Li Z K,Xing Y Z, Zhang Q F,Kono I,Yano M,Fjellstrom R,DeClerck G, SchneiderD,CartinhourS,WareD,SteinL. Development and mapping of 2240 new SSR markers for rice(Oryza sativa L.).DNA Res,2002,9:199-207.
[13]International Rice Genome Sequencing Project.The map-based of the rice genome.Nature,2005,436: 793-800.
[14]Edwards J D,Janda J,Sweeney M T,Gaikwad A B,Liu B,Leung H,Galbraith D W.Development and evaluation of a high-throughput,low-cost genotyping platform based on oligonucleotide microarrays in rice.Plant Meth,2008, 4:13.
[15]Lincoln S,Daley M,Lander E.Constructing genetic maps with MAPMAKER/EXP 3.0//Whitehead Institute TechnicalReport.3rded.Cambridge:Whitehead Institute,1992.
[16]何云丽,叶乃忠,郝明,罗丽华,肖应辉.多环境下早籼稻重组自交系群体的抽穗期QTL分析.中国水稻科学,2013,27(4):389-397. He Y L,Ye N Z,Hao M,Lu L H,Xiao Y H.QTL analysis for heading date by using recombinant inbred linesderivedfromearly-seasonindicariceacross multi-environments.ChinJRiceSci,2013,27(4): 389-397.(in Chinese with English abstract)
[17]Zeng Z B.Precision mapping of quantitative trait loci. Genetics,1994,136:1457-1468.
[18]Mei D Y,Zhu Y J,Yu Y H,Fan Y Y,Huang D R, Zhuang J Y.Quantitative trait loci for grain chalkiness andendospermtransparencydetectedinthree recombinant inbred line populations of indica rice.J Integr Agric,2013,12:1-11.
[19]周立军,刘喜,江玲,郑蕾娜,陈亮明,刘世家,翟虎渠,万建民.利用CSSL和BIL群体分析稻米垩白粒率QTL及互作效应.中国农业科学,2009,42(4): 1129-1135. Zhou L J,Liu X,Jiang L,Zheng L N,Chen L M,Liu S J, Zhai F Q,Wan J M.Analysis of QTL and GE effects on PGWC in rice(Oryza sativa L.)using CSSL and BIL populations.Sci Agric Sin,2009,42(4):1129-1135.(in Chinese with English abstract)
[20]Liu X,Wang Y and Wang S W.QTL analysis of percentage of grains with chalkiness in Japonica rice (Oryza sativa L).Gen Mol Res,2012,11:717-724.
[21]晁园,冯付春,高冠军,朱雪萍,何予卿.利用重组自交系群体定位水稻品质相关性状的QTL.华中农业大学学报,2012,31(4):397-403. Chao Y,Feng F C,Gao G J,Zhu X P,He Y Q.Mapping QTLs related with rice qualities of appearance,cooking and eating using a recombinant inbred line population.J Huangzhong Agric Univ,2012,31(4):397-403.(in Chinese with English abstract)
[22]杨亚春,倪大虎,宋丰顺,李泽福,易成新,杨剑波.不同生态地点下稻米外观品质性状的QTL定位分析.中国水稻科学,2011,25(1):43-51. Yang Y C,Ni D H,Song F S,Li Z F,Yi C X,Yang J B. Identification of QTLs of rice appearance quality traits across different ecological sites.Chin J Rice Sci,2011, 25(1):43-51.(in Chinese with English abstract)
[23]王林森,陈亮明,王沛然,王卓然,郑海,马宏阳,江玲,赵志刚,万建民.利用高世代回交群体检测水稻垩白相关性状QTL.南京农业大学学报,2016,39(2): 183-190. Wang L S,Chen L M,Wang P R,Wang Z R,Zheng H, Ma H Y,Jiang L,Zhao Z G,Wan J M.Detecting the QTLofricechalkinesstraitsusingadvanced backcrossing population.J Nanjing Agric Univ,2016, 39(2):183-190.(in Chinese with English abstract)
[24]朱昌兰.稻低直链淀粉含量的遗传及品质形成对高温耐性的QTL分析.南京:南京农业大学,2004. Zhu C L.Identifying QTLs for thermo-tolerance of quality formation and inheritance of low amylose content in rice.Nanjing:Nanjing Agriculture University,2004. (in Chinese with English abstract)
[25]李健陵,林育炯,张晓艳,杜尧东,王华,吴丽姬,胡飞.抽穗期和乳熟期高温对水稻剑叶理化特性以及产量和品质的影响.农业现代化研究,2013,34(11): 109-113. Li J L,Lin Y J,Zhang X Y,Du Y D,Wang H,Wu L J, Hu F.Effects of high temperature on physiological and biochemical characteristics of flag leaves,grain yield and quality of rice in heading and milk stage.Res Agric Mod, 2013,34(11):109-113.(in Chinese with English abstract)
[26]张桂莲,陈立云,张顺堂,黄明,唐文邦,雷东阳,李梅华,贺治洲.高温胁迫对水稻花器官和产量构成要素及稻米品质的影响.湖南农业大学学报,2007,33(2): 132-136. Zhang G L,Chen L Y,Zhang S T,Huang M,Tang W B, Lei D Y,Li M H,He Z Z.Effects of high temperature stress on rice flower organ and yield components and grain quality.J Hunan Agric Univ,2007,33(2):132-136. (in Chinese with English abstract)
[27]张国发,王绍华,尤娟,王强盛,丁艳锋,吉志军.结实期不同时段高温对稻米品质的影响.作物学报,2006, 32(2):283-287. Zhang G F,Wang S H,You J,Wang Q S,Ding Y F,Ji Z J.Effect of higher temperature in different filling stages on rice qualities.Acta Agron Sin,2006,32(2):283-287. (in Chinese with English abstract)
[28]盛婧,陶红娟,陈留根.灌浆结实期不同时段温度对水稻结实与稻米品质的影响.中国水稻科学,2007,21(4): 396-402.Sheng Q,Tao H J,Chen L G.Response of seed-setting and grain quality of rice to temperature at different time during grain filling period.Chin J Rice Sci,2007,21(4): 396-402.(in Chinese with English abstract)
[29]赵飞,尹维娜,曲丽君,东丽,华泽田.水稻外观品质与产量构成因素的QTL解析.核农学报,2014,28(6): 990-997.Zhao F,Yin W N,Qu L J,Dong L,Hua Z T.Correlation and QTL analysis of the rice appearance quality and processing quality.J Nuclear Agric Sci,2014,28(6): 990-997.(in Chinese with English abstract)
[30]陶亚军,徐梦彬,王飞,陈达,周勇,梁国华.利用染色体单片段代换系定位水稻垩白QTL.华北农学报, 2015,30(1):1-8.Tao Y J,Xu M B,Wang F,Chen D,Zhou Y,Liang G H. QTL mapping of chalkiness using chromosome single segment substituted lines in rice.Acta Agric Boreali-sin, 2015,30(1):1-8.(in Chinese with English abstract)
[31]盘毅,罗丽华,邓化冰,张桂莲,唐文邦,陈立云,肖应辉.水稻开花期高温胁迫下的花粉育性QTL定位.中国水稻科学,2011,25(1):99-102. Pan Y,Luo L H,Deng H B,Zhang G L,Tang W B,Chen L Y,Xiao Y H.Quantitative trait loci associated with pollen fertility under high temperature stress at flowering stage in rice.Chin J Rice Sci,2011,25(1):99-102.(in Chinese with English abstract)
[32]Yamakawa H,Ebitani T,Terao T.Comparison between locations of QTLs for grain chalkiness and genes responsive to high temperature during grain filling on the ricechromosomemap.BreedingSci,2008,58(3): 337-343.
[33]高方远,邱玲,陆贤军,任鄄胜,吴贤婷,任光俊,曾礼华.杂交籼稻骨干保持系岗46B稻谷粒形及垩白的QTL分析.中国水稻科学,2014,28(3):235-242. Gao F Y,Qiu L,Lou XJ,Ren Z S,Wu X T,Ren G J, Zeng L H.QTL analysis on grain shape and chalkiness of an elite maintainer line Gang 46B in hybrid rice(Oryza sativa L.).Chin J Rice Sci,2014,28(3):235-242.(in Chinese with English abstract)
Identifying QTLs for Thermo-tolerance of Grain Chalkiness Trait in Rice
ZHANG Guilian,LIAO Bin,TANG Wenbang,CHEN Liyun*,XIAO Yinghui*
(College of Agronomy,Hunan Agricultural University,Changsha 410128,China;*Corresponding author,E-mail:chenliyun996@163.com, xiaoyinghui@163.com)
【Objective】The molecular markers linked to high temperature tolerance of rice appearance quality were screened for the purpose of providing an important foundation of improvement for rice grain quality.【Method】The quantitative trait loci(QTLs)for heat tolerance of grain chalkiness trait were identified using the recombinant inbred lines(RILs)derived from a cross between the heat tolerant rice line 996 and the heat sensitive line 4628.The heat tolerance indexes of chalky grain rate,chalkiness size and chalkiness degree were used as the heat-tolerance indicators.【Result】24 QTLs for heat tolerance of grain chalky traits were detected in two years by using composite interval mapping analysis including eight for heat tolerance of chalky grain rate(HTCGR),twelve for heat tolerance of chalkiness size and fourteen for heat tolerance of chalkiness degree(HTCD).Two HTCGR QTLs on chromosome 6 and two HTCD QTLs on chromosome 7 could be detected repeatedly in two years.The two HTCD sites coincided with two HTCGR sites on chromosome 7 detected in 2015.Moreover,anyone of the four pleiotropic QTLs had impacts on the chalky grain rate,chalkiness size and chalkiness degree.【Conclusion】The QTL controlling the heat tolerance index of chalkiness degree qHTCGR6.1 and the QTL controlling the heat tolerance of chalkiness degree qHTCD7.1 might be new findings..
rice;chalkiness trait;thermo-tolerance;quantitative trait locus
Q948.112+2;Q343.1+5
:A
:1001-7216(2017)03-0257-08
2016-12-25;修改稿收到日期:2017-02-28。
国家自然科学基金资助项目(30900874);湖南省自然科学基金资助项目(11JJ3026);湖南省科技计划博士后专项(2012RS4037);湖南省教育厅重点项目(17A100);作物学开放基金资助项目(ZWKF201504)。
