日本の寒地,北海道の稲作限界地帯におけるもち米の精米蛋白質含有率および米粒白度の年次間地域間差異とその発生要因 (日语原文)
2022-09-27丹野
丹野 久
(日本水稲品質・食味研究会,日本 東京都中央区,104-0033)
キーワード:精米蛋白質含有率;米粒白度;年次間地域間差異;もち米;稲作限界地帯
北海道における糯品種の作付け地域は,粳品種からの飛散花粉との交雑により胚乳が粳性となるキセニア粒の発生や粳粒の機械的混入により品質を低下させないために,粳品種の作付け地域から離れた地域で,もち米生産団地を形成している[1]。これらの地域は,北海道の稲作地帯の中でも気象条件が厳しい稲作の限界地帯である(図 1)。そのため,北海道のもち米の作柄は気象条件の影響を受けやすく,年次間差異や地域間差異が生じやすい(図 2)。もち米は加工原料となることが多く,品質はその利用上重要である。もち米品質の安定化を図るためには,もち米品質の年次間と地域間の差異およびその発生要因を明らかにすることが必要である。

図1 日本北海道の主要なもち米産地およびうるち米産地における水稲栽培期の平均気温の推移
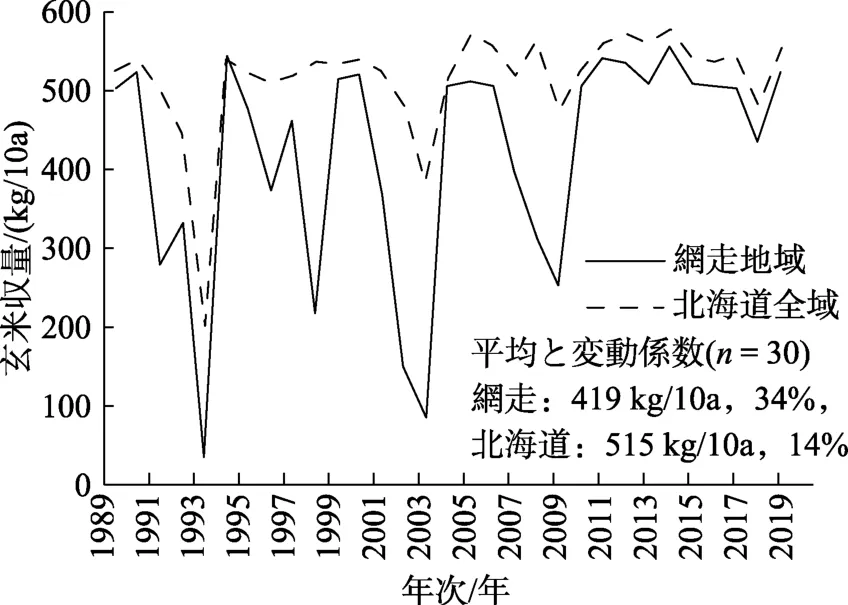
図2 稲作限界地帯の糯品種作付け地域である網走(オホーツク)地域および北海道全域における玄米収量の年次推移[1]
そこで,北海道のもち米の作柄が大きく異なった2000—2003年の4カ年に生産された糯品種「はくちょうもち」[2]の玄米サンプルを,糯品種作付けの主要な地域から収集した。例えば,水稲作付けのほとんどが糯品種である網走地域において,これら年次の玄米収量は,86~520 kg/10a,平均281 kg/10a,変動係数71%で,最小値と最大値の差異が434 kg/10aと大きかった。これは網走地域で同品種が多く作付けされた 1991—2010年の 20カ年の玄米収量 35~546 kg/10a,平均368 kg/10a,変動係数42%と比べ,最小値最大値の差異の比で0.85倍とやや小さいにすぎず,変動係数が1.7倍と大きかった[1]。すなわち,本収集サンプルを分析調査することにより,北海道もち米品質の年次間と地域間の差異を明らかにできると考えられる。
以上のことから,これらのサンプルを用いて,もち米品質である精米蛋白質含有率(以下,蛋白質と記す),米粒白度,検査等級に関わる米粒外観品質,および搗き餅の硬化性とそれに関係する糊化特性等を分析した。その分析データを,糯品種作付けの主要な地域である 6地域に分け,年次間差異と地域間差異およびそれらと気象や水稲生育特性との間の関係を調査した[3]。
例えば,もち米品質の中で,蛋白質は搗き餅の食味[4]および餅生地の伸展性[5]や膨化伸長性[6]に影響し,低いことが要望される。また,精米白度はもち米を原料とする製造品の外観に影響するため,高いことが重要である[7]。しかし,北海道のもち米は東北以南のもち米に比べ,従来,蛋白質が高く精米白度も劣るとされる[7-8](図3,図4)。そこで,本報では,まず蛋白質および米粒白度についての年次間と地域間の差異およびその発生要因を解明し,さらに目標とする米粒白度を得るための施肥量を算出する方法を示した[9]。

図3 北海道もち米における精米蛋白質含有率の年次推移と東北以南産との比較[8]

図4 北海道もち米における精米白度の年次推移と東北以南産との比較[8]
1 栽培期と生育期別の気温および生育特性の年次間地域間の差異
栽培期の栄養成長期,すなわち出穂期前にほぼ当たる5-7月積算気温,出穂開花期から登熟期にほぼ当たる8-9月の積算気温,および出穂前24日以降30日間(以下,障害型冷害危険期と記す)の平均気温と出穂後40日間の日平均積算気温(以下,登熟気温と記す)において,最小値と最大値の差異および変動係数は,年次間を地域間に比べと,2.5~8.5倍と年次間が大きかった。さらに生育特性おいても同様に年次間を地域間に比べると,出穂期は0.9,1.1倍とほぼ同じであるが,この出穂期を除いた不稔歩合,千粒重および玄米収量では,1.3~1.8倍と年次間が同様に大きかった(表1)。

表1 北海道もち米の試験年次別と地域別における栽培期間の日平均積算気温,生育期別気温および生育特性[3,10]
2 精米蛋白質含有率と米粒白度の年次間地域間差異
蛋白質における最小値と最大値の差異および変動係数は,年次間と地域間でそれぞれ,同差異1.5ポイント(最小値8.4%~最大値9.9%,以下同じ)と変動係数 8.9%,1.5ポイント(8.4%~9.9%)と 7.1%であった。同様に,玄米白度が 4.9(21.6~26.5)と 9.2,2.1(22.3~24.4)と 3.6,精米白度が 8.0(45.1~53.1)と 7.7,5.0(45.2~50.2)と 4.3であった。これらの最小値最大値の差異と変動係数は年次間を地域間に比べると,蛋白質がそれぞれ1.0,1.3倍と同じか年次間がやや大きく,玄米白度は2.3,2.6倍,精米白度は1.6,1.8倍と年次間が地域間よりも大きかった(表1)。
以上のことから,栽培期の気温は年次間差異が地域間差異よりも大きく,そのため水稲生育期の気温も同様に年次間差異が大きくなった。その影響から,玄米収量を含む水稲生育の特性でも出穂期を除いて年次間差異が大きかった。そのため,蛋白質は年次間差異が地域間差異と同じかやや大きく,玄米白度と精米白度は年次間が大きかった。
3 栽培期と生育期別の気温や生育特性での年次と地域の各平均値と変動係数との間の関係
年次と地域での各平均値と年次では各年次に供試した6地域間の変動係数,地域では各地域の 4年次間の変動係数との間の関係をみると,5-7月および8-9月の積算気温では5-7月の年次間のみで負の相関関係にあり,同積算気温が低い年ほど地域間の変動が大きくなった。また,出穂期と登熟気温では年次間と地域間とも一定の関係が見られなかったが,障害型冷害危険期の平均気温では年次間と地域間ともに負,不稔歩合では正,千粒重および玄米収量ではいずれも負の相関関係が見られた(表2,図5)。

表2 北海道もち米の栽培期間と生育期別の気温および生育特性における試験年次と地域の各平均値と変動係数との間の相関係数[10]
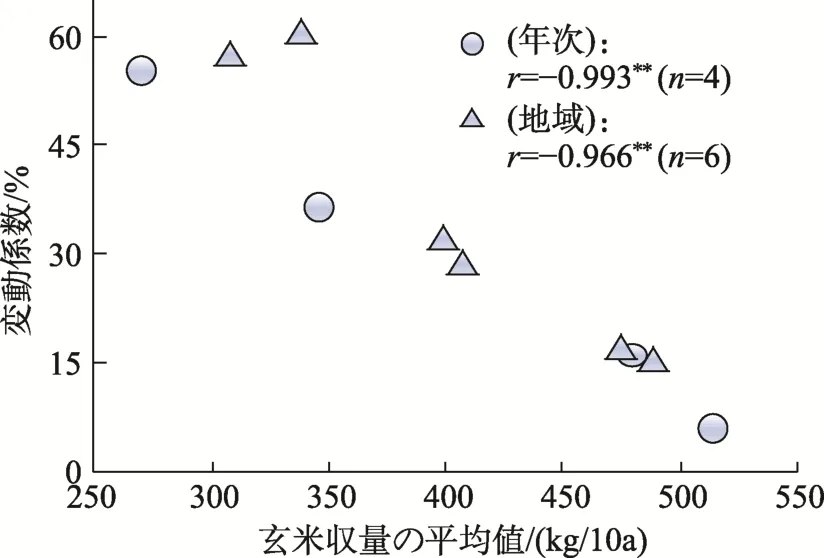
図5 玄米収量における平均値と変動係数との間の関係[10]
蛋白質の平均値と変動係数との関係では,年次と地域ともに,平均値が高いほど変動係数が大きかった(表3,図6,図7)。ただし,年次地域込みでは蛋白質が 10.5%をこえると,変動係数が小さくなる傾向があった。すなわち,地域全体が高蛋白化することで均一化していると推察された。また,玄米白度と精米白度でも年次と地域ともに平均値が低いほど変動係数が大きかった(表3,図8,図9)。

表3 北海道もち米の精米蛋白質含有率と米粒白度における試験年次と地域の各平均値と変動係数との間の相関係数[10]

図6 精米蛋白質含有率の年次別頻度分布

図7 精米蛋白質含有率における平均値と変動係数との間の関係[10]

図8 精米白度の年次別頻度分布
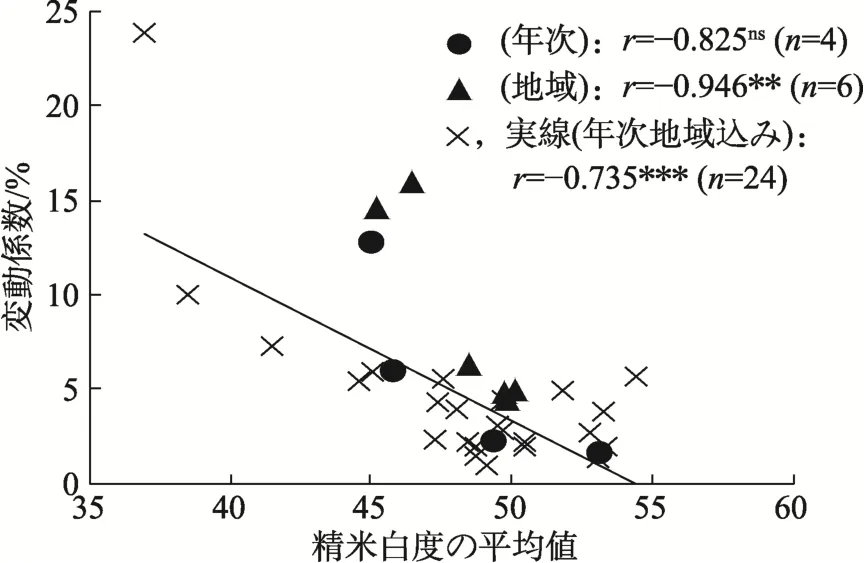
図9 精米白度における平均値と変動係数との間の関係[10]
以上のように,玄米収量および蛋白質,米粒の両白度が不良な年次や地域ほどそれぞれ地域間および年次間の変動係数が大きくなった。これらのことは,一定品質の米を安定的に生産する上で困難な要因であると推察された。
4 精米蛋白質含有率,米粒白度,生育期別気温および生育特性との間の関係
年次間では,出穂期が早く,障害型冷害危険期の平均気温が高く不稔歩合が低く,登熟期の日照時間(以下,登熟日照と記す)が長く千粒重が重く多収なほど,蛋白質が低くなった(表4,図10~12)。一方,地域間では蛋白質と不稔歩合,玄米収量および千粒重との間に年次間と同様な関係が認められたが,他の形質との関係は不明確であった。なお,蛋白質は年次と地域込みで,登熟気温が 849 ℃で最低になりそれより低くあるいは高くなるにともない高くなった(図13)。
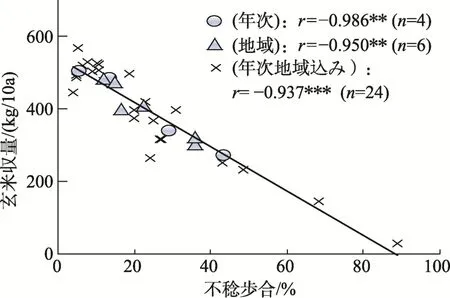
図10 不稔歩合と玄米収量との間の関係[10]
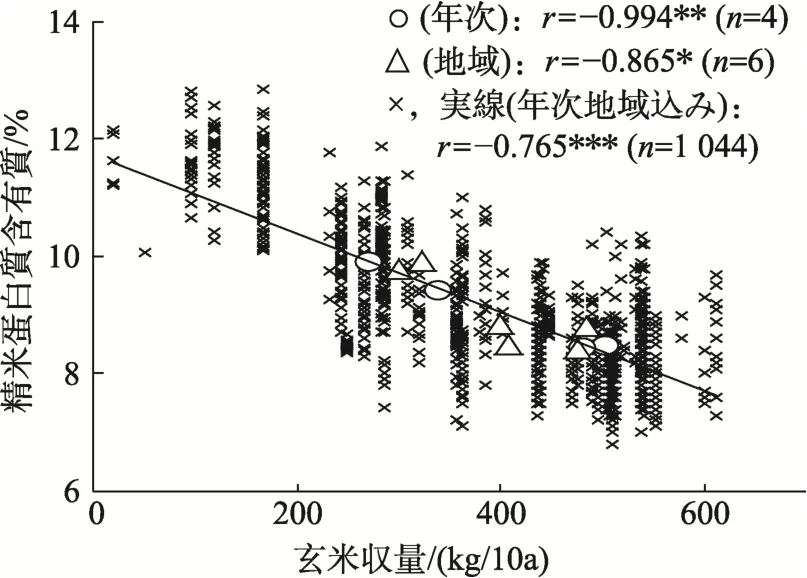
図11 玄米収量と精米蛋白質含有率との間の関係[10]

図12 不稔歩合と精米蛋白質含有率との間の関係[10]

図13 出穂後40日間の日平均積算気温と精米蛋白質含有率との間の関係[10]
玄米白度と精米白度との間には年次間,地域間でいずれも明確な正の相関関係があった(図14)。また,年次間では,障害型冷害危険期および登熟気温が高いほど,不稔歩合が低く千粒重が重く多収となり,蛋白質が低く玄米白度と精米白度が高かった(表4,図15~17)。一方,地域間では,米粒の両白度は,不稔歩合が低く千粒重が重く多収で低蛋白になるほど高くなったが,登熟気温および登熟日照との間には明確な関係がなかった。

表4 北海道もち米の年次間と地域間における精米蛋白質含有率,米粒白度,生育特性および生育期別気温の間の相関係数[10]
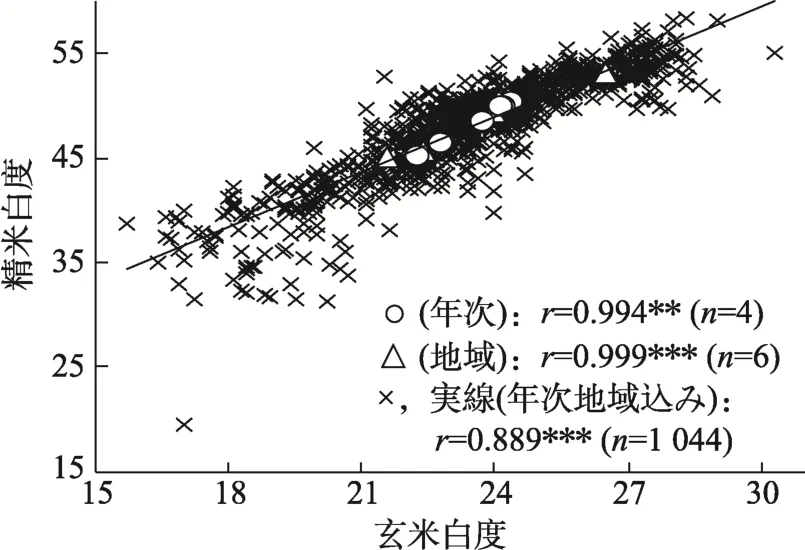
図14 玄米白度と精米白度との間の関係

図15 出穂後40日間の日平均積算気温と玄米白度との間の関係

図16 玄米収量と精米白度との間の関係[10]

图17 大米蛋白质含量与大米白度之间的关系[10]
すなわち,前述の項目1と項目2から,生育気温や生育特性の間では最小値最大値の差異や変動係数が地域間に比べ年次間で大きかった。そのため,それらの間の相関関係も年次間で地域間より明確であった。また,これらの相関関係から,前述の項目3における年次と地域の各平均値と変動係数との間の関係では,5—7月の気温低下が障害型冷害危険期の気温低下を生じさせ,不稔を発生させるとともに籾殻形成の抑制を通して千粒重を軽くさせ[11],玄米収量を低下させ,蛋白質を上昇させて米粒の両白度を低下させたことが,年次と地域のいずれにおいても,それらが不良なほど変動係数を大きくした要因となっていると推察した。
5 低蛋白米生産技術および目標の米粒白度に基づいた施肥量の決定
生産米の良質化のため,蛋白質を下げるとともに米粒の両白度を高めるには,蛋白質と米粒の両白度との間には負の相関関係が認められることから(表4,図17),栽培に当たり蛋白質を低下させることが重要である。そのため,うるち米の良食味化を目的に開発された低蛋白米生産技術を励行する必要がある[12]。
さらに,玄米白度と精米白度は1 m2の1籾当たりの登熟気温(以下,1籾当たりの登熟気温と記す)といずれも正の相関関係が見られた(図18,図19)。ここで,玄米白度の目標値を,既報[7]で目標値に設定された精米白度50に相当する25(図14)とすると,その一次回帰式から,1籾当たりの登熟気温は0.025 ℃/粒/m2以上が必要であった。同0.025℃/粒/m2は,平年気象の登熟気温を主要なもち米産地の1つである名寄市を例にして 760 ℃とすると,m2当たり籾数は30,000であった(図20)。また,m2当たり籾数30,000に対応する成熟期の窒素吸収量は 10 kg/10aであった(図21)。さらに,土壌別の施肥量と成熟期窒素吸収量および玄米収量との間の関係を明らかし,また目標収量を考慮し,目標の玄米白度を得るための土壌別での施肥量が決定できた[9]。

図18 1 m2の1籾当たり出穂後40日間の日平均積算気温(1籾当たり登熟気温)と玄米白度との間の関係[9]

図19 1 m2の1籾当たり出穂後40日間の日平均積算気温(1籾当たり登熟気温)と精米白度との間の関係[9]

図20 1 m2の1籾当たり出穂後40日間の日平均積算気温(1籾当たり登熟気温)とm2当たり籾数および登熟気温との間の関係[9]

図21 成熟期窒素吸収量とm2当たり籾数との間の関係[9]
