仿刺参体壁再生过程中两个核糖体蛋白基因的表达差异
2017-05-13秦艳杰王文文王艳枫李霞
秦艳杰,王文文,王艳枫,李霞
(1.大连海洋大学辽宁省海洋生物资源与生境修复重点实验室,辽宁大连116023;2.大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023)
仿刺参体壁再生过程中两个核糖体蛋白基因的表达差异
秦艳杰1,2,王文文1,王艳枫2,李霞1,2
(1.大连海洋大学辽宁省海洋生物资源与生境修复重点实验室,辽宁大连116023;2.大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023)
本研究首次从仿刺参(Apostichopus japonicus)体壁中克隆得到核糖体蛋白L30(ribosomal protein L30,RPL30)的cDNA全长序列(GenBank:JQ770165),该序列包括56 bp的5′-UTR,162 bp的3′-UTR和339b p的开放阅读框,共编码112个氨基酸;Blast比对分析结果显示该序列的核苷酸序列以及推导的氨基酸序列与其他物种的同源性均在75%以上;RPL30基因在仿刺参各组织中均有表达,其中肠、体腔细胞、纵肌、体壁中表达量分别为呼吸树的7.24倍(P<0.01)、4.47倍(P<0.01)、3.12倍(P<0.05)、1.35倍(P>0.05);仿刺参体壁再生不同阶段核糖体蛋白基因RPL30和RPL17的表达存在差异,体壁再生第7 d时RPL30基因的表达出现峰值,为对照组的2.13倍(P>0.05),其余天数的相对表达量均低于对照组,其中第1、5、6d的表达量与对照组差异均显著(P<0.05);RPL17基因在再生第2、5d分别出现峰值,分别为对照组的7.47倍和5.60倍(P< 0.05),其余各天的表达量与对照组差异不显著(P>0.05)。研究结果表明核糖体蛋白基因除构成核糖体参与蛋白质合成外,还有着各自的核糖体外功能。本研究结果将为进一步研究仿刺参蛋白质合成、再生及多种生理活动的调控机制奠定基础。
仿刺参;核糖体蛋白L30;基因克隆;核糖体蛋白L17;再生
仿刺参又称刺参,属棘皮动物门(Echinodermata),海参纲(Holothuroidea),楯手目(Aspidochirotida),刺参科(Stichopodidae),是我国北方沿海重要的海水养殖经济种类,其营养价值和经济价值均较高(姜健等,2004;赵鹏等,2013)。除此之外,仿刺参较强的再生能力也是众多学者关注的热点(聂竹兰等,2006),国内外学者对其再生的组织学和细胞学过程进行了大量研究(聂竹兰等,2006;Garcia-arraras et al,2001;李霞等,2007),成果显著,而对再生的分子生物学研究相对较少,主要集中在对再生相关基因的挖掘上(Santiago et al,2000;Zheng et al,2006;陈秋实等,2008;秦艳杰等,2013;赵祥吉等,2013)。
一般认为核糖体蛋白主要负责组成核糖体,参与蛋白质的合成。近年来也有研究表面核糖体蛋白还参与细胞的分裂、增殖、分化、凋亡、发育调控等过程(侯怡铃等,2010;Vilardell et al,2010)。真核生物约有80种核糖体蛋白,分别组成核糖体的大亚基(RPL)和小亚基(RPS)。核糖体蛋白L17(ribosomal protein L17,RPL17)和L30(ribosomal protein L30,RPL30)都是大亚基的组成部分,位于细胞质中。RPL17和核糖体蛋白L22家族,是核糖体蛋白大亚家族的核心成员。RPL30属于核糖体蛋白L30e家族成员,是实现核糖体的装配以及保证核糖体功能作用发挥的关键成分,它不仅能独立地与RNA相互作用,又能与自身的前体mRNA 5′端前导序列结合调节其蛋白质的合成(王艳枫等,2013)。GenBank数据库显示很多水产动物中已克隆出该基因,但尚未见仿刺参RPL30基因的报道。关于RPL30、RPL17基因功能的研究报道较少,有学者利用消减差异技术发现RPL30基因在肝癌组织中过表达(Kondoh et al,2001),并认为生长相关蛋白GABP可能是RPL30基因的正调节物(Lü et al,2007)。本研究以前期仿刺参体壁再生差减文库构建过程中筛选到的一个高表达的EST序列(秦艳杰等,2013)为中间序列,采用SMART-RACE法,克隆获得仿刺参的RPL30基因的全长cDNA序列,并利用Real-time PCR技术检测了RPL30基因在仿刺参各组织中的表达差异,分析了RPL30和RPL17两个核糖体基因在仿刺参再生过程中的表达模式。旨在为进一步研究核糖体蛋白基因在仿刺参组织发育、再生修复及其他生理活动中的调控机制提供资料。
1 材料与方法
1.1实验材料
试验用刺参取自旅顺龙王塘海区,为1-2龄个体,麻醉状态下体长为10~14 cm。室温暂养于大连海洋大学重点开放实验室,每天半量换水1次,投喂人工配制饵料。
不同组织的取材:随机挑选5头生长状况良好的刺参,分别采集它们的体壁、肠、呼吸树、纵肌、体腔细胞(体腔液3 000 rpm/min离心5 min后收集体腔细胞),每种组织混合于1个冻存管中,液氮保存。再生不同阶段的取材:取正常健康的刺参45头,冰上麻醉15~30 min,于其背部无脊处切取体壁组织约100 μg,形成人工创伤,创伤后正常饲养。实验组:分别取创伤后1~8 d再生出的体壁组织,每个时间段各取3头刺参;对照组:正常即无创伤的体壁组织。将取得的体壁组织置于冻存管迅速投入液氮中保存。
1.2总RNA提取及纯化
将液氮冻存的正常仿刺参各组织及再生不同阶段的体壁组织,采用TRIZOL一步法分别提取总RNA,获得的总RNA经RNase-Free DNase和RNasin Plus RNase Inhibitor处理,再经酚氯仿抽提、糖原和醋酸钾溶液沉淀、乙醇洗涤、0.1% DEPC处理水定容获得纯化的RNA,-80℃保存备用。用1.0%琼脂糖凝胶电泳检测总RNA完整性,以28S和18S组分的比率为参考,判断RNA样品的降解程度;核酸蛋白检测仪检测总RNA的浓度和纯度。
1.3引物设计
利用Primer Premier 5.0软件设计RPL30基因RACE引物:L303S、L305A1、L305A2和Realtime PCR特异性引物:L30F和L30R,内参引物为Cytb F和Cytb R。实验中所用到引物均由上海生物工程有限公司合成,引物序列见表1。

表1 实验用引物序列
1.4 RPL30的cDNA序列克隆
1.4.1 中间序列的获得
本实验室先期构建了仿刺参体壁再生消减cDNA文库,发现一段长为253bp的EST序列与核糖体蛋白基因RPL30具有较高同源性。以此序列为中间序列设计引物进行cDNA全长克隆。
1.4.2 3′RACE扩增
3′RACE采用SMARTTM RACE cDNA Amplification Kit(Clontech,美国)试剂盒进行。3′RACE扩增体系(25μL):ddH2O,13.5μL;10×PCRBuffer,2.5 μL;dNTP,2.0 μL;Mg2+,2.0 μL;3'RACEReady cDNA,1.25 μL;UPM,2.5 μL;L303S,1.0 μL;rTaq酶,0.25μL。3′RACE扩增程序:94℃,5 min;94℃,30 s;60℃,30 s;72℃,2 min;共35个循环,72℃,10 min。
1.4.3 5′RACE扩增
5′RACE扩增也采用SMARTTMRACE cDNA Amplification Kit(Clontech,美国)试剂盒进行。5′RACE1stPCR反应体系(25μL):灭菌水,13.75 μL;10×PCRBuffer,2.5μL;dNTP,2.0μL;Mg2+,2.0μL;5'RACE–Ready cDNA,1.0 μL;UPM,2.5 μL;L305A1,1.0 μL;rTaq酶,0.25 μL。5′RACE 1st PCR反应程序:94℃,5 min;94℃,30 s,72℃,2 min,5个循环;94℃,30 s,68℃,30 s,72℃,2 min,5个循环;94℃,30 s,66℃,30 s,72℃,2 min,5个循环;94℃,30 s,64℃,30 s,72℃,2 min,5个循环;94℃,30 s,62℃,30 s,72℃,2 min,25个循环;72℃,10 min。
5′RACE 2nd PCR反应体系(25 μL):ddH2O,13.75 μL;10×PCR Buffer,2.5 μL;dNTP,2.0 μL;Mg2+,2.0 μL;模板(稀释100倍的1st PCR产物),1.0 μL;NUP,2.5 μL;L305A2,1.0 μL;rTaq酶,0.25μL。5′RACE2ndPCR反应程序:94℃,5 min;94℃,30 s;60℃,30 s;72℃,2 min;共35个循环,72℃,10 min。
1.4.4 PCR产物的克隆测序
分别取5 ul 3′RACE/5′RACE PCR最终产物进行1.0%琼脂糖凝胶电泳检测,将获得的PCR产物回收并与PMD19-T载体连接,转入E.coli DH5α感受态细胞进行蓝白斑筛选,菌落PCR检测获得的阳性克隆送至上海生工生物工程有限公司进行测序。
1.5生物信息学分析
在NCBI数据库中利用BLAST程序将获得的仿刺参RPL30序列与其他物种进行同源性比对和相似性搜索;ORF Finder软件确定正确的开放阅读框,并翻译成氨基酸序列;使用Expasy网站中的Proparam功能(http://web.expasy.org/protparam/)进行氨基酸序列分析;运用SOPMA在线工具(http://npsa-pbil.ibcp.fr/cgi-bin/secpred_sopma.pl)进行二级结构分析;用Predict Protein在线工具(https://www.predictprotein.org/)进行蛋白功能位点预测;利用Expasy网站中ProtScale在线工具(http://web.expasy.org/protscale/)进行亲水性/疏水性分析;利用ClustalX2.0软件和MEGA4.0软件采用邻位相连法(Neighbor-Joining,NJ)在Bootstrap置信值为1 000的条件下构建系统进化树。
1.6 Real-timePCR定量分析
1.6.1 cDNA序列的合成
分别取10 μL仿刺参体壁、肠、呼吸树、纵肌、体腔细胞5种组织及体壁再生8个时间段的总RNA,按照反应体系:PCR Bμffer(5×)4μL,随机引物2μL,RT Enzyme 1μL,总RNA 10μL,灭菌水3μL,反应程序:42℃60 min,85℃10 min,反转录获得cDNA。
1.6.2 Real-time PCR反应
根据已获得的RPL30全长序列和已报道的RPL17全长序列(GenBank NO:JQ922561),设计合成Real-time PCR特异性引物L30F/L30R和L17F/L17R(表1)和内参引物Cytb F/Cytb R(表1)。荧光定量PCR反应体系:GreenqPCRMix(2×)10μL,Forward/ReversePrimer(10μM)各0.4μL,ROX0.4μL,cDNA模板1.0 μL,灭菌水定容至20 μL。反应条件:95℃10 min;95℃15 s,60/55℃60 s,40个循环;之后进行融解曲线绘制,反应条件:95℃ 15 s,60℃30 s,95℃15 s。荧光定量反应在StepOnePLUS(ABI,USA)PCR仪上进行,每个反应重复3次,2-△△CT法分析目的基因的相对表达量。
2 结果与分析
2.1序列分析
最终获得长度为557bp的cDNA序列(图1),已提交到GenBank,登录号为JQ770165。该序列包括56bp的5′-UTR,162bp的3′-UTR,开放阅读框339bp,起始密码子为ATG,编码氨基酸为甲硫氨酸,终止密码子为TAG,共编码112个氨基酸(图1),预测该蛋白相对分子质量为12.469 5 Kd,等电点为9.57,总平均亲水性为-0.326,定位于细胞质中(RI=1;Expected Accurcy=56%)。除起始密码子编码的甲硫氨酸外的111个氨基酸中,有9个带负电荷的氨基酸(天冬氨酸D和谷氨酸E),17个带正电荷的氨基酸(精氨酸R和赖氨酸K),其余氨基酸为非极性、疏水性氨基酸和极性、中性氨基酸。疏水性分析结果表明该蛋白为不稳定的亲水性蛋白。二级结构预测,结果显示37个氨基酸属于α螺旋,占33.04%,35个氨基酸属于延伸链,占31.25%,17个氨基酸属于β折叠,占15.18%,23个氨基酸属于无规卷曲,占20.54%。α螺旋和延伸链这两类结构元件在RPL30中所占比例较大,而β折叠和无规则卷曲则散布于整个蛋白质中。对核糖体蛋白RPL30的功能位点进行预测,发现刺参RPL30共有6种功能位点,包括:蛋白激酶C磷酸化位点(SKK、SGK、TLK、SLR)4个;酪蛋白激酶II磷酸化位点(SEIE)1个;酪氨酸激酶磷酸化位点(RKSEIEYY)1个;N-豆蔻酰化位点(GIKQTL、GTACGK)2个;核糖体蛋白L30e信号1位点(SGKYILGIKQTLKSLRQGKAKLIIL)1个;核糖体蛋白L30e信号2位点(ELGTACGKYFRVCTLCVTDPG)1个。

图1 仿刺参RPL30 cDNA序列和氨基酸序列分析
2.2同源性比对与系统进化树的构建
将序列在NCBI上进行Blastn比对,发现该序列与其他生物的RPL30基因相似性在75%以上,其中与紫海胆(Strongylocentrotus Purpuratus,XM_778057)的相似性最高为80%,E值为6e-79;与文昌鱼(Branchiostoma belcheri,AF420432)的相似性为79%,E值为6e-66;与黄岛长吻虫(Saccoglossus kowalevskii,XM_002732441)的相似性为78%,E值为1e-67。
Blastx比对分析结果显示该序列与其他生物的RPL30相似性在80%以上,其中与紫海胆(S. purpuratus,XP_783150)的相似性最高为88%,E值为7e-67;与沙蚕(Eurythoe complanata, ABW23229)相似性为83%,E值为4e-61;与非洲爪蟾蜍(Xenopus laevis,NP_001080621)的相似性为83%,E值为4e-61;与斑马鱼(Danio rerio,NP_956322)的相似性为81%,E值为3e-59;与家鼠(Mus musculus,AAI00609)的相似性为80%,E值为8e-60。
利用NCBI网站的BlastX功能在GenBank中搜寻和筛选到19个物种(图2)的RPL30的氨基酸序,应用软件ClustalX2.0、MEGA4.0,采用邻位相连法(Neighbor-Joining,NJ)构建系统进化树,Bootstrap置信值为1 000。结果显示仿刺参与同为棘皮动物的紫海胆聚为一支,亲缘关系最近,尾索动物海鞘、半索动物黄岛长吻虫的聚类顺序与仿刺参也较近。脊椎动物单独聚为一支,其中鱼类和哺乳动物分别各自聚为一个分支。
2.3 RPL30基因在仿刺参不同组织中的表达
RPL30基因在仿刺参5种组织中的表达情况如图3所示,各组织表达量在显著性水平为0.05时表现为肠>体腔>纵肌>体壁≈呼吸树,其中肠、体腔细胞中的表达量分别为呼吸树的7.24、4.47倍,与呼吸树的差异均极显著(P<0.01),肠与体腔细胞中的表达量差异极显著(P<0.01);纵肌中表达量为呼吸树的3.12倍,差异显著(P<0.05);体壁为呼吸树的1.35倍,差异不显著(P>0.05)。
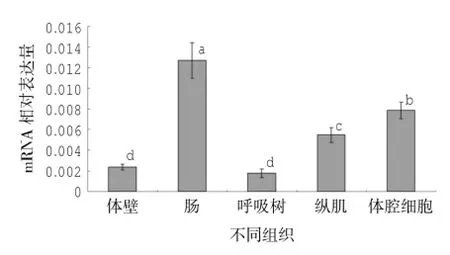
图3 仿刺参各组织中RPL30基因的相对表达量(以Cytb基因为内参,标注不同字母代表差异显著)
2.4 RPL30基因在刺参体壁再生不同阶段的表达
RPL30基因在仿刺参再生不同阶段的表达量变化如图4所示,结果显示:仿刺参体壁再生8d过程中RPL30基因的表达量呈现先下降再升高的趋势,其中仅在第7 d高于对照组,为对照组的2.13倍,差异显著(P<0.05);其余天数的相对表达量均低于对照组,其中第1、5、6 d的表达量分别为对照组的0.32、0.26和0.16倍,差异均显著(P< 0.05),第2、3、4、8 d的相对表达量为对照组的0.53、0.42、0.54和0.39倍,差异不显著(P>0.05)。
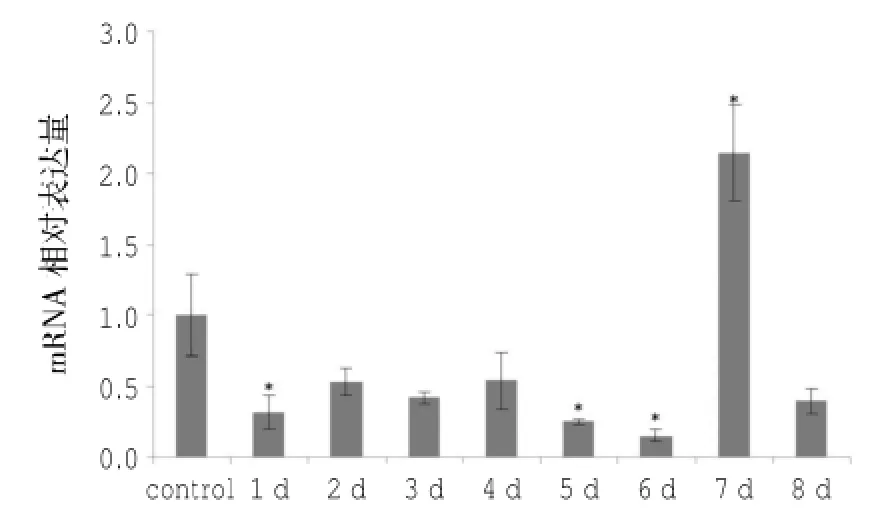
图4 仿刺参体壁再生不同时期RPL30基因的相对表达量
2.5 RPL17基因在刺参体壁再生不同阶段的表达
正常和创伤后1-8 d的体壁组织中RPL17基因的表达变化情况如图5,结果显示:创伤8 d过程中RPL17基因的表达量在第2和第5 d时显著高于对照组,分别为对照组的7.47倍(P<0.01)和5.60倍(P<0.05),且第2 d和第5 d的差异不显著(P>0.05);其余第1、3、4、6、7、8 d的表达量与对照组之间不存在显著差异(P>0.05),可见该基因在刺参体壁创伤后第2 d和第5 d分别出现表达高峰。

图5 刺参体壁再生不同时期RPL17基因的相对表达量(以Cytb为参照)

图6 台湾岛西南反气旋涡随时间演变过程
3 讨论
本研究通过SMART-RACE法,首次克隆得到仿刺参核糖体蛋白RPL30基因的全长cDNA序列。对已经克隆得到的不同物种的RPL30的基因序列及其编码的氨基酸序列进行BLAST分析,结果发现从无脊椎动物的软体动物鲍、环节动物沙蚕、棘皮动物海胆、尾索动物海鞘到脊椎动物鱼类斑马鱼、两栖类非洲爪蟾蜍、哺乳动物鼠和人等这样的进化跨度内该核糖体蛋白基因均保持有70%以上的同源性,由此可说明RPL30基因在生物进化中相对保守,这种高度的遗传稳定性对维持相关的功能具有很重要的作用。另外,对仿刺参、紫海胆、斑马鱼、非洲爪蟾蜍、人的RPL30二级结构进行比较,发现这些物种二级结构组成元件基本相同,只是每个组成元件所占比例不同;蛋白功能位点分析表明紫海胆和人的RPL30蛋白同样含有6种功能位点,推测此蛋白功能较为保守。系统进化树结果显示,以RPL30为分子进化标准得到的物种间亲缘关系与它们在生物学分类上的关系是一致的。
本研究中通过Real-time PCR分析了RPL30基因在仿刺参中的表达模式。结果显示RPL30基因在仿刺参各组织中都有表达,表明该基因对仿刺参组织生长发育起到一定的调控作用。其中,RPL30基因在肠和体腔中的表达量显著高于其他组织,推测这可能与仿刺参自身的组织结构特征相关。肠组织是仿刺参主要的消化道,含有消化酶在内的多种酶类,需进行大量的物质代谢和能量代谢(李霞等,2012),因此该组织的生理活动相对活跃,RPL30基因在肠组织中的表达量也相对较高。仿刺参体腔液中存在多种类型的体腔细胞,体腔细胞的功能类似于脊椎动物的血细胞,除参与物质运输外,也是仿刺参体内主要的免疫防线,体腔细胞会通过包裹、吞噬或释放具有杀菌作用的细胞因子的方式对抗外来物质,由于仿刺参缺乏特异性免疫组织和器官,其免疫系统主要是由细胞免疫和体液免疫组成的非特异性免疫系统(王淑娴等,2012),因此为了随时抵抗外源细菌的侵蚀以及完成自身的新陈代谢,体腔中该类细胞通常会处于增殖活跃期,因此RPL30基因在体腔细胞中表达量也较高;体壁、呼吸树、纵肌中两个基因的表达量相对较低,推测与本研究中所取的仿刺参为健康状态良好的成体,未经任何生理或人为因素刺激,因此这些组织的结构和功能相对稳定有关(王艳枫等,2013)。
本实验中RPL30基因在再生8 d时间内只有第7 d时基因的表达量才显著高于对照正常体壁中的表达量,前6 d的表达量均低于正常体壁组织中的表达量。推测是由于仿刺参在手术创伤前期出现了炎症反应,手术创伤后某些致炎因子作用于机体造成细胞的损坏,使细胞显现变性、坏死,因此前6 d为机体对抗炎症反应的阶段,此阶段损伤细胞的速度远大于细胞增殖的速度,表现为基因表达量较正常体壁的表达量出现下降。第7 d,炎症反应得到控制,细胞损坏速度下降,细胞增殖活动显现出来,因此此时基因的表达出现显著升高的趋势。随后第8 d表达量下降,分析出现下降的原因可能是再生过程是一个细胞分化、增殖、迁移的过程(聂竹兰等,2006;Garcia-arraras et al.2001;李霞等,2007),再生前期细胞需要大量增殖来启动组织的再生,积累到一定量后,RPL30基因可能会通过某些信号途径阻止细胞的增殖活动(Sara et al,2005;朱可可等,2009),细胞活动开始变为以迁移为主,因此再生后期基因的表达量出现下降。
同为组成核糖体大亚基的RPL17基因在仿刺参中已有克隆和组织表达的报道(王艳枫等,2013),本研究中发现RPL17基因在仿刺参体壁再生过程中的表达趋势为先上升后下降,再上升再下降,共出现两个峰值,再生前8 d的表达量较之对照组呈现高表达的趋势。推测出现第1个峰值是对抗创伤刺激的结果,手术创伤初期免疫防御系统被激活,核糖体也必须大量合成以保证细胞增殖的需要,因此创伤后第2 d出现第一个表达量峰值。核糖体的生物合成是一个高度动态的过程,其合成速率受到外界环境的影响(Kara et al,2006)。目前已知RAS/蛋白激酶A(Toda et al,1987)、TOR(Michael et al,1999)等一些保守的信号途径调节了核糖体的生物合成速率。核糖体达到一定量时,推测RPL17基因可能会通过降低表达量启动某些信号途径,从而阻止核糖体的进一步合成,因此RPL17基因的表达量会出现第1次峰值后的下降。经过第1个阶段表达量的变化,刺参适应了创伤刺激,体壁开始出现再生,RPL17基因又开始大量表达,第5 d时该基因的表达量出现第2个峰值。此后表达量又开始下降,可能是因为细胞增殖积累一定量,细胞活动由增殖变为开始迁移,这与体壁再生的组织学研究结果(李霞等,2007;Qin et al,2015)。本研究发现RPL17和RPL30在仿刺参体壁再生过程中的表达存在一定差异,推测RPL30基因在仿刺参体壁再生过程中主要参与细胞增殖和蛋白质合成,而RPL17在再生阶段的作用可能包括免疫反应和细胞增殖等多种作用。此结果也说明两个核糖体基因在刺参的生理活动中起着不同的作用,这也从侧面证明核糖体蛋白基因除构成核糖体参与蛋白质合成外,还有着各自的核糖体外功能。徐存栓的研究(徐存栓等,2008)也发现肝再生的过程中核糖体蛋白基因也出现差异表达;一些特殊的生理状况如肿瘤中也发现核糖体蛋白基因包括RPL30基因差异表达(王辉等,2007;闫扬等,2007),有研究认为核糖体蛋白可能是细胞增生调节因子的一个新家族,通过作用于细胞周期的某一点,直接影响肿瘤的发展(Shriver et al,1998)。因此推断RPL17、RPL30基因参与了仿刺参的组织生长以及体壁的再生,但是它们是分别还是协同调控仿刺参的再生过程还有待进一步研究。
Garcia-arraras J E,Greenberg M J,2001.Visceral regeneration in holothurians.Microsc Res Tech,55(6):428-451.
Kara A B,Susan J B,2006.Comprehensive mutational analysis of yeast DEXD/H box RNA helicases involved in large ribosomal subunit biogenesis.Molecular and Cellular Biology,26(4):1 195-1 208.
Kondoh N,Shuda M,2001.Enhanced expression of S8,L12,L23a,L27 and L30 ribosomal protein mRNAs in human hepatocelluar carcinoma.Anticancer Res,21:2 429-2 433.
Lü B,Xu J,Zhu Y,2007.Systemic of the differential gene expression profile in a colonic adenoma-normal SSH library.Clin Chim Acat,378: 42-47.
Michael C S,1999.Target of rapamycin(TOR)signaling coordinates tRNA and 5S rRNA gene transcription with growth tRNA and 5S rate in yeast.Gene Therapy And Molecular Biology,4:339-348.
Qin Y,Zhao X,Li X,et al,2015.Characterization of the 26S proteasome S7 subunit gene and tts expression during regeneration in Apostichopus japonicus.Journal of the World Aquaculture Society,46 (2):159-170.
Santiago P,Roig-Lopez J L,Santiago C,et al,2000.Serum amyloid A protein in an echinoderm:Its primary structure and expression during intestinal regeneration in the sea cucumber Holothuriaglaberrima.Exp Zool,288(433):335-344.
Sara A Z,Maria E C,2005.Tor andcyclic AMP-protein kinase A:two parallel pathways regulating expression of genes required for cell growth.Eukaryotic cehull,4(1):63-71.
Shriver S P,Shriver M D,Tirpk D L,1998.Trinucleotide repeat length variation in the human ribosomal protein L14 gene(RPL14):localization to 3p21.3 and loss of heterozygosity in lung and oral cancers.Mulat Res,406:9-23.
Toda T,Cameron S,Sass P,1987.Cloning and characterization of BCY1, a locus encoding a regulatory subunit of the cyclic AMP-dependent protein kinase in Saccharomyces cervisiae.Molecular and Cellular Biology,7(4):1 371-1 377.
Vilardell J,Yu S J,Warner J R,2000.Multiple functions of an evolutionarily conserved RNA binding domain.Mol.Cell,5(4):761-766.
Zheng F X,Sun X Q,Fang B H,et al,2006.Comprarative analysis of genes expressed in regenerating intestine and non-eviscerated intestine of Apostichopus japonicas Selenka(Aspidochirotida:Stichopodidae)and cloning of ependymin gene.Hydrobiologi,571: 109-122.
陈秋实,李霞,段晶晶,等,2008.利用差异显示法研究刺参表皮再生相关基因.生物技术通报,6:124-131.
侯怡铃,侯万儒,2010.大熊猫核糖体蛋白L30亚基基因(RPL30)的cDNA克隆及序列分析.四川大学学报(自然科学版),47(5):1 154-1 158.
姜健,杨宝灵,邰阳,2004.海参资源及其生物活性物质的研究.生物技术通讯,15(5):537-538.
李霞,聂竹兰,魏杰,2007.仿刺参体壁表皮再生组织学和超微结构观察.中国水产科学,14(7):1-6.
李霞,王雪,秦艳杰,等,2012.仿刺参EGFR基因的克隆与表达分析.水产学报,36(1):41-49.
聂竹兰,李霞,2006.海参再生的研究.海洋科学,30(5):78-82.
秦艳杰,李霞,张慧敏,等,2013.仿刺参体壁创伤修复消减文库的构建与分析.大连海洋大学学报,28(3):224-229.
王辉,刘伟利,2007.核糖体蛋白基因表达与肿瘤的关系.生理科学进展,38(4):376-378.
王淑娴,叶海斌,于晓清,等,2012.海参的免疫机制研究.安徽农业科技,40(25):12 553-12 555.
王艳枫,李霞,秦艳杰,2013.刺参核糖体蛋白L17基因(RPL17)全长cDNA克隆及组织表达.农业生物技术学报,21(5):562-569.
徐存栓,杨志利,董华明,2008.核糖体相关基因在大鼠再生肝及肝肿瘤中表达异同的表达.解剖学报,39(3):281-286.
闫扬,孙秀菊,2007.核糖体蛋白基因家族与肿瘤的关系.国际遗传学杂志,30(5):387-390.
赵鹏,杨红生,孙丽娜,2013.仿刺参(Apostichopus japonicus)摄食和运动器官的结构与功能.海洋通报,32(2):178-183.
赵祥吉,秦艳杰,李霞,等,2013.刺参26S蛋白酶体S7亚基基因的克隆与组织表达分析.大连海洋大学学报,28(6):528-534.
朱可可,易聪,周兴涛,2009.核糖体蛋白基因协同表达的分子机制.安徽农业科学,37(35):17 376-17 379.
(本文编辑:袁泽轶)
Expression of two ribosomal protein genes in body wall regeneration in sea cucumber(Apostichopus japonicus)
QIN Yan-jie1,2,WANG Wen-wen1,WANG Yan-feng2,LI Xia1,2
(1.Key Laboratory of MarineBio-resources Restoration and Habitat Reparation of Liaoning Province,Dalian Ocean University,Dalian 116023,China 2.Ministry of Agriculture Key Laboratory of Mariculture&Stock Enhancement in North China's Sea,Dalian Ocean University,Dalian116023,China)
cDNA full-length sequence of ribosomal protein L30(RPL30)gene was cloned in the body wall of sea cucumber(Apostichopus japonicus).In this study,the results showed that the cDNA of RPL30(GenBank accession No. JQ770165)length was 557 bp,including the 56 bp of the 5'-UTR and 162 bp of the 3'-UTR.The open reading frame was 339 bp,encoding polypeptides of 112 amino acids.Real-time PCR was performed to analyze the expression profiling of RPL30.The results showed that the relative expression levels of RPL30 in intestine,coelomocytes,longitudinal muscle,and body wall were 7.24(P<0.01),4.47(P<0.01),3.12(P<0.05)and 1.35(P>0.05)fold of those in the respiratory tree. The expression values of RPL30 gene decreased and then rose during body wall regeneration in A.japonicus,and the peak value,2.13(P<0.05)fold of the control group,occurred at 7d regeneration,and the levels of 1d,5d and 6d were significantly lower than those in the control group.The RPL17 expression level showed two peaks at 2d and 5d regeneration were 7.47(P<0.01)and 5.60(P<0.05)fold of those values in the control group,respectively.The different expressionlevels of RPL17 and RPL30 showed that they would play important,but different roles in the immune response,cell proliferation,protein synthesis and some kinds of regulation.These results would be the necessary basis for mechanism study of protein synthesis,regeneration and other physiological regulation in sea cucumber.
sea cucumber(Apostichopus japonicas);ribosomal protein L30;cloning;ribosomal protein L17;regeneration
S917.4
A
1001-6932(2017)02-0174-08
10.11840/j.issn.1001-6392.2017.02.008
2015-12-25;
2016-02-17
国家自然科学基金(30371099);辽宁省教育厅创新团队(2007T015);辽宁省教育厅实验室专项(2008S064)。
秦艳杰(1977-),博士,副教授,主要从事海洋生物学研究。电子邮箱:qinyanjie@dlou.edu.cn。
李霞,硕士,教授。电子邮箱:qin_tina@163.com。
