长江亚热带浅水湖群藻类浊度与非藻类浊度的变异规律
2017-04-12王海军王洪铸潘保柱梁小民
王海军王洪铸潘保柱,梁小民
(1. 中国科学院水生生物研究所淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 长江科学院水力学研究所, 武汉 430010)
长江亚热带浅水湖群藻类浊度与非藻类浊度的变异规律
王海军1王洪铸1潘保柱1,2梁小民1
(1. 中国科学院水生生物研究所淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 长江科学院水力学研究所, 武汉 430010)
浊度的上升是湖泊富营养化过程中沉水植物消失、由草向藻发生稳态转换的根本原因。研究利用长江中下游浅水湖群区域数据对藻类浊度(TurbAlg)和非藻类浊度(TurbNonAlg)及其对总浊度(TurbTot)的贡献率(TurbAlg%, TurbNonAlg%)进行了推算。结果显示, 在该区域非藻类浊度绝对值及其贡献率均普遍高于藻类浊度; TurbAlg主要受限于总磷(TP), 而TurbNonAlg受水深(ZM)、TP和沉水植物等多重因素的影响。TurbAlg和TurbNonAlg呈现出随着TP浓度的升高而持续上升的趋势, 表明尽管稳态转换存在特定的阈值, 但整个富营养化过程中均需加强对营养物浓度的控制。
稳态转换; 藻类浊度; 非藻类浊度; 湖盆形态; 富营养化; 长江浅水湖群
越来越多的研究证据表明浅水湖泊生态系统存在着多稳态性[1—4]。在低营养时, 湖泊处于以沉水植物占优的清水稳态(简称草型稳态), 在高营养时则处于以浮游藻类占优的浊水稳态(简称藻型稳态), 而在中等营养时湖泊可能同时存在这两种状态[1]。研究这种多稳态的维持及稳态之间的转换具有重要的理论和实践意义, 可为风险规避和目标管理提供定量化的科学依据, 以维持有利稳态避免不利转换或实现有利转换。
浅水湖泊草藻稳态各有内在的正反馈机制维持其当前稳态。Scheffer根据温带湖泊研究结果对这种正反馈的各种过程进行了形象地阐释(图1)。沉水植物与浊度之间的相互作用是这种正反馈机制的核心, 即草型稳态中沉水植物可通过各种途径抑制浊度的增加从而为自身的进一步发展提供水下光照保障; 相反, 在藻型稳态中浊度的增加进一步抑制了沉水植物的恢复。因此, 多稳态的维持与稳态转换的发生在很大程度上取决于浊度的水平及其变化, 而浊度(总浊度, TurbTot)又由浮游藻类(这里命名为藻类浊度TurbAlg)和沉积物再悬浮(非藻类浊度TurbNonAlg)决定。也因此, 很有必要将TurbAlg与TurbNonAlg进行定量化, 以增进对多稳态维持与稳态转换的理解。
长江中下游地处亚热带, 分布着大量的浅水湖泊。因受到人为富营养化等因素的影响, 该湖群正逐步从草型稳态向藻型稳态发生转换。对于该区域湖泊的稳态转换问题已有一定的研究工作, 且主要侧重于分析转换发生的阈值[5—9]。本研究的目的在于对TurbAlg和TurbNonAlg及其对总浊度(TurbTot)的贡献(TurbAlg%, TurbNonAlg%)进行定量化, 进而分析湖盆形态、营养物水平和沉水植被等关键因子对这些浊度特征参数的影响, 以解析长江中下游浅水湖群浊度组分特征及稳态转换过程中这些浊度特征的变化规律。
1 研究湖泊与方法
研究湖泊位于长江中下游流域。多湖比较共有28个湖泊50湖年的数据。其中本研究调查的有26个湖泊(东经113°10′—121°00′, 北纬29°30′—32°00′) 36湖年的数据, 包括2001年至2002年间对7个湖泊的周年逐月调查, 2003年、2004年和2005年分别对16个、8个和3个湖泊的周年逐季度调查。从文献收集的有7个湖泊14湖年的数据, 其中从胡传林等[10]和梁彦龄等[11]分别收集了3个湖泊1987年和1992年周年逐月调查的共6周年数据, 从崔奕波等[12]收集了4个湖泊1998年和1999年周年逐季度调查的共8周年数据。
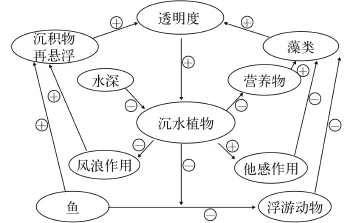
图 1 浅水湖泊草型清水和藻型浊水稳态的正反馈维持机制(译自Scheffer, 1998)Fig. 1 The positive feedback mechanism to maintain the macrophyte-dominated clear water and algae-dominated turbid water of shallow lakes (translated from Scheffer, 1998) (+, -means promoting and inhibiting, respectively)
根据湖泊大小每湖设置3—12个水样采集点,在每两个水样点间增加一个样点测量水深(ZM)、透明度(ZSD)和沉水植物生物量(BMac), 即ZM、ZSD和BMac这3个指标的样点数为每湖5—23个样点。水样采自每个点的表层(水面以下0.5 m)和底层, 混合后带回实验室进行分析。TP和浮游藻类叶绿素a (Chl. a)的测定是在实验室中按照国家标准进行。TP的测定方法是钼锑抗分光光度法。Chl. a的测定方法是通过丙酮萃取, 用分光光度计(Unico UV-2000, Shanghai, China)读取665 nm和750 nm波长的吸光率。ZM、ZSD分别采用测深锤、透明度盘测量。沉水植物使用镰刀型采草器(采集面积为1/5 m2)进行采集, 在每个点采集2—4次, 样品混合。植物采回后清洗底泥、枯草等杂质, 称量湿重。
为了区分浮游藻类和除藻类之外的悬浮颗粒对浊度的相对贡献, 我们参考Pørtielje等[13]、Ibelings等[14]的做法, 将TurbTot(ZSD的倒数)分成3个部分, 即TurbAlg、TurbNonAlg和无悬浮颗粒时的背景浊度(TurbBck)。从理论上讲, 对这些浊度的计算最好是用长江浅水湖群数据构建的方程来进行。然而,在长江流域有着很长历史的渔业活动可在很大程度上促进沉积物的再悬浮, 影响到TurbBck进而影响TurbAlg和TurbNonAlg的推算。因此, 本研究将直接借用Pørtielje等基于荷兰浅水湖泊构建的方程进行各种浊度分析。在他们的推算方法中, 数据集中最小的浊度值(即最大ZSD)0.16/m设为TurbBck, 假定此时浮游藻类和非藻类悬浮颗粒物不存在。另外, 通过建立Chl. a与TurbTot(1/ZSD)的经验关系, 在这种散点关系中可以找到一条线使得Chl. a浓度有一个对应的最小1/ZSD值, 将该值定义为除去TurbBck后浮游藻类所造成的浊度。这条线的斜率取值所有样点(TurbTot-TurbBck)/Chl. a的5%, 结果为0.01/m。因此, 非藻类浊度的计算公式为: TurbNonAlg=1/ZSD-0.16-0.01 Chl. a。
STATISTICA 8.0和Microsoft Excel 2010用于数据处理和统计分析。所用的数据均为全湖周年均值。Unequal N Post-hoc ANOVA用于检验样本含量n值不相等时两组数据平均值的差异。
2 结果
2.1 浊度组分特征
通过经验公式推算了长江浅水湖群藻类浊度(TurbAlg, /m)和非藻类浊度(TurbNonAlg, /m)的大小(图 2)。结果显示, TurbNonAlg平均高达0.96/m, 最小0.11/m, 最大2.43/m。而TurbAlg则明显较小, 平均只有0.19/m, 最小0.02/m, 最大1.39/m。从所占总浊度百分比来看, TurbNonAlg%平均超过71.0%, 最小36.0%, 最大92.0%; 而TurbAlg%平均仅约11.0%, 最小0.5%, 最大47.0%。
2.2 浊度组分与湖盆形态的关系
选择代表湖盆形态的面积(Area)和平均ZM这两个参数, 分析其与TurbAlg、TurbNonAlg、TurbAlg%和TurbNonAlg%的关系(图 3)。结果显示, Area与这些参数的关系都不显著(图 3a, b)。TurbAlg和TurbAlg%与ZM的关系也都不显著, 但TurbNonAlg和TurbNonAlg%均随ZM的增加而显著下降(图 3c, d)。
2.3 浊度组分与湖水总磷的关系
在TP尺度上TurbAlg和TurbNonAlg均有显著上升的趋势(图 4a), 且TurbAlg在TP超过约100 μg/L以后上升加快。从两种浊度所占百分比来看, TurbAlg%同样是在TP超过约100 μg/L以后上升加快(图 4b), TurbNonAlg%则是在这个浓度水平之后开始下降。总体来说, 在整个TP尺度上, 非藻类浊度绝对值及其所占百分比普遍高于藻类浊度。
2.4 浊度组分与沉水植物的关系
在有草湖泊中的TurbAlg和TurbAlg%分别平均为0.05/m和6.4%, 均显著低于无草湖泊中的0.44/m和18.7% (post-hocunequal NHSD ANOVA, P<0.001)。有草湖泊中的TurbNonAlg(平均0.67/m)显著低于无草湖泊(平均1.46/m)(P<0.001)。TurbNonAlg%在有草和无草湖泊中分别平均70.5%和72.1%, 二者差异不显著(P=0.73)。各浊度参数与BMac之间的关系显示(图 5), 有草湖泊中TurbNonAlg和TurbNonAlg%均随BMac的下降而显著上升, 而TurbAlg和TurbAlg%随BMac的变化规律均不显著。

图 2 藻类浊度(TurbAlg, /m)、非藻类浊度(TurbNonAlg, /m) (a)及其占总浊度百分比(TurbAlg%, TurbNonAlg%) (b)的分布 (n=50)Fig. 2 Distribution of algal turbudity (TurbAlg, /m), non-algal turbidity (TurbNonAlg, /m) and their contribution to total turbidity in percentage (TurbAlg%, TurbNonAlg%) (n=50)

图 3 藻类浊度(TurbAlg, /m)、非藻类浊度(TurbNonAlg, /m)及其占总浊度百分比(TurbAlg%, TurbNonAlg%)与湖泊面积(Area, km2)和水深(ZM, m)的关系(n=50)Fig. 3 The relationships of lakes area (Area, km2) and water depth (ZM, m) with algal turbidity (TurbAlg, /m), non-algal turbidity (TurbNonAlg, /m), and their contribution to total turbidity in percentage (TurbAlg%, TurbNonAlg%)
3 讨论
关于藻类和非藻类悬浮颗粒引起的浊度, 因缺乏有效的直接测定方法, 已有研究对于各种组分浊度的分析普遍基于间接推算。Kirk[15]和Buiteveld[16]曾分别通过经验回归和理论推导找到了消光系数及TurbTot与叶绿素、碎屑和无机颗粒之间的定量关系。这些关系经过独立验证后被证明具有较好的外推性[1]。但是从这些关系中, 我们仍难以分别来定量解析各种颗粒的独立贡献。Pørtielje等[13]基于大量的荷兰温带浅水湖泊数据将叶绿素与TurbTot进行直接耦合, 并通过一些推算得出TurbTot与TurbAlg和TurbNonAlg之间的定量关系。这种浊度计算方法有它的局限性, 即ZSD和浊度的内涵并不完全一样, TurbBck所假定的这些情形在真实生态系统中也并不存在, 不能完全去除藻类和非藻类悬浮物的作用。但是, 这种推算仍然有必要, 也可有效地区别藻类和非藻类对浊度的相对贡献。
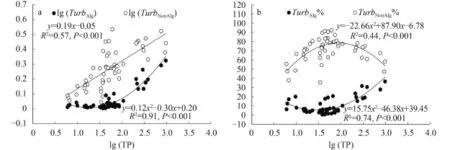
图 4 藻类浊度(TurbAlg, /m)、非藻类浊度(TurbNonAlg, /m)及其占总浊度百分比(TurbAlg%, TurbNonAlg%)与湖水总磷(TP, μg/L)的关系(n=50)Fig. 4 The relationships of total phosphorous (TP, μg/L) of lake water with algal turbidity (TurbAlg, /m), non-algal turbidity (TurbNonAlg, /m), and their contribution to the total turbidity in percentage (TurbAlg%, TurbNonAlg%)

图 5 藻类浊度(TurbAlg, /m)、非藻类浊度(TurbNonAlg, /m)及其占总浊度百分比(TurbAlg%, TurbNon-Alg%)与沉水植物生物量(BMac, g/m2)的关系(n=50)(实心为草型湖泊, 空心为藻型湖泊; 横坐标显示的逆序刻度以体现沉水植物的消失过程; 空心点在数值上做了处理以方便辨别两组数据)Fig. 5 The relationships of submersed macrophyte biomass (BMac, g/m2) with algal turbidity (TurbAlg, /m), non-algal turbidity (TurbNonAlg, /m), and their contribution to the total turbidity in percentage (TurbAlg%, TurbNonAlg%)
从本研究分析的结果可以看出, 在长江浅水湖群中非藻类颗粒对TurbTot的贡献要明显大于浮游藻类。将Pørtielje等[13]所拟合的公式与本研究的数据进行比较(图 6), 可以看出长江浅水湖群特定Chl. a对应的最小浊度要远高于荷兰浅水湖泊。这说明长江浅水湖群中TurbNonAlg在TurbTot中的贡献要远大于荷兰湖泊。利用Pørtielje的公式对丹麦浅水湖泊进行计算(由Aarhus University的Martin Søndergaard提供原始数据)[17], 结果显示TurbAlg%平均约为30.0%, 远高于本研究11.0%; 而TurbNonAlg%平均为46.5%, 远低于本研究的71.0%。本研究中涉及的ZM、TP和沉水植物均未能解释这种差异。丹麦湖泊平均ZM约为1.5 m, 明显低于本研究结果, 理应对应着更高的TurbNonAlg; TP平均为110 μg/L, 与本研究相当; 沉水植物因单位不同未能直接比较, 但本研究的结果显示有草无草湖泊的TurbNonAlg贡献基本一致。这种差异更可能是源于长江浅水湖群相比于荷兰和丹麦湖泊承受着高强度的渔业活动, 尽管这个结论尚需更直接的证据。
对比TurbAlg和TurbNonAlg与各种关键影响因子之间的关系, 可以看出TurbAlg主要受限于TP; 对于TurbNonAlg而言ZM、TP和BMac均可产生重要影响。TurbNonAlg与ZM之间密切的关系, 意味着对湖泊稳态转换而言ZM比Area具有更重要的意义。这一点在Wang等[4]中也得到了阐述。在湖泊TP和BMac尺度上, 尽管TurbNonAlg%在超过一定浓度的TP时有一个下降过程, 在水草消失后与消失前的差别也不大,但在这之前其贡献率均有个逐步上升过程, 而且非藻类浊度的绝对值在整个过程中一直呈上升趋势。因此, 尽管湖泊发生稳态转换有特定的阈值[4,14,18],浊度的控制在整个富营养化过程中仍具有重要的意义。
[1]Scheffer M. Ecology of Shallow Lakes [M]. The Netherlands: Kluwer Academic Publishers. 1998, 357
[2]Jeppesen E, Jensen J P, Søndergaard M, et al. Changes in nitrogen retention in shallow eutrophic lakes following a decline in density of cyprinids [J]. Archiv fur Hydrobiology, 1998, 142(2): 129—151
[3]Scheffer M, Carpenter S R. Catastrophic regime shifts in ecosystems: linking theory to observation [J]. Trends in Ecology and Evolution, 2003, 18(12): 648—656
[4]Wang H J, Wang H Z, Liang X M, et al. Total phosphorus thresholds for regime shifts are nearly equal in subtropical and temperate shallow lakes with moderate depths and areas [J]. Freshwater Biology, 2014, 59(8): 1659—1671
[5]Yang X D, Shen J, Dong X H, et al. Historical trophic evolutions and their ecological responses from shallow lakes in the middle and lower reaches of the Yangtze River: Case studies on Longgan Lake and Taibai Lake [J]. Science in China: Series D Earth Sciences, 2006, 49(Supp.1): 51—61
[6]Nian Y G, Song Y W, Li Y J, et al. Regime shift theory and ecological restoration discussion in eutrophic shallow lakes [J]. Research of Environment Sciences, 2006, 19(1): 67—70 [年跃刚, 宋英伟, 李英杰, 等. 富营养化浅水湖泊稳态转换理论与生态恢复理论. 环境科学研究, 2006, 19(1): 67—70]
[7]Chen K N, Bao X M, Shi L X, et al. Ecological restoration engineering in Lake Wuli, Lake Taihu: a large enclosure experiment [J]. Journal of Lake Sciences, 2006, 18(2): 139—149 [陈开宁, 包先明, 史龙新, 等. 太湖五里湖生态重建示范工程-大型围隔实验. 湖泊科学, 2006, 18(2): 139—149]
[8]Wang H J, Liang X M, Jiang P H, et al. TN: TP ratio and planktivorous fish do not affect nutrient-chlorophyll relationships in shallow lakes [J]. Freshwater Biology, 2008, 53: 935—944
[9]Chang F Y, Pan X J, Kang L J, et al. A study on the growth and some physiological characteristics of Anabaena spiroides isolated from Lake Erhai [J]. Acta Hydrobiologica Sinica, 2009, 33(3): 385—390 [常锋毅, 潘晓洁, 康丽娟, 等. 洱海螺旋鱼腥藻生长生理特征的初步研究. 水生生物学报, 2009, 33(3): 385—390]
[10]Hu C L, Huang X F. Collected Papers on the Fishery Ecology and Exploitation Technology of the Bao’an Lake [M]. Beijing: Science Press. 1991, 137 [胡传林, 黄祥飞.保安湖渔业生态和渔业开发技术研究文集.北京: 科学出版社. 1991, 137]
[11]Liang Y L, Liu H Q. Resources, Environment and Fishery Ecological Management of Macrophytic Lake (No. 1) [M]. Beijing: Science Press. 1995, 355 [梁彦龄, 刘伙泉.草型湖泊资源、环境与渔业生态管理(一). 北京: 科学出版社. 1995, 355]
[12]Cui Y B, Li Z J. Fishery Resources and Conservation of Environment in Lakes of the Changjiang River Basin [M]. Beijing: Science Press. 2005, 424 [崔奕波, 李钟杰. 长江流域湖泊的渔业资源与环境保护. 北京: 科学出版社. 2005, 424]
[13]Pørtielje R, Van der Molen D T. Relationships between eutrophication variables: from nutrient loading to transparency [J]. Hydrobiologia, 1999, 408: 375—387
[14]Ibelings B W, Portielje R, Lammens E H R R, et al. Resilience of alternative stable states during the recovery of shallow lakes from eutrophication: Lake Veluwe as a case study [J]. Ecosystems, 2007, 10(1): 4—16
[15]Kirk J T O. Characteristics of the light-field in highly turbid waters-a monte-carlo study [J]. Limnology and Oceanography, 1994, 39(3): 702—706
[16]Buiteveld H. A model for calculation of diffuse light attenuation (PAR) and Secchi depth [J]. Netherlands Journal of Aquatic Ecology, 1995, 29(1): 55—65
[17]Van den Berg M S, Coops H, Meijer M L, et al. Clear water associated with a dense Chara vegetation in the shallow and turbid Lake Veluwemeer, the Netherlands. In: Jeppesen E, Søndergaard M, Christoffersen K (Eds.), Structuring Role of Submerged Macrophytes in Lakes [M]. New York: Springer. 1998, 339—352
[18]Jeppesen E, Jensen J P, Kristensen P, et al. Fish manipulation as a lake restoration tool in shallow, eutrophic, temperate lakes. 2. threshold levels, long-term stability and conclusions [J]. Hydrobiologia, 1990, 200(1): 219—227
VARIATIONS OF ALGAL AND NON-ALGAL TURBIDITY AMONG THE YANGTZE SUBTROPICAL SHALLOW LAKES
WANG Hai-Jun1, WANG Hong-Zhu1, PAN Bao-Zhu1,2and LIANG Xiao-Min1
(1. State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China; 2. Yangtze River Scientific Research Institute, Wuhan 430010, China)
Increasing turbidity is the main cause for the disappearance of submersed macrophytes and regime shift from clear-water to turbid-water states along the eutrophication of lakes. Quantitative discrimination on the contribution of algal and non-algal particles to total turbidity (TurbTot) may enhance the understanding on these processes. This study investigated algal turbidity (TurbAlg), non-algal turbidity (TurbNonAlg), and their contribution to TurbTotin percentage (TurbAlg%, TurbNonAlg%) using data from Yangtze shallow lakes. The results showed that both absolute and percentage non-algal turbidity (averaged 0.96/m and 71.0%) were much higher than those of algal turbidity (averaged 0.19/m and 11.0%). Total phosphorus (TP) positively impact TurbAlgand TurbNonAlg, and when TP exceeded 100 μg/L, TurbAlgand TurbAlg% showed accelerated increases while TurbNonAlg% showed a decreasing trend. TurbAlg, TurbAlg% and TurbNonAlgwere significantly lower in macrophyte-present lakes than in macrophyte-absent lakes, while TurbNonAlg% was not significantly different between lakes. TurbNonAlgand TurbNonAlg% increased significantly with decreased biomass of submersed macrophytes (BMac), while TurbAlgand TurbAlg% showed no significant varying trend. In summary, TurbAlgwas determined mainly by TP, while TurbNonAlgwas greatly determined by ZM, TP, and submersed macrophytes. The fact of continuously increasing TurbAlgand TurbNonAlgalong with increasing TP and degrading macrophytes demonstrates that nutrient control and macrophyte maintenance are practically important during the whole process of eutrophication, although there exists a threshold for clear-to-turbid regime shifts.
Regime shifts; Algal turbidity; Non-algal turbidity; Lake morphology; Eutrophication; Yangtze shallow lakes
Q142
A
1000-3207(2017)02-0414-06
10.7541/2017.51
2015-10-23;
2016-04-17
国家自然科学基金(30900194); 国家重大基础研究发展计划(2008CB418006); 湖北省科技支撑项目 (2015BBA225); 淡水生态与生物技术国家重点实验室(2014FB14和2011FBZ14); 中国科学院重大交叉项目(KZCX1-YW-14-1)和重大方向性项目(KZCX2-YW-426-02); 中国科学院青年创新促进会基金(2014312) 资助 [Supported by the National Natural Science Foundation of China (30900194); 973 Programs (2008CB418006); Hubei Province (2015BBA225); State Key Laboratory of Freshwater Ecology and Biotechnology (2014FB14 and 2011FBZ14); Chinese Academy of Sciences (KZCX1-YW-14-1 and KZCX2-YW-426-02); Youth Innovation Promotion Association of Chinese Academy of Sciences (2014312)]
王海军(1978—), 男, 江西玉山人; 博士; 主要从事淡水生态学研究。E-mail: wanghj@ihb.ac.cn
梁小民, E-mail: liangxm@ihb.ac.cn
