A族链球菌多价疫苗的完善制备及其动物免疫保护作用研究①
2017-04-10王家超邹东花郭奕阳张征峥杨丽娟马翠卿
王家超 袁 婷 邹东花 高 雪 郭奕阳 李 剑 张征峥 杨丽娟 马翠卿
(河北医科大学免疫教研室,河北省重大疾病的免疫机制及干预重点实验室,石家庄050017)
·免疫学技术与方法·
A族链球菌多价疫苗的完善制备及其动物免疫保护作用研究①
王家超 袁 婷②邹东花 高 雪 郭奕阳③李 剑 张征峥 杨丽娟 马翠卿
(河北医科大学免疫教研室,河北省重大疾病的免疫机制及干预重点实验室,石家庄050017)
目的:完善A族链球菌(GAS)多价疫苗并探究其免疫保护效果。方法:构建GAS的多肽疫苗F7M6,将其中的M6多肽克隆作为对照,表达纯化后,将两种蛋白连同本室保存的GAS M1蛋白分别免疫小鼠,末次免疫后10 d,半数小鼠行ELISPOT检测IL-4和IFN-γ,ELISA检测各组的IgG、IgG1、IgG2a;同时,以F7M6蛋白为诊断抗原检测其与抗链“O”阳性人血清的特异性结合;致死量的M1GAS攻毒并计算存活率。结果:无论是诱导细胞免疫,还是诱导抗体产生,F7M6组的水平均为最高;攻毒后F7M6蛋白的生存率为66.7%。F7M6蛋白检测抗链“O”阳性血清的阳性检出率为95%。结论:GAS的多肽疫苗F7M6能诱导特异性细胞和体液免疫应答,对GAS感染具有良好的保护效果,且F7M6的多价表位涵盖于时下流行的大部分GAS的血清型中。
A族链球菌(GAS);多价疫苗;M蛋白;FbaA蛋白;免疫应答
A族链球菌(Group A streptococcus,GAS)是临床上最常见也是致病性最强的革兰阳性菌之一,它能够引起多种化脓性感染,同时,它还可以导致一系列的自身免疫性疾病,如风湿性心脏病、肾小球肾炎等[1]。随着耐药菌株的出现,链球菌的感染表现出在宿主体内长期存活及反复发作的特点[2,3],因此GAS感染已成为临床治疗中的棘手问题。本课题组一直致力于寻找针对GAS的有效疫苗,并筛选到了既不与人体组织发生交叉反应又高效的多个M蛋白及FbaA的表位[4-7]。由于GAS有众多血清型,近几年,发现GAS的M12在我国多地普遍流行[8,9],基于此,本实验通过网上比对和预测,选取M12的M蛋白中既跟人无交叉反应又具有免疫原性的表位基因片段,通过PCR将M12的含多个表位基因片段与验证过的FbaA的7个优势表位和GAS M1、M3、M6和M18的M蛋白的有效表位基因及M蛋白具好的免疫原性的C端保守序列串联在一起[10],再在氨基端加上木马序列(TA)和一个多聚丙氨酸DR表位(PADRE),以促进抗原提呈及平衡Th淋巴细胞反应,获得重组多价疫苗基因片段命名为f7m6,及通过PCR获得了单纯的GAS 6个型别的M蛋白的重组基因片段,命名为m6,并将上述的f7m6及m6克隆至pET28a原核载体并表达。将纯化的F7M6、M6及科室保存的来自GAS M1血清型的M蛋白[11],佐以目前人用免疫效果佳的MF59佐剂,免疫动物,观察该重组疫苗的免疫效果,并通过GAS攻击小鼠观察小鼠存活率来判定重组多肽对小鼠的免疫保护作用,以期探寻到适合中国地区应用的高效GAS疫苗。
1 材料与方法
1.1 材料
1.1.1 实验动物、菌株及主要试剂 BALB/c雌性小鼠,4~6周龄,购自北京维通利华实验动物技术有限公司;菌株E.coli.DH5α、菌株E.coli.BL21、质粒pET28a、菌株pET28a/F7M5、M1 GAS标准菌株皆为本室保存,GAS临床菌株河北省儿童医院惠赠;健康儿童血清石家庄市保健院;BamH Ⅰ、HindⅢ、T4DNA 连接酶、DNA分子量标准皆为TaKaRa公司;DNA片段回收试剂盒购自OMEGA公司;镍料购自Bio-Works公司;羊抗鼠IgA-AP、羊抗鼠IgG-AP、羊抗鼠IgG1-AP、羊抗鼠IgG2a-AP皆为SouthernBiotech公司;羊抗人IgG-HRP购自北京中杉金桥生物技术公司。
1.1.2 实验动物 将48只4~6周龄雌性BALB/c小鼠随机分成4组,每组12只,分别免疫F7M6、M6、M蛋白、PBS对照作为对照。
1.2 方法
1.2.1 F7M6特异性序列的基因克隆 通过对我国GAS流行病学的调查研究结果[8,9]选取M12血清型,通过美国疾病预防和控制中心(CDC)网站数据库(http://www.cdc.gov/ncidod/biotech/strep/str-epin-dex.htm)提供的GAS M12血清型的氨基酸序列与GenBank中包含的所有人类蛋白质序列进行比对(http://www.ncbi.nlm.nih.gov/blast Bla-st.cgi),比对结果如果连续5个氨基酸残基与人类某种蛋白的某段序列完全重合,则这5个氨基酸前面的多肽即为候选多肽[12],将该段候选多肽插入本室前期成功构建重组多肽疫苗F7M5基因序列中。借助primer5.0,设计引物。
引物由上海生物工程技术服务有限公司合成,将反应产物进行2%琼脂糖凝胶电泳鉴定。
1.2.2 M6特异性序列的基因克隆 以构建好的f7m6序列为模板,上游引物:5′-TACAGGATCCAACGGTGACGGTAATCCGC-3′,下游引物即为P1148-1216下游引物,进行PCR扩增。
1.2.3 重组质粒的构建 将PCR产物和pET28a质粒分别用BamHⅠ和HindⅢ双酶切,37℃水浴3 h。将酶切产物行纯化回收。酶切后的PCR产物克隆至pET28a酶切鉴定及测序,鉴定出阳性克隆。
1.2.4 重组质粒的表达及纯化 挑取阳性克隆单菌落接种于20 ml 含Kan的2×YT液体培养基中,37℃震摇过夜。次日1∶50转种于500 ml 2×YT液体培养基中,摇至菌密度为 A600=0.7 时,加入IPTG(终浓度为 1 mmol/L)诱导表达4 h。收集菌液,8 000 r/min离心15 min,弃上清,菌体沉淀超声破菌;采用镍料亲和层析的方法纯化目的蛋白,Bradford 法定量重组蛋白,Western blot鉴定重组蛋白的特异性。
表1 PCR引物
Tab.1 Primer of PCR
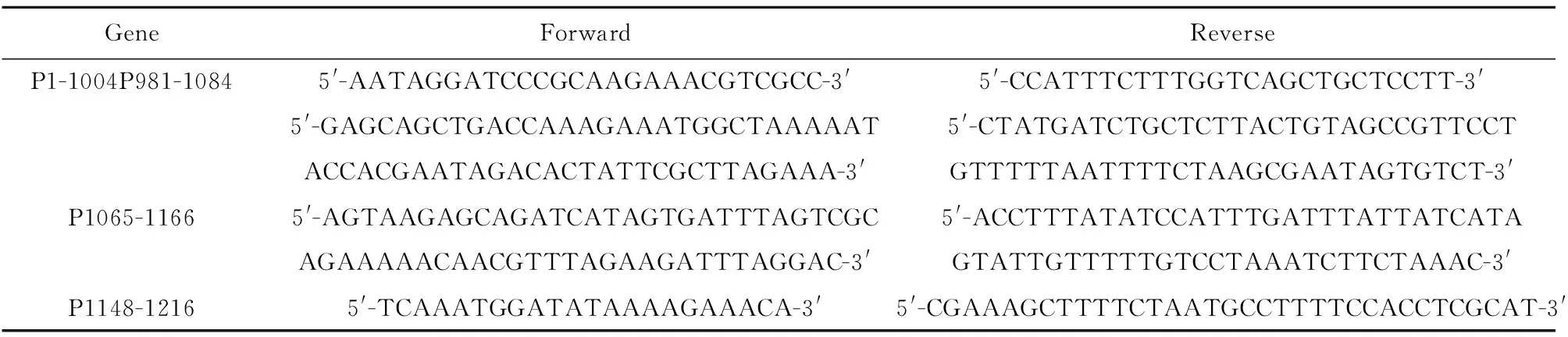
GeneForwardReverseP1-1004P981-10845'-AATAGGATCCCGCAAGAAACGTCGCC-3'5'-CCATTTCTTTGGTCAGCTGCTCCTT-3'5'-GAGCAGCTGACCAAAGAAATGGCTAAAAAT5'-CTATGATCTGCTCTTACTGTAGCCGTTCCTACCACGAATAGACACTATTCGCTTAGAAA-3'GTTTTTAATTTTCTAAGCGAATAGTGTCT-3'P1065-11665'-AGTAAGAGCAGATCATAGTGATTTAGTCGC5'-ACCTTTATATCCATTTGATTTATTATCATAAGAAAAACAACGTTTAGAAGATTTAGGAC-3'GTATTGTTTTTGTCCTAAATCTTCTAAAC-3'P1148-12165'-TCAAATGGATATAAAAGAAACA-3'5'-CGAAAGCTTTTCTAATGCCTTTTCCACCTCGCAT-3'
1.2.5 在线分析F7M6表位序列的亲水性及空间结构的预测 根据F7M6氨基酸顺序在线预测蛋白的抗原表位(http://bio.dfci.harvard.edu/Tool/antigenic.pl)和F7M6全序列亲水性分析(http://web.expasy.org/protscale/)及F7M6重组多肽空间结构的预测(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index)。
1.2.6 免疫途径、剂量及次数 抗原与MF59佐剂1∶1比例混合并充分乳化,每只小鼠皮下免疫总量为20 μg/200 μl。间隔21 d进行第2次常规免疫,共免疫4次。对照组注射等量PBS。
1.2.7 血清的采集和制备 每次免疫10 d后,下颌窦静脉采血,血液4℃放置30 min,3 000 r/min离心30 min,吸取血清,-80℃冻存备用。
1.2.8 动物实验免疫效果分析
1.2.8.1 体液免疫效果测定 以F7M6包被ELISA板,间接ELISA测定血清中IgG、IgG1、IgG2a抗体水平及动态变化。同时,以F7M6作为诊断抗原包被ELISA板,检测临床抗链“O”阳性血清及健康儿童血清的阳性率。酶标仪测405nm波长下的OD值。阳性判断:实验组OD值/阴性对照≥2.1为阳性,效价的判定为出现阳性且OD值>0.15的最高稀释度。
1.2.8.2 细胞免疫效果测定 ELISPOT检测各组小鼠脾脏免疫细胞表达IL-4和IFN-γ的水平。在96孔滤膜板中每孔加100 μl 70%乙醇,室温放置10 min,弃乙醇,PBS洗涤后,加100 μl/孔1∶250稀释的IL-4和IFN-γ抗体包被该板,4℃过夜;次日弃上清,用包被缓冲液洗涤后,加200 μl/孔含10%小牛血清的RPMI1640培养液,室温封闭1 h,弃去封闭液,加入脾细胞悬液50 μl/孔(浓度为2×107个/ml)。每只小鼠设2个组:抗原刺激组和空白组。抗原刺激组每孔补足20 μg/50 μl含相应蛋白的RPMI1640,空白组每孔仅补RPMI1640 50 μl。置CO2培养箱培养48 h后,PBST洗涤,每孔加入100 μl 1∶250稀释的生物素标记的检测抗体,室温孵育2 h。弃上清,PBST洗涤后,每孔加100 μl 稀释的亲和素标记的辣根过氧化物酶,室温孵育45 min。弃上清,PBST洗3次。每孔加100 μl新鲜配制的AEC显色液,室温孵育30 min,去离子水洗涤以终止显色,自然干燥,读板机读板。
1.2.8.3 链球菌攻击小鼠检测疫苗的保护率 末次免疫后10 d,每只小鼠腹腔攻击108CFU/ml M1GAS,连续观察45 d,记录死亡时间和数量,计算存活率,评估疫苗的保护效率。
2 结果
2.1 F7M6和M6的成功表达 以引物自身为模板行搭桥PCR成功扩增出M12血清型中与人无交叉反应的部分基因片段并插入本室前期合成的F7M5序列中[4],得到F7M6重组序列,经2%琼脂糖凝胶电泳,结果在1 200 bp左右出现基因条带,大小与预计相符。以F7M6序列为模板,PCR扩增得到M6序列,结果在750 bp左右出现条带,大小与预计相符(图1A)。纯化后的两段PCR产物分别克隆至载体pET28a上,酶切鉴定条带位置正确(图1B),阳性质粒测序正确。
将鉴定阳性质粒pET28a/F7M6和pET28a/M6转化至表达菌株E.coli BL21中,经IPTG诱导,表达产物主要以包涵体形式存在。行SDS-PAGE电泳,显示F7M6蛋白约为60 kD,M6蛋白约为30 kD,结果与预期一致(图1C)。纯化复性后的两种蛋白均可与F7M5抗体杂交(图1C)。
2.2 网上在线预测F7M6的表位和结构 网上在线预测F7M6多肽的表位(图2A)及其空间结构预测结果(图2B)提示F7M6中有多个表位,且重组表达的多肽能够折叠成复杂的空间结构。F7M6序列亲水性分析结果(图2C)所示,提示该重组蛋白具有良好的亲水性
2.3 F7M6诱导抗体的产生情况 小鼠末次后,ELISA检测小鼠血清IgG抗体水平结果显示:检测到蛋白免疫组小鼠诱导出了高水平IgG抗体,且均显著高于PBS对照组(P<0.001)(图3A)。计算各组效价,F7M6组达到6.7×105,M6组达到5.8×104,M蛋白组达到3.3×104,(图3B)。对第1次、第2次和第3次免疫后的血清1∶3 000稀释后检测IgG动态变化显示,各蛋白免疫组的IgG水平随着免疫次数的增加逐渐增高(图3C)。
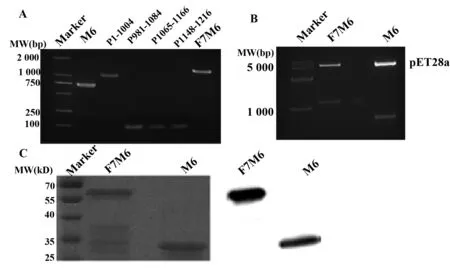
图1 F7M6和M6重组序列的构建和表达Fig.1 Construction and expression of recombinant F7M6 and M6Note: A.Amplification of F7M6 gene and M6 gene sequence by PCR;B.Recombinant plasmid pET28a/F7M6 or pET28a/M6 digested by BamH Ⅰ/Hind Ⅲ;C.Expression detection of F7M6 and M6 by SDS-PAGE and Western blot.
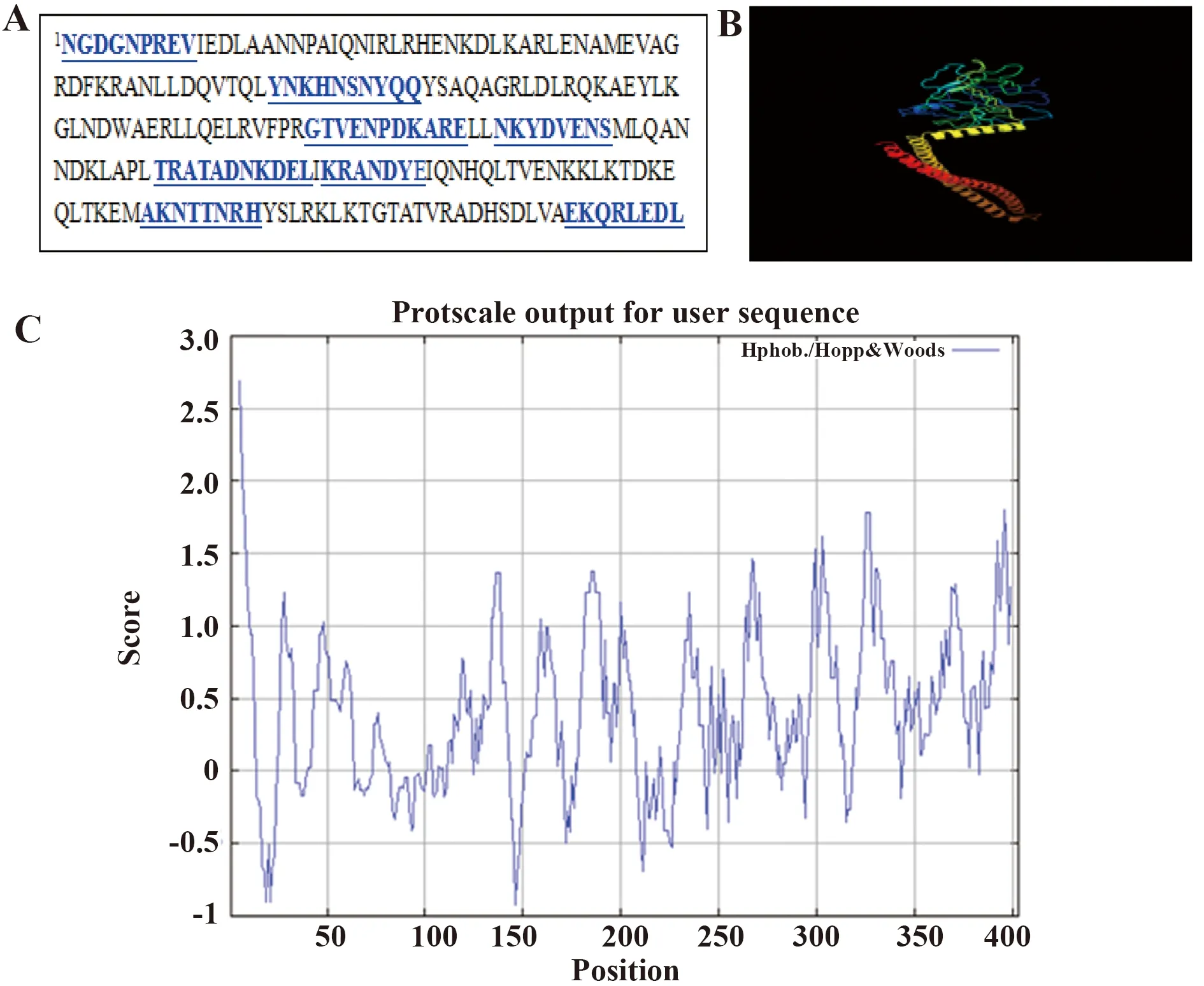
图2 网上在线预测F7M6多肽的表位Fig.2 Online prediction results of F7M6Note: A.The predicted epitopes of F7M6 gene;B.The schematic diagram of F7M6 protein;C.The hydrophilic analysis of F7M6 protein.
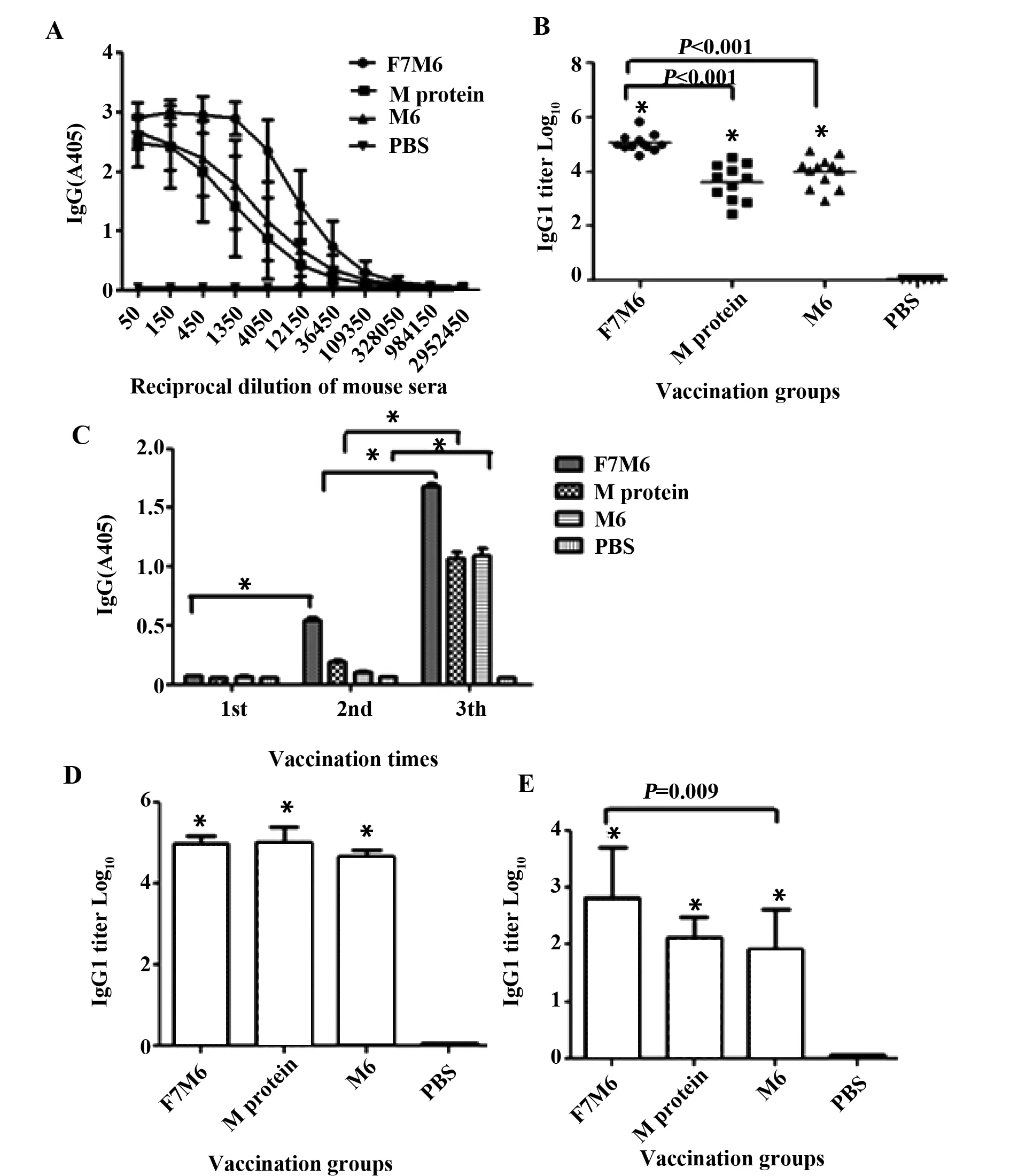
图3 ELISA检测小鼠免疫血清中特异性IgG抗体水平Fig.3 Detection of specific IgG,IgG1 and IgG2a antibody levels in mouse sera by ELISANote: A.Analysis of IgG binding to the three proteins using immunized sera on day 10 post-last vaccination;B.IgG antibody levels of the three group.*.P<0.05 vs PBS;C.Detection of serum IgG antibodies specific to the three proteins following three immunizations.*.P<0.05;D,E.Detection of IgG1(D) and IgG2a(E) antibody levels in three groups.*.P<0.05 vs PBS.
间接ELISA检测小鼠血清中IgG1和IgG2a的水平,结果显示:末次免疫后F7M6组、M6组、M组小鼠血清IgG1均显著高于PBS组(P<0.001),但三组间没有统计学差异(图3D)。三组小鼠血清IgG2a水平亦均高于PBS组(P<0.001),同时,F7M6组显著高于M6组(P=0.009),但F7M6和M蛋白组间,M6和M蛋白组间差异无统计学意义(图3E)。以上结果提示:F7M6多肽疫苗可诱导高效价IgG及诱发平衡的IgG1/IgG2a(Th2细胞/Th1细胞)免疫反应。
2.4 F7M6诱导了高水平IL-4和IFN-γ的表达 取免疫后的小鼠脾细胞悬液,行ELISPTO,结果显示,IL-4水平(图4A)和IFN-γ水平(图5B)表达趋势一致:F7M6、M6和M蛋白组均显著高于PBS组(P<0.001),且F7M6均高于M6组(P<0.001)和M蛋白组(P<0.01),而M蛋白组亦高于M6组(P<0.05)。
2.5 F7M6与抗链“O”阳性血清具有高特异性结合能力 以F7M6蛋白为诊断抗原检测抗链“O”阳性的病人血清及健康儿童血清,结果显示抗链“O”阳性人血清的阳性检出率为95%,而健康儿童血清的阳性检出率为47.3%,二者差异具有统计学意义(P<0.001),见图5 。

图4 ELISPOT检测小鼠脾脏免疫细胞表达IL-4和IFN-γ水平Fig.4 Analysis IL-4(A) and IFN-γ(B) of spleen cell suspension from immunized by ELISPOTNote: *.P< 0.05 vs PBS.
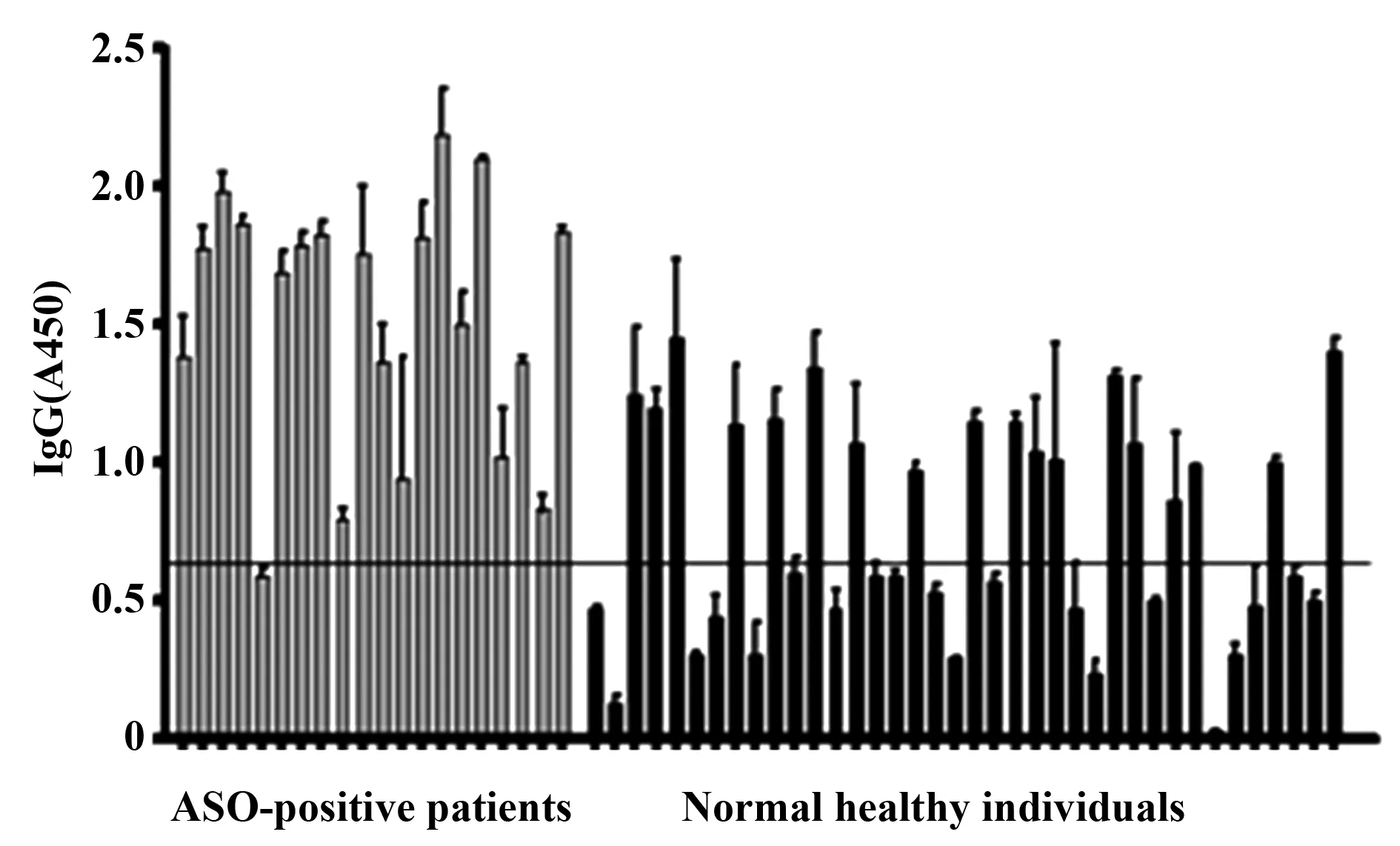
图5 ELISA分析F7M6蛋白与抗链“O”阳性血清的特异性结合能力Fig.5 Binding capacity analysis of F7M6 protein with sera from ASO-positive patients by ELISANote: The line on the column graph represents the positive threshold (OD450 =0.612).
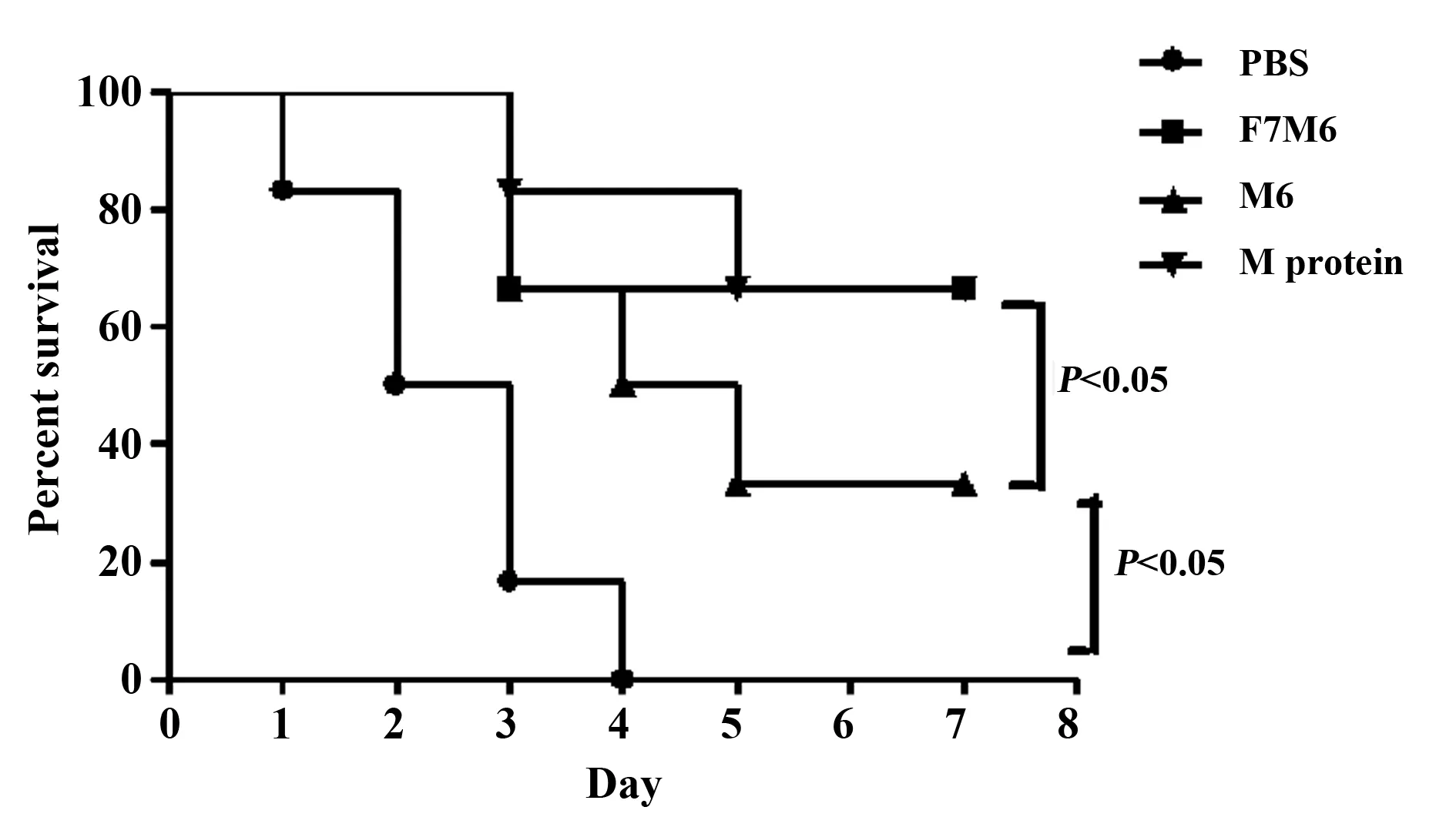
图6 M1 GAS攻毒免疫小鼠并计算其存活率Fig.6 Survival rate of immunized and control mice following challenge with M1 GASNote: Immunized and control mice were challenged with GAS bacterial suspension (equivalent to 1×108 CFU) on day 10 after the last immunization.Mice were monitored every day and calculated the survival rates.
2.6 攻毒存活率 末次免疫10 d后,小鼠腹腔攻击M1 GAS,结果显示,F7M6组和M蛋白组的存活率为66.7%,M6组为33.3%,PBS组则全部死亡(图6),统计学分析显示三组均显著高于PBS组(P<0.05),且F7M6组和M蛋白组高于M6组(P<0.05)。
3 讨论
早在上个世纪,全球范围内就对GAS疫苗进行了大规模的研究。对GAS疫苗的开发,着重于不同型别GAS表面能引起广泛保护性应答的共同分子的研究,包括C5a肽酶、外毒素B、链球菌纤维纤连蛋白结合蛋白1(Sfb1)和纤维纤连蛋白结合蛋白54(FBP54)等[13,14],但至今仍没有安全、经济、有效的疫苗问世。
对GAS疫苗研究的一个重要方向是GAS表面的M蛋白,其为GAS的主要毒力因子,也是GAS的保护性抗原,但与此同时,M蛋白与人体多种蛋白存在交叉免疫反应。M蛋白C端为高度保守区,N端为高变区A区和半可变区B区,有研究发现,M蛋白的调理性表位与自身免疫性表位是相分隔开的[15],所以对M蛋白的研究有三种方向:一是以M蛋白不同血清型N端可变区片段制备多价表位疫苗;二是以M蛋白C端保守区片段制备多价表位疫苗;三是将M蛋白融合GAS其他表面蛋白制备多价多肽融合疫苗,本研究将以上三种方法融合起来,即将与人体组织无交叉反应的可变区、C端的保守区及GAS的其他表面蛋白FbaA的有效表位串联在一起,制备出多价重组疫苗。
FbaA是近几年发现的一种新的GAS表面蛋白,它存在于近乎90%不同血清型的GAS表面,且在不同血清型中表现出较高的同源性[10,16]。有报道显示,FbaA通过结合Fn可以介导GAS对HEp-2细胞的内化[17],表明FbaA有助于GAS逃避中性粒细胞的吞噬,帮助其进入人体上皮细胞并且也具有较好的免疫原性和免疫保护性[18,19]。此外,FbaA中含有非M样蛋白,可以结合FH和FHL-1,表明FbaA可能有助于GAS逃避补体攻击。因此,FbaA蛋白也成为GAS疫苗候选蛋白。
根据我国部分地区GAS最新的流行病学调查显示,GAS感染患儿的M蛋白分型中,M12血清型所占比例几乎高达50%[8,9]。在本实验中我们将M12的保护性表位插入到本室保存的重组质粒F7M5中[4],重新构建出重组表达质粒F7M6和M6,诱导表达纯化出相应蛋白后,将F7M6蛋白、M6蛋白和本室保存的M蛋白分别免疫小鼠,小鼠血清抗体IgG水平随免疫次数逐渐增长,各实验组均能诱导高水平IgG,但以F7M6组效价升高最为明显,提示F7M6可以更有效地诱导体液免疫应答。
在本研究中,我们采用了MF59佐剂,大量研究表明,MF59作为一种可以人用的新型佐剂,其效果明显优于传统的铝佐剂[20,21]。MF59免疫原性的增强作用并不依赖于与抗原的结合,而主要是通过诱导局部免疫活化的环境,刺激并放大免疫信号,促进免疫调节细胞因子/趋化因子和免疫细胞的募集,增强树突状细胞对抗原的摄取和提呈能力,从而达到增强体液免疫应答和细胞免疫应答的作用[22]。尽管GAS有多种途径进入人体,但在本实验中F7M6多价疫苗在MF59佐剂的协助下可更好地通过诱导Th1细胞活化CTL细胞来抵抗胞内感染。IL-4促进Th细胞向Th2型分化。IL-4通过与IFN-γ共同作用,可以促进B细胞的活化,诱导IgG1的产生。加之,F7M6多价疫苗同时诱导了高效价的IgG1及IgG2,由此分析得知,F7M6可以诱导均衡的Th1/Th2免疫应答。相比之下,M6和M蛋白组只能诱导高水平的IgG1,显示F7M6疫苗优于另两组疫苗。
疫苗的研究需要综合考虑许多因素,包括人群的地理分布,疾病亚型的变异等。本研究有望为我国GAS疫苗的研制奠定基础。
[1] Olivier C.Rheumatic fever--is it still a problem? [J].J Antimicrob Chemother,2000,45 (2):13-21.
[2] Pichichero ME,Casey JR.Systematic review of factors contributing to penicillin treatment failure in Streptococcus pyogenes pharyngitis [J].Arch Otolaryngol Head Neck Surg,2008,137(6):851-857.
[3] Lucilla B,Roberta C,Simona R,etal.Therapeutic failures of antibiotics used to treat macrolide-susceptible Streptococcus pyogenes infections may be due to biofilm formation [J].J Clin Microbiol,2006,44(8):2721-2727.
[4] Ma C,Zheng L,Li W,etal.FbaA-and M protein-based multi-epitope vaccine elicits strong protective immune responses against group A streptococcus in mouse model [J].Microbes Infect,2014,16(5):409-418.
[5] 郭奕阳,马翠卿,魏 澎,等.A 族链球菌 (GAS) Fba 蛋白单克隆抗体对应表位核心氨基酸的确定 [J].中国免疫学杂志,2009,25(12):1059-1062.
[6] 马翠卿,顾海燕,郭奕阳,等.A 族链球菌表面蛋白 Fba 单克隆抗体的制备及其对应表位的初步鉴定 [J].中国免疫学杂志,2008,24(11):1046-1048.
[7] 魏 澎,马翠卿,郭奕阳,等.A 族链球菌表面蛋白 Fba 保护性区段的鉴定 [J].中华传染病杂志,2010,28(5): 257-261.
[8] 梁云梅,常贺生,沈叙庄,等.北京地区儿童携带的酿脓链球菌emm分型和超抗原speA及speC的相关性研究 [J].中华实用儿科临床杂志,2010,25(22):1700-1702.
[9] 梁云梅,常贺生,沈叙庄,等.致儿童咽扁桃体炎酿脓链球菌emm分型 [J].临床儿科杂志,2011,29(4):382-385.
[10] Terao Y,Kawabata S,Kunitomo E,etal.Fba,a novel fibronectin-binding protein from Streptococcus pyogenes,promotes bacterial entry into epithelial cells,and the fba gene is positively transcribed under the Mga regulator [J].Mol Microbiol,2001,42(1):75-86.
[11] Cui QM,Cai HL,Xiu RW,etal.Similar ability of FbaA with M protein to elicit protective immunity against group A streptococcus challenge in mice [J].Cell Mol Immunol,2009,6(1):73-77.
[12] Goodman RE.Practical and predictive bioinformatics methods for the identification of potentially cross-reactive protein matches [J].Mol Nutr Food Res,2006,50(7):655-660.
[13] Courtney HS,Hasty DL,Dale JB.Serum opacity factor (SOF) of Streptococcus pyogenes evokes antibodies that opsonize homologous and heterologous SOF-positive serotypes of group A streptococci [J].Infect Immun,2003,71(9):5097-5103.
[14] Mcarthur J ,Medina E ,Mueller A ,etal.Intranasal vaccination with streptococcal fibronectin binding protein Sfb1 fails to prevent growth and dissemination of Streptococcus pyogenes in a murine skin infection model [J].Infect Immun,2004,72(12):7342-7345.
[15] Samanidou VF,Nika MK,Papadoyannis IN.HPLC as a tool in medicinal chemistry for the monitoring of tricyclic antidepressants in biofluids [J].Mini Rev Med Chem,2008,8(3):256-275.
[16] Ramachandran V,Mcarthur JD,Behm CE,etal.Two distinct genotypes of prtF2,encoding a fibronectin binding protein,and evolution of the gene family in streptococcus pyogenes [J].J Bacteriol,2004,186(22):7601-7609.
[17] Cue D ,Dombek PE,Lam H ,etal.Streptococcus pyogenes serotype M1 encodes multiple pathways for entry into human epithelial cells [J].Infect Immun,1998,66(10):4593-4601.
[18] 李彩虹,马翠卿,王 锦,等.A族链球菌表面新发现蛋白Fba真核表达质粒的构建及其诱导的免疫应答 [J].中国免疫学杂志,2007,23(9):835-838.
[19] 马翠卿,李彩虹,王秀荣,等.A族链球菌表面蛋白Fba的原核表达及免疫原性分析 [J].中华传染病杂志,2008,26(3):146-150.
[20] Traquina P ,Morandi M ,Contorni M ,etal.MF59 adjuvant enhances the antibody response to recombinant hepatitis B surface antigen vaccine in primates [J].J Infect Dis,1996,174(6):1168-1175.
[21] Granoff DM,Mchugh YE,Raff HV,etal.MF59 adjuvant enhances antibody responses of infant baboons immunized with haemophilus influenzae Type b and Neisseria meningitidis group C oligosaccharide-CRM197 conjugate vaccine [J].Infect Immun,1997,65(5):1710-1715.
[22] O′Hagan DT,Ott GS,Gregorio ED,etal.The mechanism of action of MF59-an innately attractive adjuvant formulation [J].Vaccine,2012,30(29):4341-4348.
[收稿2016-06-08 修回2016-07-11]
(编辑 张晓舟)
Optimization and evaluation of multi-epitopes vaccine and its immune protection against to GAS infection
WANGJia-Chao,YUANTing,ZOUDong-Hua,GAOXue,GUOYi-Yang,LIJian,ZHANGZheng-Zheng,YANGLi-Juan,MACui-Qing.
DepartmentofImmunology,HebeiMedicalUniversity,KeyLaboratoryofImmuneMechanismandInterventiononSeriousDiseaseinHebeiProvince,Shijiazhuang050017,China
Objective:To optimize group A streptococcus (GAS) polyvalent epitope-based vaccine,and explore its immune protection effect.Methods: Psolyvalent vaccine of GAS,F7M6,was constructed and expressed,in which the M6 peptide was constructed as control.12 mice of each group were immunized with 20 μg per dose of purified F7M6,M6 or M protein (stored in our lab),and PBS as negative control.Half of the mice in each group were sacrificed on 10 days of the last immunization,and the levels of IgG,IgG1 and IgG2a,were detected by ELISA and the levels of IL-4 and IFN-γ were detected by ELISPOT.At the same time,the left immunized animals were challenged with a lethal dose of M1GAS,and determined the protective effect of the vaccine.Besides,the specific binding ability of F7M6 protein with the sera from ASO-positive patients was detected by ELISA.Results: In F7M6 group,the levels of not only sera IgG,IgG1,IgG2a,but also IL-4 and IFN-γ were the highest one compared with other groups.Following challenge with type M1 GAS,the survival rates of F7M6 group was 66.7%.Additionally,when F7M6 acted as diagnostic antigen,the positive detective rate of ASO-positive patient sera was 95%.Conclusion: Polypeptides F7M6,could elicit the best humoral and cell-mediated immune response than other groups,and induce a protective effect in immunized mice against M1 GAS infection.Excitedly,the epitopes of F7M6 protein covers the most of GAS serotypes at present.
Group A streptococcus(GAS);Multi-epitope vaccine;M protein;FbaA protein;Immune response
10.3969/j.issn.1000-484X.2017.03.016
①本文受国家自然科学基金 (31370914和 81172810)和河北省自然科学基金(H2016206516)资助。
王家超(1990年-),男,在读硕士,主要从事感染免疫与分子免疫研究,E-mail:865444893@qq.com。
及指导教师:马翠卿(1974年-),女,博士,教授,硕士生导师,主要从事感染免疫与分子免疫研究,E-mail:macuiqing@sina.com。
R392
A
1000-484X(2017)03-0392-06
②共同第一作者。
③河北医科大学第四医院,石家庄050017。
