基于SSR标记的水稻光温敏不育系遗传多样性分析
2017-04-05陈锦文谢旺有王天生陈惠清谢少和黄荣裕许桂芳
陈锦文,谢旺有,王天生,陈惠清,谢少和,黄荣裕,许桂芳
(福建省泉州市农业科学研究所,福建 晋江 362212)
基于SSR标记的水稻光温敏不育系遗传多样性分析
陈锦文,谢旺有,王天生,陈惠清,谢少和,黄荣裕,许桂芳
(福建省泉州市农业科学研究所,福建 晋江 362212)
采用SSR分子标记对17个水稻光温敏不育系进行遗传多样性分析。结果表明,在水稻基因组中较均匀分布的168个SSR位点中91个SSR位点具有多态性,多态性频率为54.17%。91个SSR标记共检测出229个等位基因,每个SSR位点检测到2~4个,平均2.517个等位基因,平均多态性信息含量指数(PIC)为0.412。根据17份光温敏不育系的遗传相似系数矩阵作出树状图,聚类分析结果表明17个不育系遗传相似系数变幅为0.59~0.93;以遗传相似系数0.61为阀值,可将供试不育系分为4个群,聚类分析结果与亲缘系谱分析结果基本吻合。
光温敏不育系;SSR标记;遗传多样性
水稻(OryzasativaL.)是世界上最重要的粮食作物之一,我国杂交水稻的推广应用为粮食增产做出了巨大贡献[1]。水稻作为自花授粉植物,杂交水稻的成功获益于雄性不育的利用,包括细胞质雄性不育(Cytoplasmic male sterility, CMS)和核雄性不育(Genic male sterility, GMS)。石明松于1973年在湖北省沔阳县发现了水稻变异株农垦58S[2-3]。作为天然光敏核不育水稻的新材料,农垦58S的发现开启了我国在利用水稻杂种优势培育两系杂交水稻方面的新序幕[4]。1986年,袁隆平结合Ikehashi 等人报道的水稻广亲和研究成果,率先提出“两系法”利用水稻杂种优势的战略设想,利用自然两用系,即在长日高温下制种,在短日低温下繁种,一系两用,从此拉开了大规模研究光温敏核不育水稻的序幕[5]。此后,两系法在培育杂交水稻新组合方面发挥了广泛作用,据统计,截止2010年底,全国共有427个两系组合通过审定,其中国审组合62个。目前两系杂交水稻已经成为我国稻米生产中不可或缺的类型,在保障粮食安全中发挥着极其重要的作用[4]。
杂种优势主要来源于杂交稻亲本间的遗传差异,亲本间亲缘关系越远,杂种优势越强。作为两系杂交水稻的母本,光温敏不育系对杂种优势的贡献很大,但由于光温敏不育系的雄性不育性受遗传与环境条件共同调控,因此筛选和培育优良的光温敏不育系在生产上需要花费大量的人力、物力和财力。对水稻育种而言,遗传多样性越丰富,用于改良栽培品种或者选育新品种的潜力越大[6]。而优良育种材料的获得在一定程度上取决于育种群体遗传多样性的丰富程度[7]。开展光温敏不育系遗传多样性研究,有助于选配强优势的两系杂交稻组合,更有效地利用杂种优势。
SSR分子标记由于具有灵敏度高、稳定性好、精确性和有效性高等优点,因此被广泛应用于植物基因定位和QTLs分析、DNA指纹和品种鉴定、种质资源保存和利用、系谱分析和标记辅助育种等,目前也已成为遗传多样性分析应用最为广泛的分子标记之一。伍豪等[8]利用34对SSR引物对广西生产上主要应用的25份不育系进行遗传多样性分析,共检测到112个等位基因,平均每对SSR引物可检测到的等位基因为3.2941个,Nei’s基因多样性指数平均为0.4922,平均多态信息含量(PIC)为0.4381,属于中度多态位点,据此认为广西生产上主要的不育系遗传多样性不高,亲缘关系较近,遗传背景较单一。马旭丹等[9]利用56对SSR引物对20份水稻不育系进行遗传多样性分析,共检测到191个多态性片段,平均3.41个等位基因,聚类分析结果与亲缘系谱表现出较好的一致性。方珊茹等[10]利用99对SSR引物对15个不育系进行遗传多样性分析,结果共检测到175个等位基因,平均2.47个,基因多样性指数平均为0.716,PIC平均为0.393,聚类分析结果与供试的不育系谱相吻合。欧立军[11]利用30对SSR引物对光温敏不育系628S衍生的5个形态微变异的株系进行多态性分析,共获得了4个多态性引物,多态性频率13.33%,获得了除11S之外4个株系特异性条带,由此得到其它4个株系材料是新的遗传变异的分子证据。郭慧等[12]利用135对SSR标记对24份水稻细胞质雄性不育系进行多态性分析,共获得77个等位位点,平均多态信息含量(PIC)为0.254,分析表明生产上应用的细胞质雄性不育系遗传背景比较单一,不利于充分发挥水稻的杂种优势利用潜力。
本研究应用SSR分子标记对芽变103S衍生的农艺性状差异较大的4个光温敏不育系株系与生产上应用较多的13个光温敏不育系进行了遗传多样性检测,以期为此4个株系的差异比较、配组利用及生产上应用的光温敏不育系的遗传多样性研究提供分子证据。
1 材料与方法
1.1 供试材料
选取生产上广泛应用的13个光温敏不育系及本课题组选育的4个光温敏不育系,共17份材料(表1)。
1.2 基因组DNA提取
2014年6月将17份材料种植于泉州市农科所洛阳基地,取在抽穗期经镜检败育彻底的单株叶片,采用CTAB法提取基因组DNA,用BACK MAN DU 800分光光度计检测DNA质量及浓度,将DNA浓度统一调整为50 ng/μL备用。
1.3 SSR分析
在水稻12条染色体上较均匀地选取168个SSR标记进行遗传多样性分析。选取的引物由上海生物工程有限公司合成。DNA片段扩增体系为10.0 μL,其中包括0.8 μL模板DNA (50 ng/μL),1 μL 10×PCR buffer(Mg2+Plus),0.4 μL dNTPs Mixture(各2.5 mmol/L),SSR(10 μmol/μL)正、反向引物各0.25 μL ,0.2 μL Taq酶(5 U/μL),6.5 μL ddH20 (PCR试剂购自Takara公司),在型号为GeneAmp PCR System 9700仪器上进行扩增。PCR反应程序为:94 ℃预变性5 min;94 ℃变性30 s,55 ℃退火30 s,72 ℃延伸2 min,35个循环;最后72 ℃延伸7 min,4 ℃保存备用。PCR产物在8%浓度的非变性聚丙烯酰胺凝胶上以90 V恒压电泳120 min,用荧光染料Genefinder(购自厦门百维信生物科技有限公司)在摇床上染色20 min,最后在BIO-RAD凝胶成像系统上拍照保存,记录电泳结果。
1.4 数据分析
一对SSR引物检测单个位点,每个多态性条带视为1个等位基因,参考网站http://www.gramene.org/中SSR标记信息记录扩增片段的长度。根据引物扩增结果,选取扩增条带清晰且具有多态性的SSR引物进行统计分析。根据扩增片段大小,建立1、0数据库,在相同迁移位置上,有条带的记为1,无条带的记为0,缺失条带的记为9。按照DNA分子标记数据分析方法[13],将SSR标记作为等位基因进行多态性分析,每扩增得到的一条带记为一个性状。利用NTSYS 2.10数据分析软件进行SHAN聚类分析,并绘制树状聚类图。0~1数据采用DataTrans 1.0 程序转化后[14],应用Popgene 3.2计算等位基因数(Na)、有效等位基因数(Ne)、Shannon’s信息指数(I)和平均多态性信息量指数(polymorphism information content,PIC)。
2 结果与分析
2.1 SSR标记的多态性
选用水稻12条染色体上均匀分布的168个SSR标记对供试光温敏不育系进行多态性分析,共筛选出多态性标记91个,多态性频率为54.17%。从表2中可看出,91个多态性标记共检测到229个等位基因。不同标记所检测到的等位基因数量不同,变异范围为2~4个,平均为2.517个。其中RM6324、RM85、RM1359、RM473E、RM258、RM224和RM1261等7个标记的等位变异较为丰富,均能检测到4个等位基因;其他大部分标记均能检测到2~3个等位基因。Shannon’s信息指数I平均为0.671,变幅为0.224~1.320;平均多态性信息含量指数PIC为0.412,变幅为0.111~0.718;PIC值高于平均值的多态性位点数是52个,占57.14%;其中RM258的PIC最高,遗传多样性最丰富。图1为RM366对供试不育系的多态性检测结果。
2.2 聚类分析
基于91个多态性标记所获得的数据,应用NTSYS软件计算出供试光温敏不育系间的遗传距离为0.079~0.507,平均为0.376。根据遗传距离采用UPGMA法进行聚类(图2),结果表明供试光温敏不育系在阀值为0.59处聚为两大类群,一类为培矮64S、Y58S、1892S、Y58-2S及未知系谱来源的潭S,彼此间的遗传距离为0.157~0.454,平均为0.321,其中培矮64S、Y58S、1892S、Y58-2S均由农垦58S或其衍生系选育出来的,系谱来源未知的光温敏不育系潭S聚在此群里。另一大类群包括本研究选用的其他光温敏不育系。在阈值0.61处分为4个亚群,272S单独分为一个亚群,与其它光温敏不育系间的遗传距离为0.341~0.476,平均为0.412,表现出较大的遗传差异;株1S、陆18S、品57S、明软02S及明糯S-1聚为一个亚群,彼此间的遗传距离为0.205~0.393,平均为0.334;糯性光温敏不育系明糯S-1单独聚为一个亚群;广占63-4S、RGD-7S及本课题组选育的4个光温敏不育系聚为一个亚群。在阈值0.76处可将1202S与1211S、1215S及1217S区别,彼此间的遗传距离为0.079~0.244,平均为0.173,这4个株系是由芽变103S选育出来的;同一系谱的株系1211S、1215S在阀值0.92处可以区别,遗传距离为0.079。

1:Y58S;2:明软02S;3:广占63-4S;4:株1S;5:陆18S;6:272S;7:品57S;8:1202S;9:1211S;10:1215S;11:培矮64S;12:RGD-7S;13:1217S;14:Y58-2S;15:明糯S-1;16:1892S;17:潭S;M:DL100。
图1 RM366对17个样本的多态性分析
3 结论与讨论
随着分子标记技术的不断发展,基于PCR技术的RAPD、AFLP、RFLP及SSR等分子标记被广泛用于遗传多样性研究,前人[15-17]通过比较研究4种分子标记,认为SSR的多态性信息量最多。SSR是共显性标记,因具有多态性丰富、操作简单、检验快速、重复性好等优点,被广泛应用于遗传多样性分析。
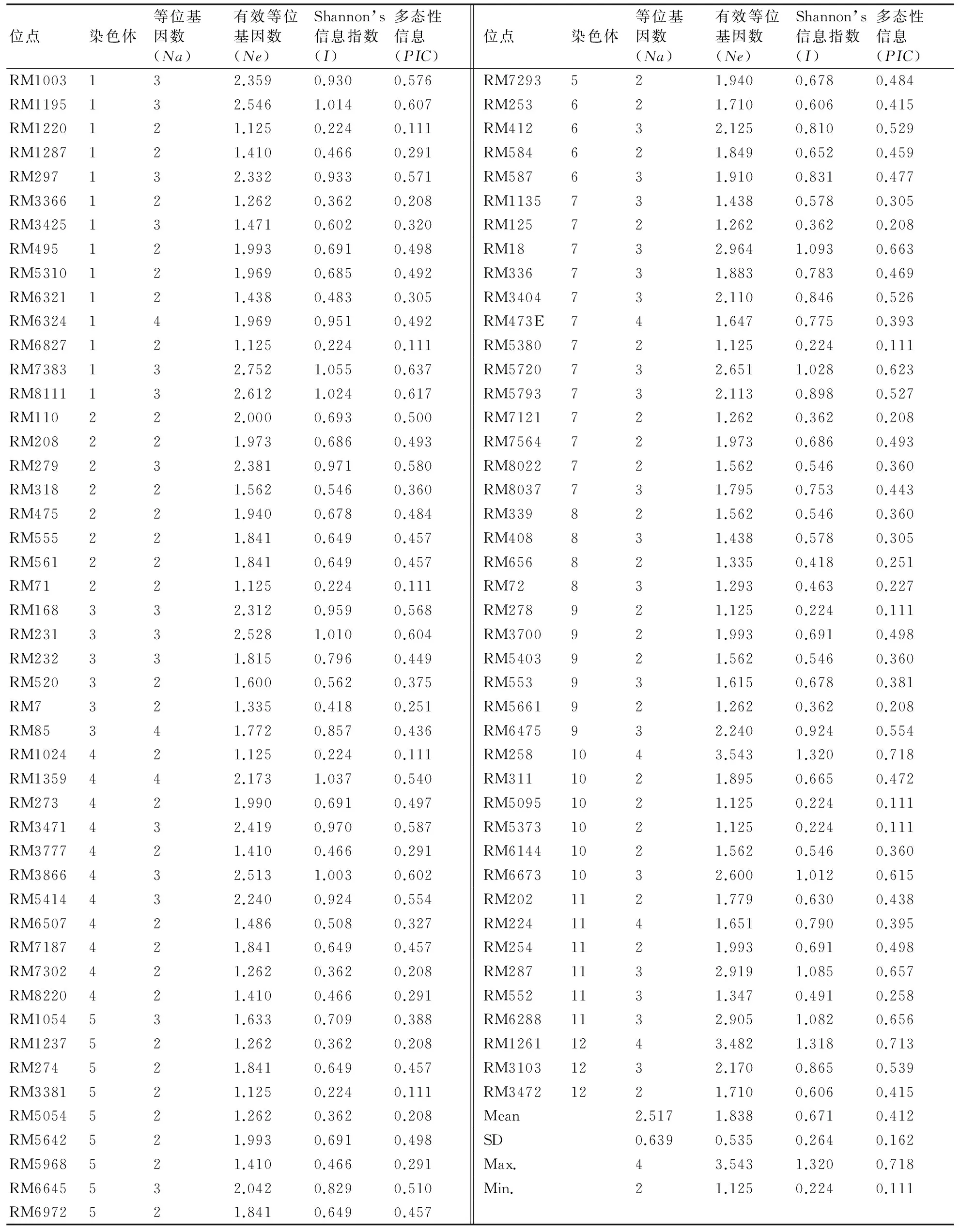
表2 91个SSR标记在供试不育系中的遗传多样性信息
本研究采用SSR标记将供试光温敏不育系分为两大类群4个亚群。272S单独聚为一个亚群,它是以金山S-2为不育基因供体,用保持系金山1B为父本,通过金山S-2与金山1B杂交后再用金山1B回交1次,选育获得的性状优良的两系不育系[18],与同样源于金山S-2的品57S间的遗传距离为0.393,品57S与272S植株的农艺性状差异较大,说明在杂交配组系谱筛选分离世代时,筛选得到了差异较大的两个光温敏不育系品种(系)。其它光温敏不育系的遗传差异较大。农垦58S及其衍生的光温敏不育系聚为一个群,这与马旭丹等[9]的研究结论一致,农垦58S类群通过聚类将未知系谱来源的潭S聚在农垦58S衍生选育的一个类群中,推测潭S为农垦58S或其衍生系选育而来。株1S、陆18S、品57S、明软02S及明糯S-1聚为一个群,从系谱分析结果看,株1S与陆18S均是以抗罗早为母本选育而来[19],具有相似的遗传背景,彼此间的遗传距离为0.203,聚类分析结果与系谱来源相吻合;来源于三明市农科院的光身糯稻光温敏不育系明糯S-1是以SE21S为母本,双光S为父本选育而来[20],与明软02S具有较近的遗传背景,彼此间遗传距离为0.389,明糯S-1的糯稻兼光身特性与其他几个光温敏不育系差异较大,在聚类中单独聚为一个亚群。广占63-4S、RGD-7S及本课题组选育的4个光温敏不育系聚为一个群,广占63-4S与RGD-7S的遗传距离为0.419,广占63-4S是N422S/广占63经系统选育而成,RGD-7S是以测64S为母本、312/广恢3550为父本选育成的RGD-7S与BL122杂交回交选育而成[19],1202S、1211S、1215S及1217S均以芽变103S为母本选育而来,1217S是从黑米不育系1215S分离出来的株系,4个株系的聚类结果与系谱相吻合;聚类分析结果表明本课题组选育的4个光温敏不育系与生产上大面积应用推广的广占63-4S及RGD-7S亲缘关系更近。
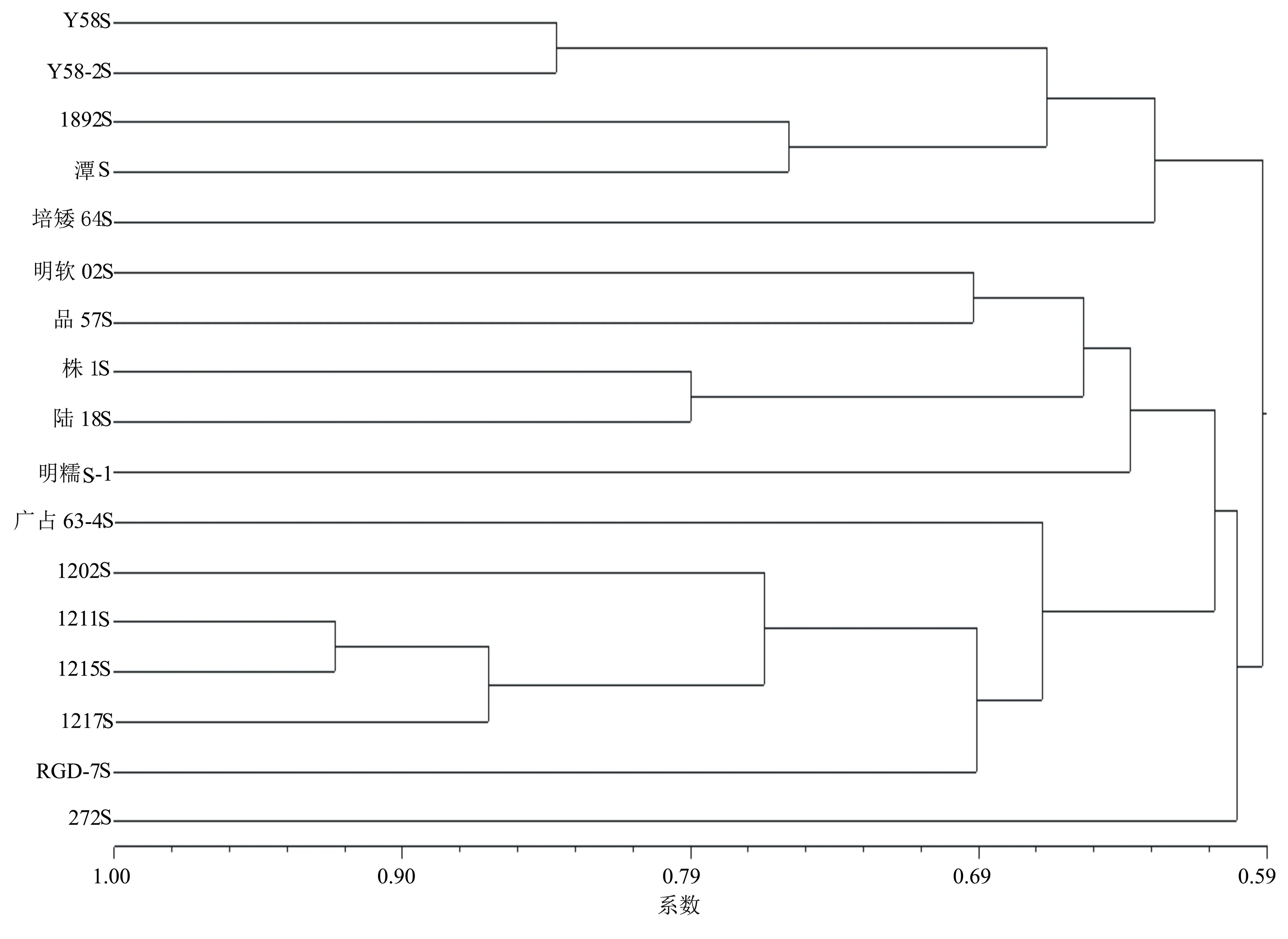
图2 17份光温敏不育系的SSR聚类分析
传统系谱分析法常缺乏育种家选种过程中产生的选择和突变效应,所以仅仅用系谱法较难把握一些材料的亲缘关系,而分子标记技术可以准确检测到材料中DNA水平上的遗传变异,能够更好地为材料利用提供指导[21]。田间农艺性状观察、系谱分析及分子标记技术相结合应用,可对现行资源的充分开发利用及其对新的种质资源的综合利用提供良好的指导。利用SSR分子标记对水稻光温敏不育系进行多样性研究,可为两系杂交稻的遗传改良和亲本配组提供理论依据,对提高分子育种水平及光温敏不育系的挖掘利用有着积极的推动作用。笔者利用上述基于分子标记鉴定存在差异位点的4个光温敏不育系,正加紧进一步进行杂交配组,以期选育出新的强优势两系杂交稻新组合。
[1] 程式华,李建.现代中国水稻[M].北京:金盾出版社,2007.
[2] 石明松.晚粳自然两用系选育及应用初报[J].湖北农业科学,1981(7):3.
[3] 石明松.对光照长度敏感的隐性雄性不育水稻的发现与初步研究[J].中国农业科学,1985(2):44-48.
[4] 斯华敏,刘文真,付亚萍,等.我国两系杂交水稻发展的现状和建议[J].中国水稻科学,2011,25(5):544-552.
[5] 袁隆平.杂交水稻超高产育种战略设想[J].杂交水稻,1986(2):3-4.
[6] 赵一洲,李正茂,路洪彪,等.辽宁省水稻骨干亲本演变及遗传多样性分析[J].河南农业科学,2014,43(12):28-33.
[7] 张继峰,刘玉梅,方智远,等.青花菜相同亲本的DH与F2群体遗传多样性的比较[J].园艺学报,2012,39(6):1090-1098.
[8] 伍豪,刘百龙,王威豪,等.广西生产上主要应用的三系杂交稻不育系遗传多样性分析[J].南方农业学报,2015,45(4):550-554.
[9] 马旭丹,孙辽,李元元,等.部分水稻不育系的指纹图谱构建和遗传多样性分析[J].杂交水稻, 2015(6):64-70.
[10] 方珊茹,郑燕梅,吴春珠,等.基于SSR标记的杂交籼稻主要不育系遗传多样性分析[J].福建农业学报,2012,27(11):1173-1177.
[11] 欧立军.微卫星(SSR)分子标记在杂交水稻不育系不同株系遗传差异分析中的应用[J].江苏农业科学,2009,37(5):45-46.
[12] 郭慧,李树杏,徐建第,等.24份水稻细胞质雄性不育系的SSR多态性分析[J].杂交水稻,2007,22(3):56-61.
[13] 周延清.DNA分子标记技术在植物研究中的应用[M].北京:化学工业出版社,2005:249-252.
[14] 盖红梅,任民.SSR数据处理宏程序DataTrans 1.0[J].分子植物育种:网络版,2011(9):1359-1365.
[15] Pejic I, Ajmone-Marsan P, Morgante M, et al. Comparative analysis of genetic similarity among maize inbred lines detected by RFLPs, RAPDs, SSRs, and AFLPs[J]. Theoretical and Applied Genetics, 1998, 97: 1248-1255.
[16] 袁力行,傅骏骅,Warburton M, et al.利用RFLP、SSR、AFLP和RAPD标记分析玉米自交系遗传多样性的比较研究[J].遗传学报,2000,27(8):725-733.
[17] Belaj A, Satovic Z, Cipriani G, et al. Comparative study of the discriminating capacity of RAPD,AFLP and SSR markers and of their effectiveness in establishing genetic relationships in olive[J]. Theoretical and Applied Genetics, 2003, 107(4): 736-744.
[18] 吴建梅,林荔辉,林培清,等.优质水稻光温敏核不育系272S的选育[J].杂交水稻,2012,27(1):11-15.
[19] 斯华敏,付亚萍,刘文真,等.水稻光温敏雄性核不育系的系谱分析[J].作物学报,2012,38(3):394-407.
[20] 黄显波,邓则秦,郑家团,等.光身糯稻两系不育系明糯S-1选育研究[J].福建稻麦科技,2006,24(1):1-2.
[21] 王胜军,陆作楣,万建民.采用表型和分子标记聚类研究杂交籼稻亲本的遗传多样性[J].中国水稻科学,2006,20(5):475-480.
(责任编辑:许晶晶)
Genetic Diversity Analysis of Rice PTGMS Lines Based on SSR Markers
CHEN Jin-wen, XIE Wang-you, WANG Tian-sheng, CHEN Hui-qing,XIE Shao-he, HUANG Rong-yu, XU Gui-fang
(Quanzhou Institute of Agricultural Sciences in Fujian Province, Jinjiang 362212, China)
The genetic diversities of 17 rice PTGMS lines were analyzed by adopting 168 pairs of SSR markers which were evenly distributed on the 12 chromosomes of rice. Among these 168 pairs of SSR markers, 91 pairs of SSR markers showed a polymorphism with the polymorphic frequency of 54.17%. A total of 229 alleles were detected with these 91 polymorphic SSR markers, and the number of alleles per SSR locus ranged from 2 to 4 with an average of 2.517. The average polymorphic information content index was 0.412. According to the genetic similarity coefficient matrix of the tested 17 rice PTGMS lines, a dendrogram was constructed by UPMGA, and a clustering analysis was conducted, the results showed that the range of genetic similarity coefficient of 17 rice PTGMS lines was 0.59~0.93. When genetic similarity coefficient 0.61 was used as a threshold, the tested rice PTGMS lines could be divided into four groups, which was basically consistent with their pedigree relationship.
PTGMS line; SSR marker; Genetic diversity
2016-11-14
泉州市优秀人才培养专项(14A27);福建省泉州市科技项目(2014N29、2015N27);福建省星火项目(2016S0015)。
陈锦文(1984─),男,福建德化人,助理研究员,硕士研究生,主要从事水稻抗病育种研究。
S511
A
1001-8581(2017)03-0001-06
