短花针茅生长和繁殖策略对载畜率的响应
2017-04-01古琛赵天启王亚婷王玺贾丽欣赵萌莉
古琛,赵天启,王亚婷,王玺,贾丽欣,赵萌莉
内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010019
短花针茅生长和繁殖策略对载畜率的响应
古琛,赵天启,王亚婷,王玺,贾丽欣,赵萌莉*
内蒙古农业大学草原与资源环境学院,内蒙古 呼和浩特 010019
生长和繁殖是植物的两个最基本的生命过程,二者之间的资源分配是植物生活史策略的核心问题。为了响应家畜放牧的干扰,草原植物需要不断地在营养生长和生殖生长之间进行调节。试验以内蒙古荒漠草原为研究对象,采用4个载畜率水平完全随机区组试验设计,研究荒漠草原建群种短花针茅(Stipa breviflora)生长和繁殖策略对不同载畜率的响应,对揭示短花针茅应对放牧干扰的响应策略有重要的生态学意义,也可为荒漠草原生态系统的保护和放牧利用提供参考。结果表明:(1)短花针茅种群盖度在轻度放牧区显著增大,在重度放牧区显著降低(P<0.000 1),其种群和株丛地上生物量、高度均随载畜率的增大而显著降低(P<0.000 1),但密度随载畜率的增大而增大(P<0.000 1);(2)轻度放牧区短花针茅的分蘖数(P=0.048)和株丛径(P=0.023)显著高于其他处理,且分蘖节深度随着载畜率的增大而显著变浅(P=0.042);(3)轻度放牧显著增加了短花针茅的种子产量,重度放牧使之显著降低(P<0.000 1),且种子生物量占植株地上生物量的比例在轻度放牧区最高(P<0.000 1);(4)短花针茅总分枝数和生殖枝数在轻度放牧区最多(P<0.000 1),随着载畜率的增大而显著减少,重度放牧显著降低了生殖枝所占比例而显著提高了营养枝的比例(P<0.000 1)。短花针茅的生长、分蘖(无性繁殖)和有性生殖在轻度放牧区均显著升高,而随着载畜率的增大而显著降低,说明轻度放牧(载畜率为0.91sheep·hm-2·0.5 a-1)是短花针茅种群的最适载畜率,也是其生长和繁殖的载畜率阈值。
荒漠草原;放牧强度;种群特征;分蘖;有性生殖
生长和繁殖是植物的两个最基本的生命过程,生物量在生长和繁殖之间的分配是植物生活史策略(life-history strategy)的核心问题(Weiner et al.,2009)。生活史理论(life-history theory)认为植物在生长和繁殖之间的资源分配具有很强的灵活性(Miller et al.,2006)。繁殖是植物完成一个生命循环、繁衍后代并影响植物种群动态、保证群落稳定、维持草地生物多样性和生态系统健康的关键(Wang et al.,2015)。植物的繁殖方式主要有有性生殖和营养繁殖两种:有性繁殖有利于植物对当地环境的适应以及基因的进化,可增加植物的遗传多样性;而营养繁殖幼苗能够依靠母体提供营养,存活率高,有利于增加植物对环境的适合度(fitness)(Ikegami et al.,2007;Neves et al.,2014;侯牡丹等,2009)。但植物的有性生殖对气候和环境的变化极为敏感(Moreira et al.,2015;张勃等,2015),而无性繁殖是维持种群结构稳定的主要方式(Dong et al.,2015)。这对于气候波动较大的草原尤其是荒漠草原的植物是极为重要的适应方式(Pluess et al.,2005)。
荒漠草原是欧亚大陆草原的重要组成部分,是草原向荒漠区过渡旱生性最强的草地生态系统(卫智军等,2016)。因气候干燥、生境严酷,植被低矮稀疏,群落结构简单,生态系统十分脆弱(王晨晨等,2014),对自然和人类活动的干扰较为敏感,与其他草原类型相比稳定性较差(潘占磊等,2016)。放牧是草地生态系统中最重要的人为干扰因素之一,也是荒漠草原最主要、最直接、最经济的利用方式(古琛等,2015)。为了响应家畜放牧的干扰,草原植物需要不断地在无性繁殖和有性生殖之间进行调节。植物对无性繁殖投入资源的增加必将导致其对有性生殖资源投入的减少(Pluess etal.,2005)。丛生禾草的种群由植株和分蘖两部分组成,放牧对其繁殖的调节具体体现在种群、株丛和分蘖水平上(Butler et al.,1988;张晓娜等,2010)。
许多学者针对不同放牧强度对短花针茅(Stipa breviflora)荒漠草原土壤养分(孙世贤等,2013)、土壤理化性质(Li et al.,2008)、土壤呼吸(张新杰等,2014)、植被空间格局和土壤养分(Lin et al.,2010a;2010b)、植物多样性和草地生产力(韩国栋等,2007)、植物不同功能群生物量(古琛等,2015)、物种多度格局(杜宇凡等,2016)、植被的空间异质性(黄琛等,2014)、短花针茅叶片功能性状(刘文亭等,2016)等进行了大量的研究。短花针茅作为荒漠草原的建群种,其繁殖方式对于草原的发展方向具有举足轻重的作用。研究其在不同载畜率下的生长和繁殖策略,对揭示丛生禾草应对放牧干扰的响应策略有重要的生态学意义。
为此,本试验以内蒙古荒漠草原建群种短花针茅种群为研究对象,分析不同载畜率对短花针茅生长的影响及对无性繁殖和有性繁殖过程的调节,旨在探讨以下4个问题,(1)不同载畜率如何影响短花针茅的群落特征?(2)短花针茅的分蘖特征是否受载畜率的影响?如何变化?(3)短花针茅的繁殖方式在不同载畜率下如何变化?(4)不同放牧压力下短花针茅有性生殖和无性繁殖哪个占主导?通过对以上问题的回答,试图揭示不同放牧强度对荒漠草原建群种短花针茅的生长和繁殖规律的影响,以期为荒漠草原生态系统的保护和放牧利用提供参考。
1材料与方法
1.1试验地概况
试验区位于阴山北麓,内蒙古自治区乌兰察布市西北部的葛根塔拉草原,四子王旗王府一队,内蒙古农牧科学院草原研究所荒漠草原实验基地(41°47′17″N,111°53′46″E)。地势平坦开阔,海拔1450 m。
该旗地处中温带大陆性季风气候区,具有干旱、少雨、多风和蒸发量大的气候特征。年均气温3.4 ℃,月平均温度最高月为6—8月,年均气温分别为21.5、24.0和23.5 ℃,≥0 ℃积温为2500~3100 ℃,≥10 ℃的年积温为2200~2500 ℃。无霜期90~115 d;平均日照时数为3117.7 h。年均降水量299.4 mm,且年际变化较大,降水主要集中在5—9月,这段时间的降水量占全年总降水总量的80%以上。年潜在蒸发量约2300 mm,远超过降水量,是降水量的7~10倍,高蒸发量致使空气干燥,干旱严重,湿润度仅为0.15~0.30。
草地植被类型为短花针茅+冷蒿(Artemisia frigida)+无芒隐子草(Cleistogenes songorica)。草层低矮,平均高度为8 cm,盖度为15%~25%,植被较稀疏。植物群落由20多种植物组成,主要为多年生草本短花针茅、无芒隐子草、银灰旋花(Convolvulus ammannii)、阿尔泰狗娃花(Heteropappus altaicus)、冰草(Agropyron cristatum)、羊草(Leymus chinensis)、细叶葱(Allium tenuissimum)、蒙古葱(Allium mongolicum)等;一、二年生草本有猪毛菜(Salsola collina)、猪毛蒿(Artemisia scoparia)、栉叶蒿(Artemisia pectinata)等。还有灌木小叶锦鸡儿(Caragana microphylla)、矮灌木狭叶锦鸡儿(C. stenophylla)、半灌木驼绒藜(Ceratoides latens)、小半灌木木地肤(Kochia prostrata)、冷蒿等。
土壤为淡栗钙土,土层厚度大约为1 m,在40~50 cm深处有钙积层,土体坚硬且渗透能力差,有机质含量较低。土壤微生物有好气性细菌、放线菌和霉菌等。
1.2试验设计和测定方法
1.2.1样地概况
放牧试验于2003年6月开始,在内蒙古乌兰察布市四子王旗短花针茅草原上,选取地势相对平坦、植被均一的约50 hm2天然草地围封,进行不同载畜率放牧试验。采用完全随机区组设计,将围封的草地分为12个小区,3个区组(3次重复),每个区组中设4个处理(4个载畜率水平)(图1),分别为对照(CK)、轻度放牧(Light grazing,LG)、中度放牧(Moderate grazing,MG)和重度放牧(Heavy grazing,HG)。载畜率值依次为0、0.91、1.82和2.71 sheep∙hm-2∙0.5 a-1,绵羊只数分别为0、4(轻度)、8(中度)和12(重度)只。每个区组内的4个处理完全随机排列,每个处理的面积约为4.4 hm-2。放牧种为当地成年蒙古羯羊。从2003—2016年每年放牧期为半年,6月1日开始,11月30日结束。每日早晨6点将绵羊赶入各小区让其自由采食,下午6点赶回圈内饮水、休息。
图1 试验小区和区组示意图Fig. 1 Schematic diagram for experimental plot and block
1.2.2测定方法
2016年放牧开始前,在各小区分别围封10个1 m×1 m样方以测定群落特征。短花针茅种群生长和繁殖野外采样于6月中旬进行。在已围封好的样方内测定短花针茅种群的高度、盖度、密度、株丛径和生物量,其中,高度为卷尺测定3次自然高度的均值,盖度为目测法测定的分盖度,密度是单位面积内短花针茅的物种数,株丛径为卷尺测定植株的平均直径。再在每个样方内随机选取1丛完整株丛单独测定其地上生物量、总分枝数、生殖枝数、营养枝数、种子重量、分蘖数和分蘖深度等指标,每小区10次重复。将所取样品带回实验室后置于鼓风干燥烘箱中于65 ℃烘干24 h后称其干质量。
种子资源分配比例=种子生物量/植株地上生物量×100%
1.2.3数据分析方法
试验数据采用Excel 2013软件进行整理、简单分析和图表制作,不同处理间短花针茅种群特征、株丛结构等的差异性采用SAS.9.4在α=0.05显著性水平下进行单因素方差分析(One-way ANOVA)。
2结果与分析
2.1载畜率对短花针茅种群特征的影响
短花针茅种群的盖度在轻度放牧区最高,比对照区高57.1%;重度放牧区最低,比对照区低47.3%(P<0.0001);中度放牧区与对照区相比差异不显著(图2a)。
短花针茅种群密度随载畜率的增大而逐渐增大,重度放牧区最大,为56.9 count∙m-2;中度放牧区和轻度放牧区次之,分别为32.4和28.1 count∙m-2;对照区最小,为21.5 count∙m-2。除中度和轻度放牧区之间差异不显著外,其他处理间差异均显著(P<0.0001)(图2b)。
短花针茅种群高度随载畜率的增大而显著降低,依次为对照区17.0 cm,轻度放牧区14.5 cm,中度放牧区11.1 cm和重度放牧区7.1 cm,且各处理间差异均显著(P<0.0001)(图2c)。
短花针茅种群生物量也表现出随载畜率的增大而降低的趋势,对照区与轻度放牧区最高,分别为42.0 g∙m-2和49.1 g∙m-2;中度放牧区次之,为25.5 g∙m-2;重度放牧区最低,为12.6 g∙m-2。除对照与轻度放牧区之间差异不显著外,其他处理间差异显著(P<0.0001)(图2d)。
2.2载畜率对短花针茅株丛径的影响
随载畜率的增大,短花针茅株丛径呈先增大后减小的趋势。轻度放牧区的株丛径最大,为5.2 cm;中度与重度放牧区次之,分别为4.1 cm和3.9 cm;对照区最小,为2.7 cm。除中度与重度放牧区之间差异不显著外,其他处理间差异显著(P=0.023)(图3)。
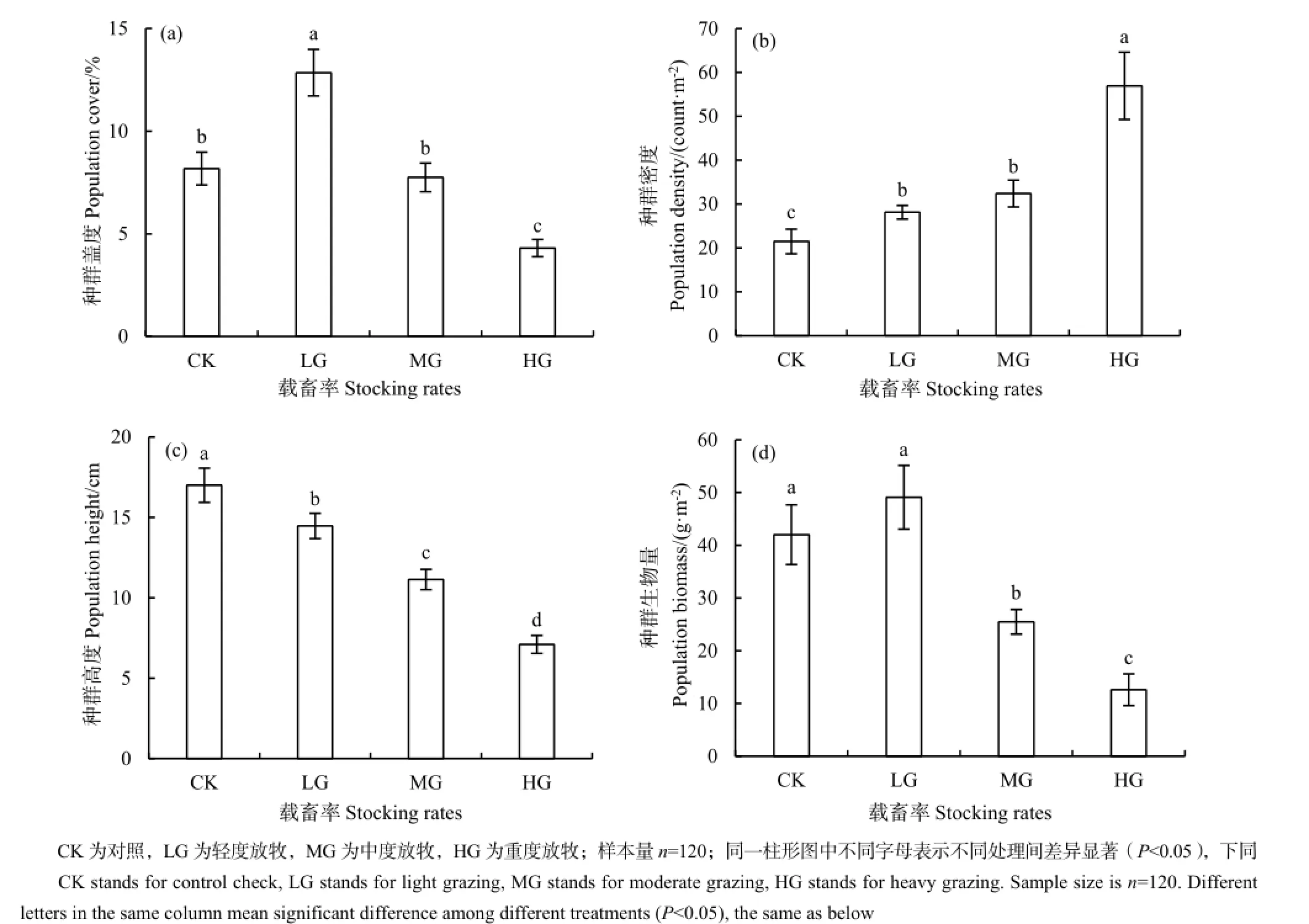
图2 不同载畜率短花针茅的种群特征Fig. 2 Population characteristics of Stipa breviflora under different stocking rates
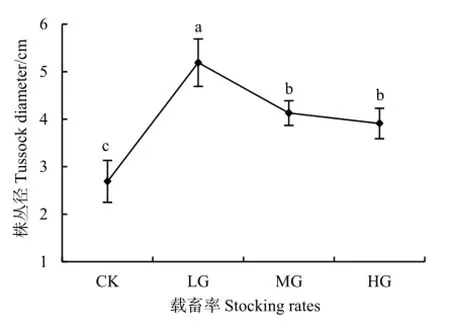
图3 不同载畜率短花针茅株丛径Fig. 3 The diameter of S. breviflora tussock under different stocking rates
2.3载畜率对短花针茅分蘖数量和深度的影响
短花针茅分蘖深度随载畜率的增大而显著降低,从高到低分别是对照区2 cm,轻度放牧区1.7 cm,中度放牧区1.5 cm和重度放牧区1.1 cm,各处理间差异均显著(P=0.042)(图4a)。轻度放牧显著增加了短花针茅的分蘖数,与其他处理相比差异显著(P=0.048),其余各处理间差异不显著(图4b)。
2.4载畜率对短花针茅株丛地上生物量与种子产量的影响
短花针茅株丛地上生物量随载畜率的增大而降低,对照区与轻度放牧区最高,分别为2.7 g∙tossock-1和2.5 g∙tossock-1;中度放牧区次之,为1.7 g∙tossock-1;重度放牧区最小,为1 g∙tossock-1。除对照区与轻度放牧区间差异不显著外,其他处理间差异显著(P<0.0001)(图5a)。
与对照区相比,短花针茅的种子产量在轻度放牧区显著增加了49.0%,在重度放牧区显著减少了60.8%(P<0.0001),中度放牧区差异不显著(图5b)。种子产量占地上生物量的比例随载畜率的增大呈先增大后减小的趋势,轻度与中度放牧区显著高于对照与重度放牧区(P<0.0001)(图6)。
2.5载畜率对短花针茅株丛结构的影响
载畜率对短花针茅的总分枝数和生殖枝数的影响规律一致,二者均随载畜率的增大而降低。重度放牧显著降低了总分枝数和生殖枝数,二者在轻度和中度放牧区与对照区相比差异不显著,但在轻度放牧区显著高于中度放牧区(P<0.0001)(图7a)。

图5 不同载畜率短花针茅地上生物量与种子产量Fig. 5 Aboveground biomass and seed yield of S. breviflora under different stocking rates
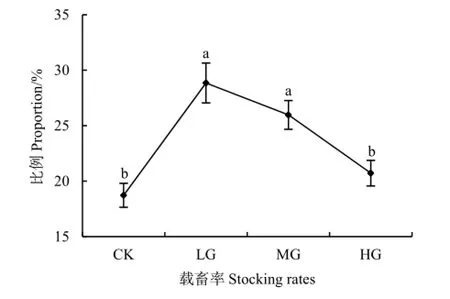
图6 不同载畜率短花针茅种子资源分配比例Fig. 6 The proportion for resource allocating in seed of S. breviflora under different stocking rates
短花针茅营养枝数在轻度放牧区和重度放牧区与对照区比差异不显著,重度放牧区的营养枝数显著多于轻度放牧区,中度放牧区与对照区相比,短花针茅营养枝数显著减少(P=0.013)(图7a)。
随着载畜率的增大,短花针茅生殖枝所占比例呈先增大后减少趋势,营养枝所占比例则正好相反。与对照区相比,中度放牧使短花针茅生殖枝比例显著增加,营养枝比例显著减少;重度放牧使短花针茅生殖枝比例显著减少,营养枝比例显著增大;轻度放牧区与对照区相比,营养枝和生殖枝比例差异不显著(P<0.0001)(图7b)。
3讨论
3.1 载畜率对短花针茅种群特征的影响
放牧是天然草原重要的影响因子和进化驱动力,家畜放牧对草地生态系统的影响是控制草地群落特征的主导因子(宋磊等,2016;李永宏,1988)。本试验结果表明,短花针茅种群盖度在轻度放牧区显著增大,重度放牧区显著降低(图2a);高度随载畜率的增大而显著降低(图2c);地上生物量随载畜率的增大而减少(图2d)。这与宋磊等(2016)、乔丽青等(2014)、侯牡丹等(2009)的研究结论一致,生物量、高度、盖度三者之间相辅相成,随载畜率变化的趋势基本一致。长期处于放牧压力下,植物出现明显矮小化,导致生物量减少、盖度降低(李西良等,2015)。这是因为植物通过降低自身高度,达到躲避家畜采食的目的,是草原植物响应动物采食的重要策略。再者,随着载畜率的增大,家畜的过度采食和践踏对短花针茅的破坏加剧,使其光合面积减少,再生能力下降,光合产物的积累降低,从而导致生物量减少,盖度降低(任海彦等,2009;段敏杰等,2010)。本试验还发现短花针茅种群的密度随载畜率的增大而增大(图2b),这主要是因为放牧践踏使短花针茅株丛破碎化,随着放牧强度的增大,大株丛的比例减少,中小株丛的比例增大,由原来的大株丛形成若干个小的株丛,后脱离母株通过“分丛”进行营养繁殖,使种群密度增大(白永飞等,1999;杜利霞等,2008;侯牡丹等,2009)。
3.2 载畜率对短花针茅分蘖数和深度的影响
分蘖是短花针茅无性繁殖的主要方式,分蘖的多少直接影响种群个体的数量。侯牡丹等(2009)指出轻度放牧下植株的分蘖数最多,这与本试验得出的轻度放牧显著提高了短花针茅分蘖数(图4b)的结果一致。对照区因为没有家畜采食的刺激,分蘖活动处于抑制状态;中度和重度放牧区分蘖数少则是因为载畜率过高导致家畜的采食活动增强,株丛的生活力减弱,生长受到抑制;轻度放牧刺激了短花针茅的生长,促进其分蘖,使其有较高的分蘖数(侯牡丹等,2009)。而且轻度放牧显著增大了短花针茅的株丛径(图3),株丛径越大,分蘖数越多。
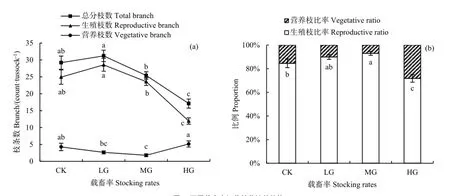
图7 不同载畜率短花针茅株丛结构Fig. 7 Tussock structure of S. breviflora under different stocking rates
本试验还得出,短花针茅分蘖节深度随载畜率的增大而显著降低(图4a),分蘖节分布深度逐渐变浅,主要是因为随着载畜率的增大,放牧家畜过度践踏,使土壤的紧实度和容重增大。通透性变差,孔隙度降低(安慧等,2013;戎郁萍等,2001),从而引起短花针茅株丛分蘖节向地表移动,更有利于分蘖的出土。体现了短花针茅对土壤环境改变的主动响应(白永飞等,1999)。
3.3 载畜率对短花针茅繁殖策略的影响
繁殖策略是植物在其生活史繁殖过程中对外界干扰的响应(刘志民等,2003),是资源向生存、生长和繁殖活动中最优分配的结果。植物在不同生境中的繁殖对策反映了其对环境的适应能力和生殖的潜能。绝大多数草地植物都可以同时采取有性和无性繁殖方式来实现种群繁衍和扩张。普遍认为,植物在扰动大、竞争相对较弱的生境中,倾向于有性繁殖;在扰动小、相对稳定的生境中,倾向于无性繁殖(Wu et al.,2010;张勃等,2015)。
生殖分配过程对放牧干扰非常敏感,草原植物在面对放牧干扰时会权衡它们的繁殖策略,这可能是植物在长期的进化过程中形成的应对放牧干扰的策略(乔丽青等,2014)。大量研究表明,过度放牧会导致植物有性生殖能力减弱。过度放牧导致大针茅生殖枝数和种子数大量减少,种子的千粒重和萌发力下降,从而抑制了大针茅的有性生殖(蒙荣等,2004)。放牧使生殖枝数和花朵数等生殖构件数量减少,单株种子的数量和产量下降,生物量向籽粒分配的比例降低(乔丽青等,2014)。放牧抑制了羊草种群的有性生殖,种群水平种子的资源分配随放牧强度增加而显著降低,在重度放牧条件下几乎没有分化生殖枝(王仁忠,2000)。随着载畜率的增大,冷蒿有性繁殖分配减少,生殖构件显著下降,生殖枝数和种子产量降低,甚至在重度放牧中有性生殖几乎消失(杜利霞等,2007)。
本试验得出的结论与以上研究结果基本一致,随着载畜率的增大短花针茅的有性生殖能力下降。短花针茅的总分枝数和生殖枝数在轻度放牧水平下呈上升趋势,而后随着载畜率的增大而降低(图7a)。这与白永飞等(1999)研究得出的禁牧和轻度放牧区克氏针茅(Stipa krylovii)株丛的总枝条数和生殖枝数均显著高于中度和重度放牧区的结论一致。而且重度放牧显著降低了生殖枝所占的比例,相反显著增大了营养枝的比例(图7b)。轻度放牧使短花针茅的种子产量显著增加,而重度放牧使之显著降低(图5b),且轻度放牧种子产量占地上生物量的比例最高(图6)。随着载畜率的增大,家畜的采食使短花针茅光合产物积累减少,地上生物量降低(图5a),植物的生殖生长因无法得到足够的资源而受到抑制(王仁忠,2000)。在严酷的生境条件下,植物趋向于将更多的资源分配给营养繁殖而不是有性生殖,所以其会减少种子的资源投入(Pluess et al.,2005)。随着放牧强度的增加,短花针茅的分蘖减少,生殖枝的分化率降低,种子产量减少。同时种子形成的幼苗存活率降低,实生苗减少。分蘖的减少和种子繁殖减弱,使其在群落中逐渐减少,甚至消失(杜利霞等,2008;杜利霞等,2007)。
4结论
(1)轻度放牧促进了短花针茅的生长和分蘖,其地上生物量、盖度、分蘖数和株丛径在轻度放牧区达到最大。
(2)轻度放牧同时促进了短花针茅的有性繁殖,其生殖枝数、种子数及资源在生殖分配上的比例增大,而重度放牧使有性生殖能力显著降低。
(3)轻度放牧(载畜率为0.91 sheep∙hm-2∙0.5 a-1)是短花针茅种群的最适载畜率,也是其生长和繁殖的载畜率阈值。
BUTLER J L, BRISKE D D. 1988. Population structure and tiller demography of the bunchgrass Schizachyrium scoparium in response to herbivory [J]. Oikos, 51(3): 306-312.
DONG S K, WANG X X, LIU S L, et al. 2015. Reproductive responses of alpine plants to grassland degradation and artificial restoration in the Qinghai-Tibetan Plateau [J]. Grass and Forage Science, 70(2): 229-238.
IKEGAMI M, WHIGHAM D F, WERGER M J A, et al. 2007. Responses of rhizome length and ramet production to resource availability in the clonal sedge Scirpus olneyi A. Gray [J]. Plant Ecology, 189(2): 247-259.
LI C L, HAO X Y, ZHAO M L, et al. 2008. Influence of historic sheep grazing on vegetation and soil properties of a Desert Steppe in Inner Mongolia [J]. Agriculture, Ecosystems & Environment, 128(1-2): 109-116.
LIN Y, HAN G D, ZHAO M L, CHANG S X, et al. 2010a. Spatial vegetation patterns as early signs of desertification: a case study of a desert steppe in Inner Mongolia, China [J]. Landscape Ecology, 25(10): 1519-1527.
LIN Y, HONG M, HAN G D, et al. 2010b. Grazing intensity affected spatial patterns of vegetation and soil fertility in a desert steppe [J]. Agriculture, Ecosystems & Environment, 138(3-4): 282-292.
MILLER T E X, TYRE A J, LOUDA S M, et al. 2006. Plant reproductive allocation predicts herbivore dynamics across spatial and temporal scales [J]. American Naturalist, 168(5): 608-616.
MOREIRA X, ABDALA-ROBERTS L, LINHART Y B, et al. 2015. Effects of climate on reproductive investment in a masting species: assessment of climatic predictors and underlying mechanisms [J]. Journal of Ecology, 103(5): 1317-1324.
NEVES A C, NOGUEIRA F B, de ASSIS L R, et al. 2014. Reproductive allocation in rhizomatous, seminiferous, and pseudoviviparous Leiothrix (Eriocaulaceae) species [J]. Plant Ecology, 215(9): 987-996. PLUESS A R, STOCKLIN J. 2005. The importance of population originand environment on clonal and sexual reproduction in the alpine plant Geum reptans [J]. Functional Ecology, 19(2): 228-237.
WANG X, TAUB D R, JABLONSKI L M, et al. 2015. Reproductive allocation in plants as affected by elevated carbon dioxide and other environmental changes: a synthesis using meta-analysis and graphical vector analysis [J]. Oecologia, 177(4): 1075-1087.
WEINER J, CAMPBELL L G, PINO J, et al. 2009. The allometry of reproduction within plant populations [J]. Journal of Ecology, 97(6): 1220-1233.
WU G L, HU T M, LIU Z H, et al. 2010. Trade-off of sexual and asexual recruitment in a dominant weed ligularia virgaurea (maxim.) In alpine grasslands (China) [J]. Polish Journal of Ecology, 58(1): 81-86.
安慧, 徐坤. 2013. 放牧干扰对荒漠草原土壤性状的影响[J]. 草业学报, 22(4): 35-42.
白永飞, 李德新, 许志信, 等. 1999. 牧压梯度对克氏针茅生长和繁殖的影响[J]. 生态学报, 19(4): 479-484.
杜利霞, 李青丰, 董宽虎. 2007. 放牧强度对短花针茅草原冷蒿繁殖特性的影响[J]. 草地学报, 15(4): 367-370.
杜利霞, 李青丰. 2008. 放牧对荒漠草原短花针茅繁殖特性的影响[J].山西农业大学学报(自然科学版), 28(1): 4-6.
杜宇凡, 古琛, 乌力吉, 等. 2016. 不同载畜率下短花针茅荒漠草原物种多度格局[J]. 生态学杂志, 35(3): 675-683.
段敏杰, 高清竹, 万运帆, 等. 2010. 放牧对藏北紫花针茅高寒草原植物群落特征的影响[J]. 生态学报, 30(14): 3892-3900.
古琛, 杜宇凡, 乌力吉, 等. 2015. 载畜率对荒漠草原群落及植物功能群生物量的影响[J]. 生态环境学报, 24(12): 1962-1968.
韩国栋, 焦树英, 毕力格图, 等. 2007. 短花针茅草原不同载畜率对植物多样性和草地生产力的影响[J]. 生态学报, 27(1): 182-188.
侯牡丹, 王明玖, 李志强, 等. 2009. 放牧强度对短花针茅草原3种主要植物营养繁殖性能的影响[J]. 内蒙古农业大学学报(自然科学版), 30(1): 71-75.
黄琛, 张宇, 王静, 等. 2014. 不同放牧强度下短花针茅荒漠草原植被的空间异质性[J]. 植物生态学报, 38(11): 1184-1193.
李西良, 刘志英, 侯向阳, 等. 2015. 放牧对草原植物功能性状及其权衡关系的调控[J]. 植物学报, 50(2): 159-170.
李永宏. 1988. 内蒙古锡林河流域羊草草原和大针茅草原在放牧影响下的分异和趋同[J]. 植物生态学与地植物学学报, 12(3): 27-34.
刘文亭, 卫智军, 吕世杰, 等. 2016. 内蒙古荒漠草原短花针茅叶片功能性状对不同草地经营方式的响应[J]. 生态环境学报, 25(3): 385-392.
刘志民, 蒋德明, 高红瑛, 等. 2003. 植物生活史繁殖对策与干扰关系的研究[J]. 应用生态学报, 14(3): 418-422.
蒙荣, 胡秋芳, 卫智军, 等. 2004. 不同放牧制度大针茅草原植物种群有性繁殖能力研究[J]. 中国草地, 26(4): 11-15.
潘占磊, 王忠武, 韩国栋, 等. 2016. 短花针茅荒漠草原甲烷通量对增温和施氮的响应[J]. 生态环境学报, 25(2): 209-216.
乔丽青, 田大栓, 万宏伟, 等. 2014. 不同载畜率下瓣蕊唐松草的生长和繁殖对策[J]. 植物生态学报, 38(8): 878-887.
任海彦, 郑淑霞, 白永飞. 2009. 放牧对内蒙古锡林河流域草地群落植物茎叶生物量资源分配的影响[J]. 植物生态学报, 33(6): 1065-1074.
戎郁萍, 韩建国, 王培, 等. 2001. 放牧强度对草地土壤理化性质的影响[J]. 中国草地, 23(4): 42-48.
宋磊, 董全民, 李世雄, 等. 2016. 放牧对青海湖北岸高寒草原植物群落特征的影响[J]. 草业科学, 33(8): 1625-1632.
孙世贤, 卫智军, 陈立波, 等. 2013. 放牧强度季节调控对短花针茅荒漠草原土壤养分的影响[J]. 生态环境学报, 22(5): 748-754.
王晨晨, 王珍, 张新杰, 等. 2014. 增温对荒漠草原植物群落组成及物种多样性的影响[J]. 生态环境学报, 23(1): 43-49.
王仁忠. 2000. 放牧影响下羊草种群生殖生态学的研究[J]. 应用生态学报, 11(3): 399-402.
卫智军, 张爽, 吕世杰, 等. 2016. 春季休牧后放牧强度变化对短花针茅荒漠草原主要植物种群空间分布的影响[J]. 生态环境学报, 25(5): 729-736.
张勃, 陈海军, 侯向阳, 等. 2015. 内蒙古锡林郭勒草原贝加尔针茅的繁殖特性及其生态响应[J]. 甘肃农业大学学报, 50(4): 103-108.
张晓娜, 哈达朝鲁, 潘庆民. 2010. 刈割干扰下内蒙古草原两种丛生禾草繁殖策略的适应性调节[J]. 植物生态学报, 34(3): 253-262.
张新杰, 韩国栋, 王忠武. 2014. 不同载畜率对短花针茅荒漠草原土壤呼吸的影响[J]. 生态环境学报, 23(5): 743-748.
The Response of Growth and Reproduction for Stipa breviflora to Different Stocking Rates
GU Chen, ZHAO Tianqi, WANG Yating, WANG Xi, JIA Lixin, ZHAO Mengli*
Collage of Grassland, Resources and Environment, Inner Mongolia Agricultural University Hohhot, Inner Mongolia 010019, China
Resource allocation between growth and reproduction, the two basic life processes, are the core of plant life-history strategy. Plants have to balance the sexual and asexual reproduction to adapt to grazing disturbance. In our study, a completely randomized block design experiment with four different stocking rates was set up in the field to investigate the effects of different stocking rates on the growth and reproduction of Stipa breviflora, the dominant specie on desert steppe in Inner Mongolia. It is of ecological significance to reveal the response of S. breviflora from grazing disturbance. This study provides a reference for the protection of desert steppe ecosystem, as well as, grassland grazing management. Results showed that: (1) The foliage coverage of S. breviflora was higher under light grazing and lower under heavy grazing (P<0.000 1), while the aboveground biomass both for communities and individual and height decreased (P<0.000 1) but the density conversely increased (P<0.000 1) with the increase in stocking rates. (2) Both number of tillers (P=0.048) and diameter (P=0.023) of S. breviflora tussock increased under light grazing, and tillers depth decreased (P=0.042) with increasing stocking rates. (3) The seed yield and proportion of S. breviflora was highest with light grazing but decreased in heavy grazing (P<0.000 1). And (4) likewise, both the total and reproductive tillers of S. breviflora were highest (P<0.000 1) under light grazing, and then decreased with increasing stocking rates, under heavy grazing, the proportion of reproductive tillers was decreased, while vegetative tillers’ proportion increased (P<0.000 1). Growth promotion of S. breviflora, as well as, tillers and reproduction were heightened by light grazing, but decreased with increase stocking rates. Results suggested that light grazing (stocking rate is 0.91 sheep∙hm-2∙0.5 a-1) is the optimal stocking rate and the threshold for both of the growth and reproduction of S. breviflora in desert steppe of Inner Mongolia.
desert steppe; grazing intensity; community trait; tiller; sexual reproduction
10.16258/j.cnki.1674-5906.2017.01.006
S812.29; X17
A
1674-5906(2017)01-0036-07
古琛, 赵天启, 王亚婷, 王玺, 贾丽欣, 赵萌莉. 2017. 短花针茅生长和繁殖策略对载畜率的响应[J]. 生态环境学报, 26(1): 36-42.
GU Chen, ZHAO Tianqi, WANG Yating, WANG Xi, JIA Lixin, ZHAO Mengli. 2017. The Response of growth and reproduction for Stipa breviflora to different stocking rates [J]. Ecology and Environmental Sciences, 26(1): 36-42.
国家自然科学基金项目(31660108;31460110);内蒙古农业大学草地资源教育部重点实验室资助项目
古琛(1988年生),男,博士研究生,主要从事草地生态学方向研究。E-mail: guchen0706@sina.com *通信作者。E-mail: nmgmlzh@126.com
2016-10-17
