围栏措施对青藏高原高寒草甸群落结构和稳定性的影响
2017-03-27罗久富周金星赵文霞董林水郑景明
罗久富,周金星,赵文霞,董林水,郑景明
(1.北京林业大学林学院,北京 100083; 2.北京林业大学 水土保持国家林业局重点实验室,北京 100083;3.滨州学院 山东省黄河三角洲生态环境重点实验室,山东 滨州 256603)
围栏措施对青藏高原高寒草甸群落结构和稳定性的影响
罗久富1,2,周金星2,赵文霞1,董林水3,郑景明1
(1.北京林业大学林学院,北京 100083; 2.北京林业大学 水土保持国家林业局重点实验室,北京 100083;3.滨州学院 山东省黄河三角洲生态环境重点实验室,山东 滨州 256603)
人工条件下高寒脆弱生态系统群落结构及其稳定性的变化是研究其生态功能的重要内容。本研究以乌玛塘高寒草甸示范区为例,对研究区植被进行植物群落学调查,对植物群落功能群和稳定性进行分析,研究了高寒草甸植物群落在围栏4年和8年后,围栏内外群落的结构变化和稳定性差异。结果表明,1)围栏内物种多样性大于围栏外;围栏4年后群落内一年生、二年生植物重要值明显高于围栏外;围栏8年后,群落优势种发生变化,高山嵩草(Kobresiapygmaea) 优势度降低,一年生、二年生植物甘肃马先蒿(Pediculariskansuensis)成为优势物种之一;多年生禾草,杂类草以及一年生、二年生植物3个功能群所含物种数显著多于围栏外(P<0.05);2)围栏4年后群落稳定性表现为围栏内>围栏外,8年后表现为围栏内<围栏外。短期围栏使得群落稳定性高于未围栏的群落,但是群落结构差异并不明显,随着围栏时间增加,群落结构发生明显变化,且群落稳定性降低,围栏外群落则趋向于稳定。
高寒草甸;功能群;M. Godron;退化;毒杂草;高山嵩草;甘肃马先蒿
草地生态系统通常为自然、半自然生态系统类型,是受人类活动影响最大的陆生生态系统之一,在生态系统功能及其维持机理等方面得到了广泛的研究[1-4]。在草地植物群落研究中,围栏措施作为一种可控的方法被广泛地应用。随着研究的深入,植物功能群(plant functional group)作为植被随环境变化的基本单元逐渐被引入到研究中,是对外界干扰和环境因子具有相似响应的一类群体[5-6]。通过植物群落功能群的分析,可以探讨植物群落发展过程中群落结构的变化。围栏封育后青藏高原草地禾本科和莎草科的生物量在群落内迅速增加,而杂类草的生物量逐渐降低[7-8],围封后内蒙古草原群落内禾草以及一年生植物丰富度降低,灌木增加,群落朝退化方向发展[9]。围栏对群落结构时间效应的影响也备受关注,研究发现,随着围封年限的增加,高寒草原群落盖度和生产力呈先显著增加后显著降低的趋势[10],而短期围栏对群落的物种组成影响并不显著[11]。与此相反,也有研究发现短期的围栏措施后,高山嵩草(Kobresiapygmaea)和矮生嵩草(K.humilis)草地的功能群多样性相对于放牧区发生了显著改变[12]。以上研究表明,研究区的生物与非生物因子等的差异,致使各研究的结果并未得到一致的结论。
同时,人类活动范围的扩大和强度的增加使草地植物群落的稳定性备受关注[13]。稳定性的概念伴随系统控制论而产生,1955年MacArthur提出稳定性的概念,之后围绕着稳定性的概念、影响因素、维持机制以及评价方法等展开了诸多研究,但是结果不尽一致,综合各研究可将稳定性归纳为演替稳定性、抵抗力稳定性和恢复力稳定性3个方面[14-15]。同时,多稳态(alternative stable states)理论指出,反馈循环的差异会导致生态系统稳态发生转换,伴随产生的是群落结构和功能的变化[16],群落结构和功能的变化则反映了群落内物质流、能量流以及信息流的变化,这些变化与生态系统的服务价值密切相关,所以,群落稳定性的研究对于评估生态系统功能、价值以及指导生产有着非常重要的作用。研究发现,不同生境中的植物群落稳定性即使在相同的处理措施下,也会存在极大的差异,围栏对青海湖流域高寒草甸、高寒草原以及温性草原群落的稳定性影响并不相同,在干旱地区水分因素是影响群落稳定性的重要因子[17-18],科罗拉多州北部禾草草原群落内优势种种群对群落稳定性的影响力超过了物种多样性[19]。综合前人研究结果,研究区域的不同加上群落本身的复杂性致使诸多研究难以得出一致的结论,在海拔4 000 m以上的高原生态系统中,相关的研究成果更少,因此,本研究将研究区定位于海拔4 500 m以上的青藏高原高寒草甸区,分析不同围栏年限下的高寒草甸植物群落结构和稳定性差异。
高寒草甸生态系统是在寒冷而湿润的气候条件下,由耐寒的多年生中生草本植物为建群种而形成的一种生态系统类型,广泛分布于青藏高原,是高原畜牧业生产的物质基础,也是我国内陆及周边地区巨大的生态屏障[20]。但是近年来气候的变化、人类活动范围的扩张和资源利用强度的加剧,致使高海拔脆弱生态区受到严重干扰[21]。因此,研究高海拔脆弱生态区植物群落的稳定性及其功能群结构有助于加深了解植物群落本身稳定性的特征和规律,为开展植物群落保护利用、促进和维持植被的健康发展提供帮助。本研究对西藏乌玛塘高寒草甸示范区围封的植物群落与开放植物群落结构和稳定性进行研究,利用改进后的数学生态学方法——M. Godron稳定性测度法和功能群分析方法,定性定量评价围栏对青藏高原高寒草甸植物群落结构和稳定性的影响,旨为丰富青藏高原高寒草甸生态系统植被研究体系。
1 材料与方法
1.1 研究区域概况
高寒草甸样地乌玛塘示范区(91°32′ E,30°32′ N),海拔4 546 m,地处当雄和那曲交接处,距当雄县城46 km。示范区属于高原亚高寒带季风半干旱气候区,年平均气温-7~0 ℃,年降水量300~400 mm。极端低温-35.9 ℃,极热月(7月)平均气温10.8 ℃,≥10 ℃的年积温280.7 ℃·d,无霜期20 d,年蒸发量1 831.6 mm,年相对湿度50%,全年日照时数2 881.0 h。示范区土壤主要由高山草甸、草原土组成,表层具有约10 cm厚的草皮层,30~50 cm厚的腐殖层,氮、磷、钾含量高。示范区植被主要是以高山嵩草为优势种的高寒草甸植物群落,常见伴生种有钉柱委陵菜(Potentillasaundersiana)、矮生嵩草、早熟禾(Poasp.)、黄耆(Astragalussp.)等。
1.2 研究方法
1.2.1 样地设置及调查 实验示范区于2005年6月建设,设置20 m×50 m围栏,围栏外全年放牧,主要放牧家畜为牦牛和绵羊,放牧强度约2绵羊单位·hm-2,至2013年围栏内外群落差异明显,分别于2009年8月和2013年8月于植物生长盛期进行调查。参考《陆地生态学研究方法》[22]中草本植物调查样地大小设置方法,在围栏内外各布设9个样地,样地大小为1 m×1 m。调查记录植物种类组成、物种盖度、高度等。
1.2.2 划分功能群 本研究根据生活型及分蘖类型将功能群划分为:豆科(legumes),多年生禾草(perennial grasses),莎草(sedges),多年生杂类草(forbs),一年生、二年生植物(annuals and biennials)。
1.2.3 植物群落稳定性测度方法 M. Godron稳定性测定方法[23]是目前生态学研究应用较为广泛的一种测定群落稳定性的方法。其原理是将所研究群落中所有植物的频度,按从大到小排列,计算总种数倒数的累计百分数和相对频度的累计百分数,对二者做散点图,用一条曲线连接各散点,同时做直线y=100-x,与曲线交点即稳定性参考点(x,y)。交点坐标越趋近于(20,80),说明群落越稳定,反之,越不稳定。关于草地群落稳定性的研究表明,利用盖度计算群落的空间稳定性结果更具说服力[11,17],所以在本研究中将M. Godron方法中的相对频度换成相对盖度,同时为了摒除人为主观判断带来的误差,研究中采用数学模型对数据进行模拟[24]。本研究中,为构建更好的拟合曲线,采用一元三次方程曲线模拟散点图,点坐标为(a,b),建立如下数学方程:
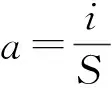
b=∑(C1+C2+C3+…Ci) (i=1,2,3…S)
平滑曲线模拟模型:
y=ax3+bx2+cx+d
(1)
直线方程:
y=100-x
(2)
式(1)中:x和y分别代表总种数倒数的累计百分数以及相对盖度的累计百分数。将式(2)带入式(1)并求解,根据实际情况,舍弃远大于100的结果,由此得到直线与曲线的交点坐标(x,y),计算(x,y)与(20,80)的欧氏距离。S为群落内物种数,Ci为第i个物种的相对盖度,物种排列顺序按照其盖度值从大到小排列。
1.2.4 物种多样性 以群落内物种的数目表示多样性R=S。S为群落内物种数。
1.2.5 物种重要值 采用重要值综合指标[25-26]来处理数据:



2 结果与分析
2.1 围栏对高寒草甸植物群落结构的影响
2.1.1 对功能群优势度的影响 围栏4年后,围栏内外群落均为典型的高山嵩草群落,围栏内莎草功能群重要值为0.520,围栏外为0.568,围栏内高山嵩草重要值低于围栏外(表1)。其次,多年生杂类草和多年生禾草在围栏内外群落也占有较高的重要值。一年生、二年生植物和豆科植物所占比例较少,其中一年生、二年生植物诸如鳞叶龙胆、两裂婆婆纳等细小柔弱性草本种类明显多于围栏外。

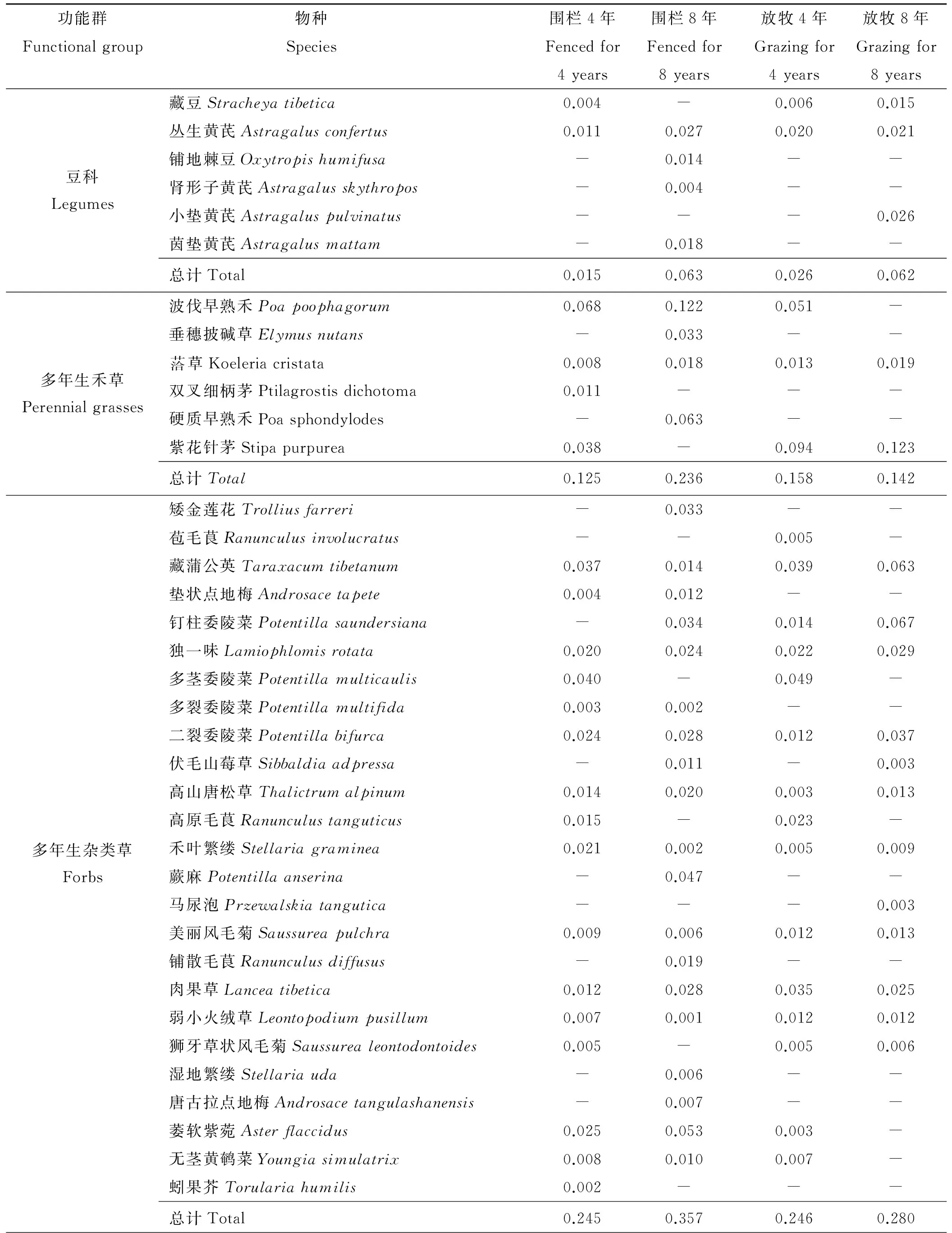
表1 围栏内外高寒草甸植物生活型功能群组成及其重要值
续表1
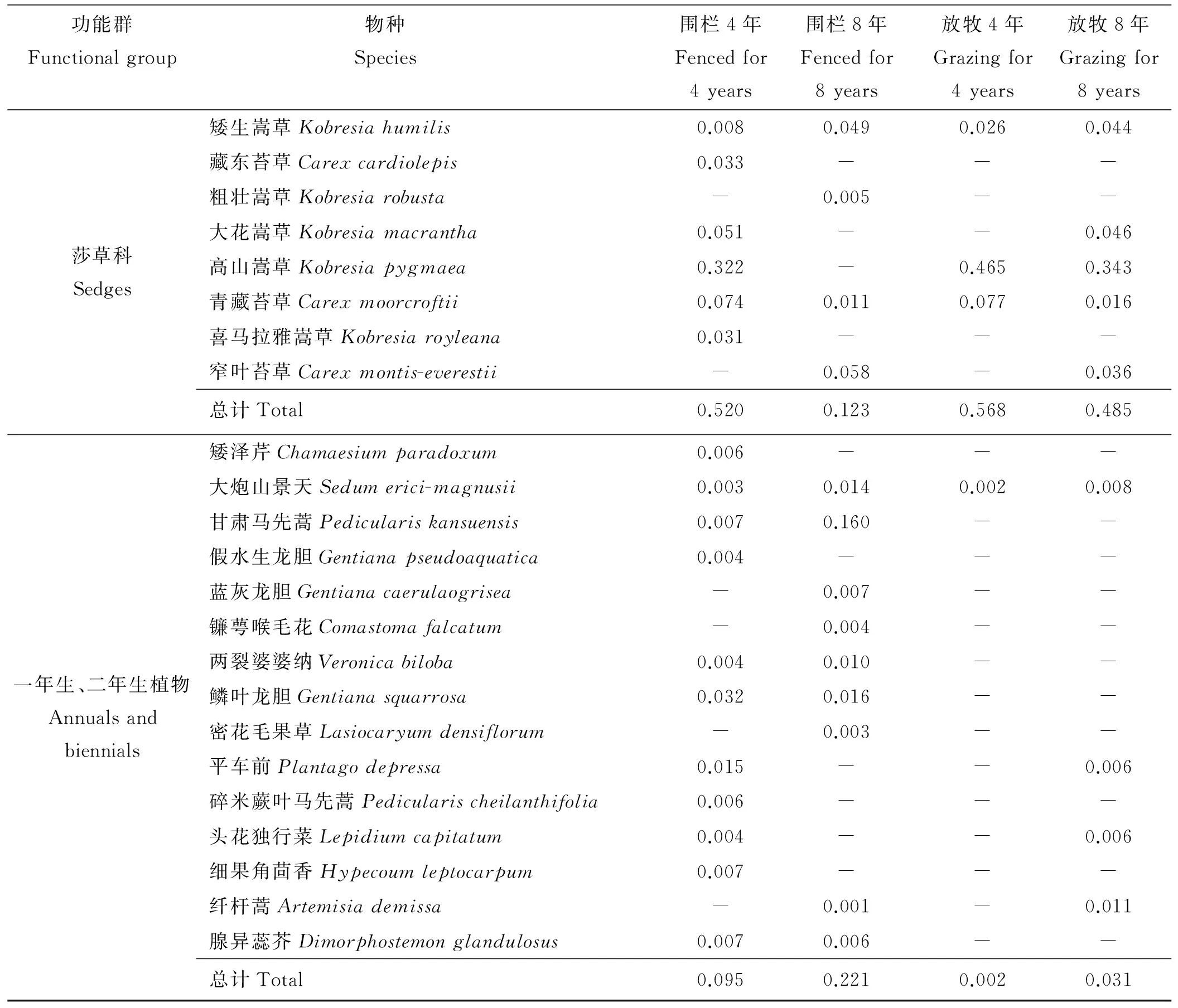
功能群Functionalgroup物种 Species 围栏4年Fencedfor4years围栏8年Fencedfor8years放牧4年Grazingfor4years放牧8年Grazingfor8years莎草科Sedges矮生嵩草Kobresiahumilis0.0080.0490.0260.044藏东苔草Carexcardiolepis0.033---粗壮嵩草Kobresiarobusta-0.005--大花嵩草Kobresiamacrantha0.051--0.046高山嵩草Kobresiapygmaea0.322-0.4650.343青藏苔草Carexmoorcroftii0.0740.0110.0770.016喜马拉雅嵩草Kobresiaroyleana0.031---窄叶苔草Carexmontis-everestii-0.058-0.036总计Total0.5200.1230.5680.485一年生、二年生植物Annualsandbiennials矮泽芹Chamaesiumparadoxum0.006---大炮山景天Sedumerici-magnusii0.0030.0140.0020.008甘肃马先蒿Pediculariskansuensis0.0070.160--假水生龙胆Gentianapseudoaquatica0.004---蓝灰龙胆Gentianacaerulaogrisea-0.007--镰萼喉毛花Comastomafalcatum-0.004--两裂婆婆纳Veronicabiloba0.0040.010--鳞叶龙胆Gentianasquarrosa0.0320.016--密花毛果草Lasiocaryumdensiflorum-0.003--平车前Plantagodepressa0.015--0.006碎米蕨叶马先蒿Pedicularischeilanthifolia0.006---头花独行菜Lepidiumcapitatum0.004--0.006细果角茴香Hypecoumleptocarpum0.007---纤杆蒿Artemisiademissa-0.001-0.011腺异蕊芥Dimorphostemonglandulosus0.0070.006--总计Total0.0950.2210.0020.031
注:“-”表示该物种未出现。
Note: “-”means the plants didn’t appear.
从以上结果可以看出,围栏后各功能群物种组成以及物种数与围栏外群落相比均发生了较大的变化。随着围栏时间增加,围栏内高山嵩草群落退化,而多年生禾草功能群的波伐早熟禾和一年生、二年生植物功能群的甘肃马先蒿迅速繁殖,同时其它杂类草侵入,使得围栏内群落杂类草优势度上升,群落性质相对于围栏外群落,已经发生了改变。
2.1.2 对各功能群所含物种数的影响 围栏后,围栏内外群落物种数及各生活型功能群所含物种数呈现出较明显不同。围栏4年及8年后,围栏内群落总物种分别39种和40种,围栏外分别24种和26种。围栏4年后,围栏内一年生、二年生植物物种数为2.11,显著高于围栏外(0.11)(P<0.05),其余各功能群围栏内均差异不显著(P>0.05)。围栏8年后,围栏内外豆科物种数分别为1.56和1.44,莎草科植物分别为1.67和1.78,这两功能群内物种数差异均不显著,其它功能群物种数均为围栏内显著高于围栏外(P<0.05),其中围栏内外禾本科物种数分别为2.56和1,杂类草分别为8.67和6.11,一年生、二年生植物分别为3.89和0.44(图1)。
2.2 围栏后高寒草甸植物群落稳定性变化
根据改进后的M. Godron稳定性测定方法,对群落进行平滑曲线模拟(图2)。求得平滑曲线和直线的交点坐标,结合M. Godron稳定性原理,即交点坐标(x,y)越接近平衡点(20,80),说明该群落越稳定,反之则越不稳定。为直观表达,将交点坐标与平衡点坐标的距离以欧式距离表示(表2)。
寒草甸研究区群落M. Godron稳定性判定结果(表2)表明,围栏4年后,围栏内群落M. Godron稳定性判定欧氏距离为1.97,围栏外为10.99,8年后,围栏内M. Godron稳定性判定欧氏距离为8.71,围栏外为1.78。根据原理,与平衡点(20,80)越接近,欧氏距离越小,群落稳定性越高。因此,在研究示范区,围栏4年后,群落M. Godron稳定性高于围栏外群落,8年后,群落M. Godron稳定性低于围栏外群落。围封4年后的围栏外群落,高山嵩草决定散点图的第1个点,坐标为(4.17,81.95),导致该散点图所有点均在y=80之上,散点图拟合曲线R2=0.983 4,拟合曲线与直线的交点离平衡点较远。围栏内外群落稳定性变化趋势相反,短期的围栏措施使得群落稳定性较好,而随着时间增加,群落的稳定性降低,围栏外群落达到稳定状态滞后于围栏内。
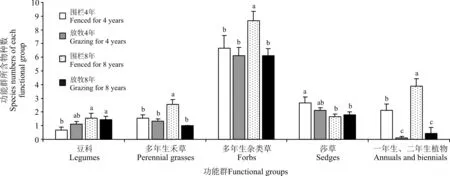
图1 围栏内与围栏外群落各功能群所含物种数
注:不同小写字母表示同一功能群不同处理间差异显著(P<0.05)。
Note: Different lowercase letters within a functional group indicate significant differences at the 0.05 levle.
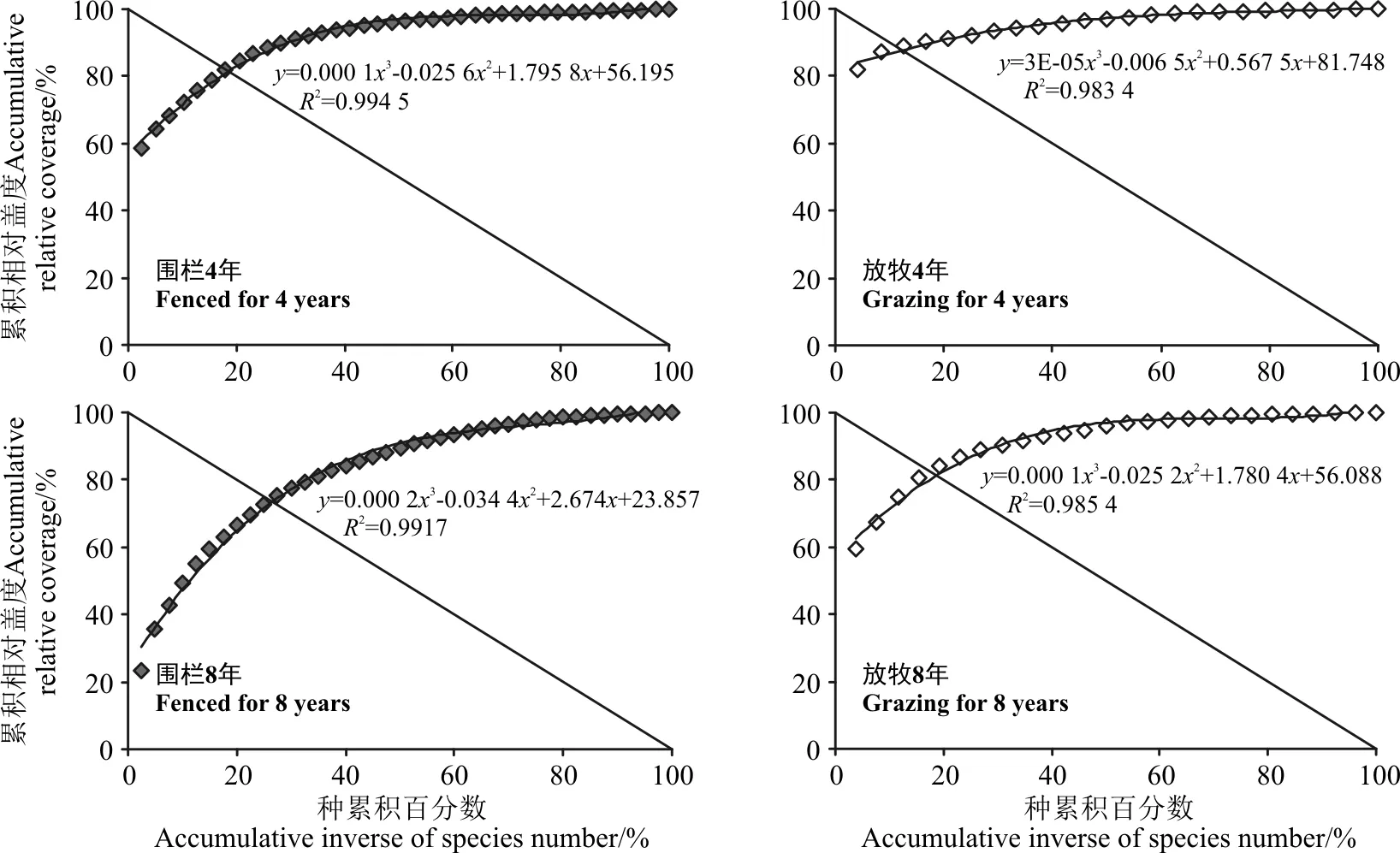
图2 围栏4、8年后围栏内外植被的M. Godron稳定性拟合曲线
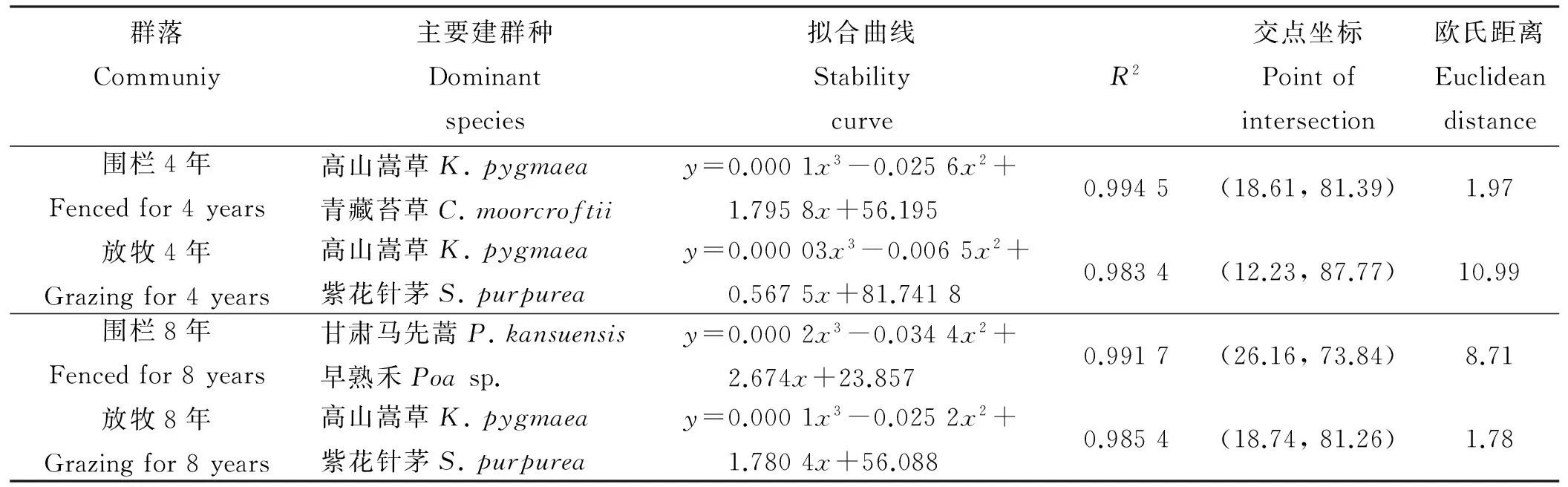
群落Communiy主要建群种Dominantspecies拟合曲线StabilitycurveR2交点坐标Pointofintersection欧氏距离Euclideandistance围栏4年Fencedfor4years放牧4年Grazingfor4years高山嵩草K.pygmaea青藏苔草C.moorcroftiiy=0.0001x3-0.0256x2+1.7958x+56.1950.9945(18.61,81.39)1.97高山嵩草K.pygmaea紫花针茅S.purpureay=0.00003x3-0.0065x2+0.5675x+81.74180.9834(12.23,87.77)10.99围栏8年Fencedfor8years放牧8年Grazingfor8years甘肃马先蒿P.kansuensis早熟禾Poasp.y=0.0002x3-0.0344x2+2.674x+23.8570.9917(26.16,73.84)8.71高山嵩草K.pygmaea紫花针茅S.purpureay=0.0001x3-0.0252x2+1.7804x+56.0880.9854(18.74,81.26)1.78
3 讨论与结论
3.1 高寒草甸群落结构对围栏措施的响应
围栏影响青藏高原高寒草甸区植物群落功能群优势度及其物种组成。对青海湖高寒草原围栏处理研究发现,群落多样性和物种丰富度随着围栏时间增加而显著降低[10],本研究表明,围栏后,群落物种丰富度高于围栏外,部分功能群所含的物种数也较高。造成这种差异的原因可能是本研究区海拔较高,生态系统极其脆弱,围栏后群落种间关系改变,使群落演替趋势发生了变化[27]。群落功能群优势度发生改变,围栏后,杂类草迅速繁殖,以甘肃马先蒿为主的毒杂草侵入,甘肃马先蒿有强大的种子繁殖能力以及集群分布形式,是高寒草甸群落的主要伴生杂草,同时也是重度退化的高寒草甸优势毒性杂草之一[28-29],是高寒草甸退化过程中的一年生、二年生毒杂草阶段的代表性植物,反映了高寒草甸生态系统的退化程度以及退化草甸生态系统向顶级群落恢复的演替进程[30]。甘肃马先蒿侵入后,迅速繁殖,占据大量生态位,严重威胁原生群落的优良牧草[31],使原生群落中高山嵩草的繁殖受到抑制,逐渐降低其在围栏内群落的生态优势度,群落退化[32]。而围栏外群落仍是以高山嵩草为主要优势种的高寒草甸群落。
围栏内群落一年生、二年生植物种类与围栏外群落相比差异明显,相比于甘肃马先蒿、龙胆、镰萼喉毛花等,围栏外群落中的平车前、纤杆蒿、头花独行菜等株型低矮,或具匍匐茎,更耐踩踏,因此,能适应有人类活动以及牲畜牧食的生境。同时,如垂穗披碱草、早熟禾等株高较高的物种也未在围栏外出现,而是以耐踩踏、耐牧性强的嵩草、紫花针茅等为主,这说明在群落中存在生态补偿作用(ecological compensation),即功能群对异质化的生境在生态适应性上表现出多样化,在各功能群之间形成生态补偿作用,同时在功能群内部的物种之间也形成补偿作用[1,33]。但是,围栏封育10年后,鄂尔多斯不同类型的草地群落内猪毛菜(Salsolacollina)等一年生植物和牛心朴子(Cynanchumkomarovii)等毒杂草植物大幅减少,而适口性好、营养价值高的禾草类和豆科植物等优良牧草成为主要建群种[34];未实行围栏封育的美国科罗拉多州矮草草原植物群落中一年生植物比围栏封育的群落大[35]。而本研究中围栏后,一年生、二年生植物多于围栏外,可能是围栏措施致使土壤板结,土壤水分的运输受限,限制多年生植被生长[18],而更适于一年生、二年生等以种子繁殖为主的植物。
3.2 高寒草甸群落稳定性对围栏措施的响应
围栏影响高寒草甸植物群落稳定性,生态系统的稳定性除与生物多样性存在关系外,与干扰类型和程度等都存在密切关系[36]。杨力军等[37]研究表明,在高寒草甸区,只有以嵩草属为主的植被才能具有较高的稳定性。但本研究表明,围栏4年后,围栏内外优势种均为高山嵩草,围栏外群落的高山嵩草重要值为0.465,但是稳定性却较低。高寒草甸生态系统的稳定性受到功能群内部物种的均匀度、密度以及物种本身性质的影响[38-39],所以围栏外群落在围栏4年后年稳定性较低,与高山嵩草较高的优势度等群落本身的结构密切相关。
研究发现,科罗拉多州北部矮草草原群落内优势种对群落稳定性的影响力强于物种多样性[19]。本研究表明,围栏8年后(2013年),群落稳定性低于围栏外群落。分析可知,围栏外群落长期处于以高山嵩草为最主要优势种的高寒草甸群落,而围栏内的群落性质则随时间推移发生了变化,由嵩草群落演变为毒杂草群落,优势种发生改变,甘肃马先蒿重要值迅速增加,而甘肃马先蒿等毒杂草强大的种子繁殖能力和化感作用会对周围优良牧草产生抑制作用,使得群落逆向演替加剧[40],群落稳定性较差;同时,围栏内高山嵩草迅速退化,杂类草优势度增加,杂类草之间的竞争导致该群落稳定性较低。因此,从草地健康发展考虑,长期的围栏措施是不可取的[41]。
群落稳定性与功能群结构是评价群落健康状态的指标。本研究表明,短期围栏后,群落稳定性高于围栏外群落,但是群落结构差异并不明显,群落性质未发生明显改变,随着围栏时间增加,群落结构发生明显变化,且群落稳定性降低,围栏外群落则趋向于稳定,另外,群落优势种种群变化影响群落稳定性变化。但本研究中,围封样地处于生态环境脆弱,对外界刺激响应敏感的高寒地区,该结果是否能推广到其它群落还有待进一步验证。同时,本研究结果对高寒脆弱生态区植被恢复具有一定程度的参考意义,建议因地制宜,合理划区轮牧强度和控制放牧时长[41],将短期围栏作为草地管理的措施之一。
References:
[1] 白永飞,陈佐忠.锡林河流域羊草草原植物种群和功能群的长期变异性及其对群落稳定性的影响.植物生态学报,2000,24(6):641-647. Bai Y F,Chen Z Z.Effects of long-term variability of plant species and functional groups on stability of aLeymuschinensiscommunity in the Xilin River Basin,Inner Mogolia.Acta Plytoecologica Sinica,2000,24(6):641-647.(in Chinese)
[2] Schlapfer F,Schmid B.Ecosystem effects of biodiversity:A classification of hypotheses and exploration of empirical results.Ecological Applications,1999,9:893-912.
[3] 曹广民,林丽,张法伟,李以康,韩道瑞,龙瑞军.青藏高原高寒矮嵩草草甸稳定性的维持、丧失与恢复.草业科学,2010,27(8):34-38. Cao G M,Lin L,Zhang F W,Li Y K,Han D R,Long R J.A review of maintenance,loss and recovery of stability of alpineKobresiahumilismeadow on Tibetan Plateau.Pratacultural Science,2010,27(8):34-38.(in Chinese)
[4] Luzuriaga A L,Escudero A.What determines emergence and net recruitment in an early succession plant community?Disentangling biotic and abiotic effects.Journal of Vegetation Science,2008,19:445-456.
[5] Smith T M,Woodward F I,Shugart H H.Plant Functional Types.Cambridge:Cambridge University Press,1996.
[6] Woodward F I,Cramer W.Plant functional types and climatic changes:Introduction.Journal of Vegetation Science,1996,7:306-308.
[7] 扎西文毛,赵建中,严振英,周华坤.围栏封育对退化草地群落功能群的影响.安徽农业科学,2011,39(14):8650-8652. Zhaxi wenmao,Zhao J Z,Yan Z Y,Zhou H K.Study on the functional groups of degraded in fencing.Journal of Anhui Agricultural Science,2011,39(14):8650-8652.(in Chinese)
[8] 毛绍娟,吴启华,祝景彬,李红琴,张法伟,李英年.藏北高寒草原群落维持性能对封育年限的响应.草业学报,2015,24(1):21-30. Mao S J,Wu Q H,Zhu J B,Li H Q,Zhang F W,Li Y N.Response of the maintain performance in alpine grassland to enclosure on Northern Tibetan Plateau.Acta Pratacurea Sinica,2015,24(1):21-30.(in Chinese)
[9] 徐粒,高琼,王亚林.围封6 年对温带典型草原坡地物种多样性及其与地上生物量的关系的影响.生态环境学报,2014,23(3):398-405. Xu L,Gao Q,Wang Y L.Species richness within a six-year slope exclosure in a temperate grassland and its relationship with aboveground biomass.Ecology and Environmental Sciences,2014,23(3):398-405.(in Chinese)
[10] 郑伟,董全民,李世雄,施建军,刘玉,侯宪宽,宋磊.禁牧后环青海湖高寒草原植物群落特征动态.草业科学,2014,31(6):1126-1130. Zheng W,Dong Q M,Li S X,Shi J J,Liu Y,Hou X K,Song L.Dynamics of plant community characteristics of alpine steppe under enclosure around Qinghai Lake.Pratacultural Science,2014,31(6):1126-1130.(in Chinese)
[11] 李中林,秦卫华,周守标,张纪兵,汪贞,王运静.短期围栏封育对红松洼自然保护区群落数量特征的影响.草地学报,2015,23(1):21-26. Li Z L,Qin W H,Zhou S B,Zhang J B,Wang Z,Wang Y J.Effects of short-term enclosure on community quantity characteristics in Hongsongwa Natural Reserve.Acta Agrestia Sinica,2015,23(1):21-26.(in Chinese)
[12] 李洋,严振英,郭丁,王海霞,苏淑兰,李旭东,傅华.围封对青海湖流域高寒草甸植被特征和土壤理化性质的影响.草业学报,2015,24(10):33-39. Li Y,Yan Z Y,Guo D,Wang H X,Su S L,Li X D,Fu H.Effects of fencing and grazing on vegetation and soil physical and chemical properties in an alpine meadow in the Qinghai Lake Basin.Acta Prataculturae Sinica,2015,24(10):33-39.(in Chinese)
[13] 王长庭,龙瑞军,丁路明,来德珍,李有福.草地生态系统中物种多样性、群落稳定性和生态系统功能的关系.草业科学,2005,22(6):1-7. Wang C T,Long R J,Ding L M,Lai D Z,Li Y F.Species diversity,community stability and ecosystem function extension of the continuous views.Pratacultural Science,2005,22(6):1-7.(in Chinese)
[14] MacArthur R.Fluctuations of animal populations,and a measure of community stability.Ecology,1955,36:533-536.
[15] Sennhauser E B.The concept of stability in connection with the gallery forests of the Chaco region.Vegetation,1991,94:1-13.
[16] Suding K N,Gross K L,Houseman G R.Alternative states and positive feedbacks in restoration ecology.Trends in Ecology and Evolution,2004,19(1):46-53.
[17] 王鲜鲜,张克斌,王晓,杨晓辉.宁夏盐池四儿滩湿地-干草原植被群落稳定性研究.生态环境学报,2013,22(5):743-747. Wang X X,Zhang K B,Wang X,Yang X H.Vegeta tion stability of the Siertan wetland-dry grassland in Yanchi,Ningxia.Ecology and Environmental Sciences,2013,22(5):743-747.(in Chinese)
[18] 李璠,周国英,杨路存,徐文华,钟泽兵,宋文珠.围栏封育对青海湖流域主要植物群落多样性与稳定性的影响.水土保持研究,2013,20(4):135-140. Li P,Zhou G Y,Yang L C,Xu W H,Zhong Z B,Song W Z.Effect of fence on biodiversity and stability of the main plant communities in the Qinghai Lake Area.Research of Soil and Water Conservation,2013,20(4):135-140.(in Chinese)
[19] Sasaki T,Lauenroth W K.Dominant species,rather than diversity,regulates temporal stability of plant communities.Oecologia,2011,166:761-768.
[20] 刘敏超,李迪强,温琰茂,栾晓峰.三江源地区生态系统生态功能分析及其价值评估.环境科学学报,2005,25(9):1280-1286. Liu M C,Li D Q,Wen Y M,Luan X F.The ecological function analysis and evaluation of ecosystem in Sanjiangyuan region.Acta Scientiae Circumstantiae,2005,25(9):1280-1286.(in Chinese)
[21] Zhou J X,Yang J,Peng G.Constructing a green railway on the Tibet Plateau:Evaluating the effectiveness of mitigation measures.Transportation Research Part D:Transport and Environment,2008,13:369-376.
[22] 陈吉泉,阳树英.陆地生态学研究方法.北京:高等教育出版社,2014:237. Chen J Q,Yang S Y.Terrestrial Ecology Research Methods.Beijing:Higher Education Press,2014:237.(in Chinese)
[23] Godron M,Daget P,Poissonet J,Poissnet P.Some aspects of heterogeneity in grasslands of cantal.Statistical Ecology,1972,3:397-415.
[24] 郑元润.森林群落稳定性研究方法初探.林业科学,2000,36(5):28-32. Zheng Y R.Comparison of methods for study stability of forest community.Scientia Silvae Sinica,2000,36(5):28-32.(in Chinese)
[25] 王慧敏,庞春花,张峰,赵彩莉.汾河流域中下游湿地植被优势种种间关联性分析.生态学杂志,2012,31(10):2507-2512. Wang H M,Pang C H,Zhang F,Zhao C L.Interspecific association of dominant species of wetland vegetation in the middle and lower reaches of Fenhe River,Shanxi Province of China.Chinese Journal of Ecology,2012,31(10):2507-2512.(in Chinese)
[26] 张金屯.数量生态学.2版.北京:科学出版社,2011:20-109. Zhang J T.Quantitative Ecology.2nd Edition.Beijing:Science Press,2011:20-109.(in Chinese)
[27] 魏斌,王莹,关士琪,高红娟,孔杨云,于应文.青藏高原东北缘高寒草甸珠芽蓼斑块植被构成和种间关联.草业科学,2016,33(8):1618-1624. Wei B,Wang Y,Guan S Q,Gao H J,Kong Y Y,Yu Y W.The vegetation composition and interspecific association of patches distributing inPolygonumviviparrumalpine meadow on northeast Qinghai-Tibet Plateau.Practacultural Science,2016,33(8):1618-1624.(in Chinese)
[28] 赵建平,吴学明,苏旭,祁生贵.青海省甘肃马先蒿资源调查及开发利用前景研究.青海草业,2005,14(2):44-47. Zhao J P,Wu X M,Su X,Qi S G.Study on the resource surveying researching and exploring ofPediculariskansuensisMaxm in Qinghai Province.Qingahi Prataculture,2005,14(2):44-47.(in Chinese)
[29] 周华坤,周立,赵新全,刘伟,严作良,师燕.江河源区“黑土滩”型退化草场的形成过程与综合治理.生态学杂志,2003,22(5):51-55. Zhou H K,Zhou L,Zhao X Q,Liu W,Yan Z L,Shi Y.Degradation process and integrated treatment of “black soil beach”grassland in the source regionsof Yangtze and Yellow Rivers.Chinese Journal of Ecology,2003,22(5):51-55.(in Chinese)
[30] 陈哲,周华坤,赵新全,温军,叶鑫,杨元武.高寒草甸退化对甘肃马先蒿生长状况与花期资源分配的影响.生态环境学报,2010,19(12):2800-2807. Chen Z,Zhou H K,Zhao X Q,Wen J,Ye X,Yang Y W.The effect of alpine meadow degradation onPediculariskansuensis’sgrowth and resource allocation in blooming date.Ecology and Environmental Sciences,2010,19(12):2800-2807.(in Chinese)
[31] 邱正强,马玉寿,施建军,潘多峰,李瑞江.甘肃马先蒿对“黑土型”退化草地垂穗披碱草人工草地的影响.草原与草坪,2006(5):26-29. Qiu Z Q,Ma Y S,Shi J J,Pan D F,Li R J.InfluenceofPediculariskansuensisonElymusnutansartificial grasslandin “BlackSoil Type”:Degenerated alpine grassland.Grassland and Turf,2006(5):26-29.(in Chinese)
[32] 周丽,张德罡,贠旭江,董永平,王加亭,赵雅丽.退化高寒草甸植被与土壤特征.草业科学,2016,33(11):2196-2201. Zhou L,Zhang D G,Yun X J,Dong Y P,Wang J P,Zhao Y L.The vegetation and soil characteristics of degraded alpine meadow.Practacultural Science,2016,33(21):2196-2201.(in Chinese)
[33] 王正文,邢福,祝廷成,李宪长.松嫩平原羊草草地植物功能群组成及多样性特征对水淹干扰的响应.植物生态学报,2002,26(6):708-716. Wang Z W,Xing F,Zhu T C,Li X C.The responses of functional group composition and species diversity ofAneurolepidiumchinensegrassland to flooding disturbance on Songnen Plain,northeastern China.Acta Phytoecologica Sinica,2002,26(6):708-716.(in Chinese)
[34] 张鹏莉,陈俊,崔树娟,程积民.禁牧条件下不同类型草地群落结构特征.生态学报,2013,33(2):0425-0434. Zhang P L,Chen J,Cui S J,Cheng J M.The community characteristics of different types of grassland under grazing prohibition condition.Acta Ecologica Sinica,2013,33(2):0425-0434.(in Chinese)
[35] Alder P B,Lauenroth W K.Livestock exclusion increases the spatial heterogeneity of vegetation in Colorado shortgrass steppe.Applied Vegetation Science,2000,3(2):213-222.
[36] 周华坤,周立,赵新全,刘伟,李英年,古松,周兴民.青藏高原高寒草甸生态系统稳定性研究.科学通报,2006,51(1):63-69. Zhou H K,Zhou L,Zhao X Q,Li Y N,Gu S,Zhou X M.Research on alpine meadow ecosystem stability.Chinese Science Bulletin,2006,51(1):63-69.(in Chinese)
[37] 杨力军,李希来,石德军,孙海群,杨元武.青藏高原“黑土滩”退化草地植被演替规律的研究.青海草业,2005,14(1):2-5. Yang L J,Li X L,Shi D J,Sun H Q,Yang Y W.Research or regulation of vegetation succession in degraded grassland in Qinghai and Tibetan Plateau.Qinghai Prataculture,2005,14(1):2-5.(in Chinese)
[38] 王长庭,龙瑞军,丁路明.高寒草甸不同草地类型功能群多样性及组成对植物群落生产力的影响.生物多样性,2004,12(4):403-409. Wang C T,Long R J,Ding L M.The effects of differences in functional group diversity and composition on plant community productivity in four types of alpine meadow communities.Biodiversity Science,2004,12(4):403-409.(in Chinese)
[39] 杜国祯,覃光莲,李自珍,刘正恒,董高生.高寒草甸植物群落中物种丰富度与生产力的关系研究.植物生态学报,2003,27(1):125-132. Du G Z,Qin G L,Li Z Z,Liu Z H,Dong G S.Relationship between species richness and productivity in an alpine meadow plant community.Acta Phytoecologica Sinica,2003,27(1):125-132.(in Chinese)
[40] 鲍根生,王宏生.甘肃马先蒿对高寒地区几种优良牧草的化感作用.中国草地学报,2011,33(3):88-94. Bao G S,Wang H S.Allelopathic effects ofPediculariskansuensisMaxim.on several graminaceous grass species on alpine meadow.Chinese Journal of Grassland,2011,33(3):88-94.(in Chinese)
[41] 肖金玉,蒲小鹏,徐长林.禁牧对退化草地恢复的作用.草业科学,2015,32(1):138-145. Xiao J Y,Pu X P,Xu C L.Effects of grazing prohibition on restoration of degraded grassland.Practacultural Science,2015,32(1):138-145.(in Chinese)
(责任编辑 武艳培)
Effect of fences on functional groups and stability of the alpine meadow plant community in the Qinghai-Tibet Plateau
Luo Jiu-fu1,2, Zhou Jin-xing2, Zhao Wen-xia1, Dong Lin-shui3, Zheng Jing-ming1
(1.College of Forestry, Beijing Forestry University, Beijing 100083, China;2.Key Laboratory of State Forestry Administration on Soil and Water Conservation, Beijing Forestry University,Beijing 100083, China; 3.Key Laboratory of Eco-environmental Science for Yellow River Delta,Shandong Province; Binzhou University, Binzhou 256603, China)
The changes in community structure and stability of the alpine meadow ecosystem under artificial conditions are an important way to study its ecological function. To explore this kind of change in the Qinghai-Tibet Plateau, field investigations within and outside of fences in a permanent sample plot (established in 2005) in Wumatang were conducted in August of 2009 and 2013. Species richness(R), differences in functional groups, and M. Godron stability index were analysed to explore the differences between fenced and unfenced communities. The results showed that 1) the species richness of the fenced community was greater than unfenced; the number of annual and biennial plants in the fenced community was significantly greater than in the unfenced one in 2009; the dominant species of the fenced community changed fromKobresiapygmaeatoPediculariskansuensisby 2013, and species belonging to perennial grasses, forbs, and annuals and biennials meant the fenced community was obviously richer than the unfenced community; and 2) the M. Godron stability index showed that the fenced community was more stable than the unfenced one in 2009, but less stable than the unfenced community in 2013. The fenced community structure showed an obvious change and the stability presented a downtrend as the time went by, whereas the stability of the unfenced community presented an uptrend.
alpine meadow; functional group; M. Godron; degradation; poisonous weeds;Kobresiapygmaea;Pediculariskansuensis
Zheng Jing-ming E-mail:zhengjm@bjfu.edu.cn
10.11829/j.issn.1001-0629.2016-0302
2016-06-03 接受日期:2016-08-30
国家林业局公益性行业科技专项(201504401);国家自然科学基金(30870231)
罗久富(1990-),男,四川绵阳人,在读博士生,主要从事恢复生态学研究。E-mail:yijiuyuan@yeah.net
郑景明(1971-),男,黑龙江鸡西人,副教授,博士,主要从事恢复生态和入侵生态研究。E-mail:zhengjm@bjfu.edu.cn
S812
A
1001-0629(2017)3-0565-10*
罗久富,周金星,赵文霞,董林水,郑景明.围栏措施对青藏高原高寒草甸群落结构和稳定性的影响.草业科学,2017,34(3):565-574.
Luo J F,Zhou J X,Zhao W X,Dong L S,Zheng J M.Effect of fences on functional groups and stability of the alpine meadow plant community in the Qinghai-Tibet Plateau.Pratacultural Science,2017,34(3):565-574.
