转染HBV Creg DNA的小鼠肝细胞表面干性分子表达观察
2017-03-13罗昊翔顾隆贺礼红姜文敏李佳佳黔西南民族职业技术学院医药系贵州兴义562400
罗昊翔,顾隆,贺礼红,姜文敏,李佳佳(黔西南民族职业技术学院医药系,贵州兴义562400)
转染HBV Creg DNA的小鼠肝细胞表面干性分子表达观察
罗昊翔,顾隆,贺礼红,姜文敏,李佳佳
(黔西南民族职业技术学院医药系,贵州兴义562400)
目的 观察转染pcDNA3.1-HBV Creg DNA质粒的小鼠肝细胞表面干性分子CD44、CD90、CD133的表达变化。方法 从含有pcDNA3.1-HBV Creg DNA质粒的Ecoli DH5α菌株中提取pcDNA3.1-HBV Creg DNA质粒。取SPF级正常小鼠的肝细胞,常规培养,待细胞贴壁后,加入50 μL pcDNA3.1-HBV Creg DNA混合转染细胞。采用免疫细胞化学染色法检测转染前后小鼠肝细胞表面干性分子CD44、CD90、CD133表达。结果 转染前,部分小鼠肝细胞CD90、CD133呈阳性,且阳性染色细胞都是圆形。未见有CD44阳性染色细胞。转染pcDNA3.1-HBV Creg DNA后,小鼠肝细胞表面干性分子CD44、CD90、CD133呈阳性染色。结论 转染pcDNA3.1-HBV Creg DNA质粒的小鼠肝细胞表达CD44、CD90、CD133。HBV Creg DNA可能影响小鼠肝细胞表面干性分子CD44的表达。
肝癌;肝肿瘤;乙肝相关性肝癌;干细胞;干性分子
HBV Creg DNA(nt1087-nt2488)是存在于乙型肝炎病毒(HBV)基因组中一段DNA片段,包括HBV的核心基因、两个增强子(Enh Ⅰ和EnhⅡ)、完整的X蛋白的启动子、核心启动子、反式调控序列(X蛋白编码区)、信号肽序列(前C编码区),即包含了HBV核心基因及其上游的调控序列和包装信号区(1853nt-1982nt)、顺向重复序列1(DR Ⅰ)、顺向重复序列2(DR Ⅱ)及其间的黏性末端区(1592nt-1836nt)。目前,肿瘤干细胞的研究领域尚存在较多争议。Reya等[1]提出了肿瘤干细胞假说。也有研究认为,肿瘤是由于干细胞发育异常导致的。目前,临床尚无公认的筛选肿瘤干细胞的特异性标记分子。CD90、CD133和CD44是最常见的肝癌干细胞标记分子,既往研究发现其在肝癌细胞中高表达[2],但CD90、CD133和CD44不是特异性的肝癌干细胞标记分子,正常组织和肿瘤组织中均可见其表达。乙肝相关性肝癌是我国常见的肝癌类型,HBV DNA可调节肝细胞基因的表达。但HBV是否会导致肝细胞表面干性分子的表达变化呢?我们观察了观察转染pcDNA3.1-HBV Creg DNA质粒的小鼠肝细胞表面干性分子CD44、CD90、CD133的表达变化,旨在为乙肝相关性肝癌的分子靶向治疗提供理论依据。
1 材料与方法
1.1 实验动物、试剂及仪器 2~3周龄SPF级正常小鼠1只,采用国标准啮齿类动物干饲料喂养,自由饮水进食。RPMI1640(Invitrogen Corporation),小牛血清(杭州四季青公司),CD44抗体一抗、CD90抗体一抗、CD133抗体一抗、免疫组化试剂盒(武汉博士德生物工程有限公司),胰蛋白酶(Biosharp),质粒提取试剂盒(AXYGEN),LipofectamineTM2000(Invitrogen Corporation),菌株Ecoli DH5α(Fermentas),琼脂,胰蛋白胨,酵母提取物。
1.2 pcDNA3.1-HBV Creg DNA质粒提取 将含有pcDNA3.1-HBV Creg DNA质粒的Ecoli DH5α菌株接种于含100 μg/mL Amp的LB液体培养基中,37 ℃电热培养箱中过夜培养,取出试管,按AXKGEN质粒提取试剂盒说明书提取质粒。
1.3 小鼠肝细胞分离、培养与 pcDNA3.1-HBV Creg DNA质粒转染方法 取SPF级正常小鼠1只,颈椎脱臼处死,取出肝脏,使用解剖剪将小鼠肝脏充分剪碎,使小鼠肝细胞脱落,形成肝细胞悬液。吸取肝细胞悬液,加入25 mL细胞培养瓶中,置于37 ℃、5% CO2的CO2培养箱中培养。培养的小鼠肝细胞在低倍镜下观察,细胞形态多种多样,大部分细胞呈圆形、椭圆形,少数细胞呈梭形。细胞颜色深浅不一。细胞越幼稚,细胞越小,颜色越浅。待细胞贴壁后,弃上清,加入适量0.01 mmol/L PBS冲洗2~3次,直至倒置显微镜下观察无悬浮细胞,将50 μL pcDNA3.1-HBV Creg DNA溶于500 μL无血清RPMI1640中,混匀,加入10 μL LipofectamineTM2000,混匀,室温放置25 min,将混合物加入培养瓶中,再加适量RPMI1640培养基,继续培养至第二天胰蛋白酶消化传代,传代时,一部分传于盖玻片上,置于37 ℃、5%CO2的CO2培养箱中培养过夜,第2天将传有细胞的盖玻片置入10%甲醛溶液中固定12 h,备用。
1.4 转染前后小鼠肝细胞表面干性分子CD44、CD90、CD133检测 采用免疫细胞化学。从甲醛中取出转染前、转染后的传有小鼠肝细胞的盖玻片各3片,分别使用0.01 mmol/L PBS洗2 min×3次,分别滴加H2O2适量,室温下静置30 min、0.01 mmol/L PBS洗2 min×3次,滴加1滴5%BSA室温封闭20 min,甩干盖玻片多余液体,不洗,分别滴加一抗(鼠抗人CD44抗体、鼠抗人CD90抗体和鼠抗人CD133抗体),37 ℃水浴2 h,取出,0.01 mmol/L PBS洗2 min×3次,滴加辣根过氧化物酶标记的兔抗鼠二抗,37 ℃水浴30 min,0.01 mmol/L PBS洗2 min×3次,滴加SABC试剂1滴,37 ℃水浴20 min,0.01 mmol/L PBS洗5 min×4次,显色剂显色后,普通光学显微镜下观察转染前、后小鼠肝细胞表面干性分子CD44、CD90、CD133的表达情况,阳性细胞可被染色剂染为蓝绿色,阴性细胞不显色。
2 结果
转染前小鼠肝细胞表面干性分子CD44、CD90、CD133表达情况见图1。转染前镜下可见部分细胞CD90、CD133呈阳性,且阳性染色细胞都是圆形,细胞个体较小。CD44的检测结果一直都是阴性,未见有CD44阳性染色细胞。
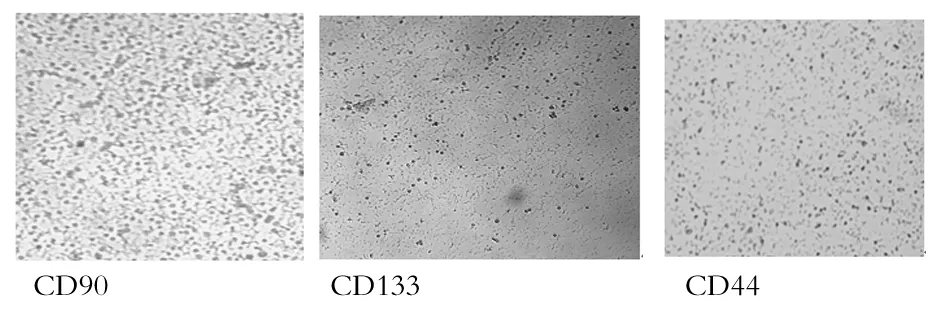
图1 转染前小鼠肝细胞表面干性分子CD44、CD90、CD133表达
转染后小鼠肝细胞表面干性分子CD44、CD90、CD133表达情况见图2。在肝细胞转染pcDNA3.1-HBV Creg DNA后,部分细胞CD44呈阳性染色。
3 讨论
有研究人员认为,肿瘤的发生是干细胞发育受阻,激活了异常分裂机制,导致细胞恶性增殖。也有研究人员认为,肿瘤组织中存在突变的干细胞,这些不正常的干细胞只能发育为肿瘤细胞,而不能发育为正常组织细胞,这是肿瘤发生及复发转移的根本原因[3]。正常组织的干细胞与肿瘤组织的干细胞到底有何不同呢?干细胞是一种前体细胞,细胞的某种干性分子不只是在一种组织存在,在很多种组织可能存在相同的干性标记分子,如CD133在造血干细胞、肝脏干细胞、肾细胞癌、乳腺癌等均有表达,所以找到特异性的肿瘤干细胞表面分子标记,对于肿瘤的分子靶向治疗有重要意义。本实验研究主要通过将外源因素pcDNA3.1-HBV Creg DNA导入正常小鼠肝细胞,观察正常小鼠肝细胞表面干性标记分子的变化,从而找到乙肝相关性肝癌特异性的细胞表面分子标记,为乙肝相关性肝癌的分子靶向治疗提供理论依据。

图2 转染后小鼠肝细胞表面干性分子CD44、CD90、CD133表达
pcDNA3.1-HBV Creg DNA为我们实验室构建的HBV DNA片段的表达载体,该HBV Creg DNA片段中,包含了HBV的核心基因、两个增强子(Enh I和EnhⅡ)、完整的X蛋白的启动子、核心启动子、反式调控序列(X蛋白编码区)、信号肽序列(前C编码区)即包含了HBV核心基因及其上游的调控序列和包装信号区(1853nt-1982nt)、顺向重复序列1(DRⅠ)、顺向重复序列2(DR Ⅱ)及其间的粘性末端区(1592nt-1836nt)。研究表明,HBV的该段DNA序列,与肝癌的发生密切相关。HBV X蛋白可以反式调控、激活细胞信号传导通路,引起表观遗传改变[3],从而提高细胞干性表达,决定肿瘤转化。HBV两个直接重复序列区域可以同源重组等多种方式整合入人类肝细胞基因组[4],通常整合入细胞基因组的HBV DNA序列还包括X基因、X基因的启动子、C基因、C基因的启动子、增强子2(EnhⅡ)、S基因等。X基因的启动子是已证明了的强启动子,整合后能够有效激活细胞基因的表达,EnhⅡ增强表面抗原和X基因启动子的转录活性[5],整合后导致HBV的X抗原和表面抗原持续表达,共同调节细胞基因的转录,激活干细胞的信号传导途径,使细胞表现出干性。此外,HBV基因整合入细胞基因组可能会导致p53基因失活,使干细胞基因过表达[6]。在小鼠肝细胞转染pcDNA3.1-HBV Creg DNA前后,细胞表面干性标记分子发生了变化,转染前,在小鼠肝细胞表面主要检测出的细胞干性标记分子为CD133和CD90部分细胞呈阳性表达,而CD44在转染前一直未检出。转染后,在小鼠肝细胞表面也能检出CD133和CD90部分细胞呈阳性表达,但CD44在转染后,检出大部分小鼠肝细胞呈阳性表达。CD133和CD90在正常组织和肿瘤组织均有实验研究检出,而对于CD44的检测,大部分实验研究都是检测肿瘤组织[7~11]。
众所周知,肝细胞属于稳定细胞。但是当肝脏受到刺激时,肝细胞既表现出较强的再生能力,肝细胞的这种再生能力说明肝脏内存在维持细胞更新的干细胞[12]。正常肝脏细胞可能不具有干性特征,但当肝脏受到损伤刺激后,肝细胞则表现出干性特征,经过损伤刺激,肝细胞表面出现干性标记分子,而我们的研究是通过取出小鼠肝脏,然后进行细胞培养,中间经过肝脏损伤刺激的过程,所以在肝细胞传代培养后,检测肝细胞部分细胞干性标记分子阳性。Rountree等[13]的研究表明分离出CD133+肝脏干细胞可以体外克隆扩增,CD133是一个干细胞标记分子[14],CD90在胚胎肝脏和肝肿瘤中也是一个很重要的干性标记[15],但是我们研究表明,CD90和CD133分子在正常干细胞和pcDNA3.1-HBV Creg DNA转染后的均有表达,说明CD90和CD133分子可能是正常肝脏干细胞表面的干性标记分子。正常肝细胞在pcDNA3.1-HBV Creg DNA转染前为检测出CD44的表达,然而,在转染后却检测出有大部分肝细胞呈阳性表达,说明带有调控序列的HBV Creg DNA片段可影响小鼠肝细胞表面干性分子CD44的表达,CD44分子的阳性表达可能与乙肝相关性肝癌的发生有关,CD44分子可能是比较有特性的肝癌干细胞的表面干性标记分子。
综上所述,转染pcDNA3.1-HBV Creg DNA质粒的小鼠肝细胞表达CD44、CD90、CD133。HBV Creg DNA可可影响小鼠肝细胞表面干性分子CD44的表达。CD44可能是乙肝相关性肝癌干细胞的干性标记分子,而CD90、CD133可能是正常肝脏干细胞的干性标记分子。
[1] Reya T, Morrison SJ, Clarke MF, et al. Stem cells,cancer,and cancer stem cells[J]. Nature, 2001,414(6859):105-111
[2] Bahnassy AA, Fawzy M, El-Wakil M, et al. Aberrant expression of cancer stem cell markers (CD44, CD90, and CD133) contributes to disease progression and reduced survival in hepatoblastoma patients: 4-year survival data[J]. Transl Res, 2015,165(3):396-406.
[3] Tian Y, Ni DJ, Yang WB, et al. Telbivudine treatment corrects HBV-induced epigenetic alterations in liver cells of patients with chronic hepatitis B[J]. Carcinogenesis, 2014,35(1):53-61.
[4] Kimbi GC, Kramvis A, Kew MC. Integration of hepatitis B virus DNA into chromosomal DNA during acute hepatitis B[J].World J Gastroenterol, 2005,11(41):6416-6421.
[5] Bai X, Zhu Y, Jin Y, et al. Temporal acquisition of sequential mutations in the enhancer II and basal core promoter of HBV in individuals at high risk for hepatocellular carcinoma[J]. Carcinogenesis, 2011,32(1):63-68.
[6]Amaddeo G, Cao Q, Ladeiro Y, et al. Integration of tumour and viral genomic characterisations in HBV-related hepatocellular carcinomas[J]. Gut, 2014,64(5):820-829.
[7] Jing FF, Kim HJ, Kim CH, et al. Colon cancer stem cell markers CD44 and CD133 in patients with colorectal cancer and synchronous hepatic metastases[J]. Intern J Oncol, 2015,46(4):1582-1588.
[8] Huang X, Sheng Y, Guan M. Co-expression of stem cell genes CD133 and CD44 in colorectal cancers with early liver metastasis[J].Surg Oncol, 2012,21(2):103-107.
[9] Chen KL, Pan F, Jiang H, et al. Highly enriched CD133(+)CD44(+) stem-like cells with CD133(+)CD44(high) metastatic subset in HCT116 colon cancer cells[J].Clin Exp Metas, 2011,28(8):751-763.
[10] Ying H, Zou QF, Ge RL, et al. The critical role of CD133+CD44+/high tumor cells inhematogenous metastasis of liver cancers [J]. Cell Res, 2012,22(1):259-272.
[11] Wang CX, Xie JP, Guo JS, et al. Evaluation of CD44 and CD133 as cancer stem cell markers for colorectal cancer[J].Oncolg Rep, 2012,28(4):1301-1308.
[12] Jeong Eun H, Sang HH, Ji-Hun Km, et al. Distribution of hepatic stem cell markers in human liver with massive hepatic necrosis[J]. Basic Applied Pathol, 2010,3(2):39-45.
[13] Rountree CB, Ding W, Dang H, et al. Isolation of CD133+Liver Stem Cells for Clonal Expansion[J]. J Vis Exp, 2011, 2011 (56):1-9.
[14] Zhong L. CD133: a stem cell biomarker and beyond[J]. Exper Hematol Oncol, 2013,2(17):1-8.
[15] Ceafalan L, Vidulescu C, Radu E, et al. Expression of stem cell markers on fetal and tumoral human liver cells in primary culture[J]. Rev Med Chir Soc Med Nat Iasi, 2005,109(1):96-104.
贵州省科技厅科学技术基金资助项目(2013-2268) 。
罗昊翔
10.3969/j.issn.1002-266X.2017.07.011
R73
A
1002-266X(2017)07-0037-03
2016-06-12)
