牛肠道病毒的研究进展
2017-02-15何欢张斌
何欢,张斌
(西南民族大学生命科学与技术学院,成都 610041)
牛肠道病毒的研究进展
何欢,张斌*
(西南民族大学生命科学与技术学院,成都 610041)
国内近几年牛肠道病毒病已经开始逐渐流行,现已发现多株新型病毒株并对此建立了多种分子及血清型方面的检测方法。通过对牛肠道病毒的基本结构、鉴定方法、检测方法、流行情况以及致病机制进行详细的介绍,以期为更好的研究牛肠道病毒在我国的流行现状及防治措施提供依据。
牛肠道病毒;分离鉴定;检测方法;发病机制
牛肠道病毒(bovine enterovirus, BEV)广泛存在于人类和各类动物机体内并且能够引起其呼吸道系统和消化系统的疾病,一般认为BEV是牛肠道中原有的栖居者,在肠道中增殖,病原性不明显。BEV的致病性有些许争议,有认为只引起轻度腹泻或健康牛的隐性感染,也有认为可引起呼吸系统和消化系统疾病[1],其致病性机理还不是很清楚。本文对牛肠道病毒的研究进展作一综述。
1 BEV的概述
1.1 形态、结构及功能 牛肠道病毒为小RNA病毒科肠道病毒属成员之一,为无囊膜的单股正链RNA病毒,具有典型的二十面体立体对称的球形结构(图1)。早期报道过BEV的晶体结构并发现牛肠道病毒衣壳蛋白的拓扑和病毒粒子整体结构与小RNA病毒科其他成员一样[3]。病毒粒子大小为25~30 nm,如图1C所示为牛肠道病毒的电镜图片[4],全长7300~7500 bp,基因组中仅有一个开放阅读框(ORF),可直接作为mRNA 翻译出一个多聚蛋白,经过一系列降解,产生4 种结构蛋白(VP1、VP2、VP3 和VP4)和7种非结构蛋白(2A、2B、2C、3A、3B、3C 和3D),如图1D所示为肠道病毒基因结构的示意图[5]。VP1、VP2、VP3与VP4蛋白包绕基因组RNA组成病毒的基本结构,而7种非结构蛋白虽不能构成病毒的主要结构,但在基因复制、转录、翻译等过程中提供重要的酶类等物质,以保障病毒的正常繁殖代谢。在其两侧为5′和3′非编码区(UTRs),在3′非编码区的末端含有一个长度可变的多聚腺苷酸尾巴(poly-A)[6]。病毒衣壳是由VP1、VP2、VP3 和VP4构成,VP4是被包埋在病毒衣壳内部的,一般感染机体不会产生相应的抗体,而其他VP1、VP2和VP3 三种结构蛋白暴露在病毒的表面,是刺激机体产生抗体的主要部位。在VP1蛋白上有最主要的抗原抗原结合位点,VP2、VP3和VP4蛋白与病毒衣壳的理化特性及某些稳定性有关又是其重要的组成部分。非结构蛋白中的各种蛋白多数与酶的活性有关,如2A具有半胱氨酸蛋白酶活性,2C具有ATP酶活性;也有在RNA的复制过程中起到重要作用的蛋白,如3B在与RNA 5′末端的pUpU形成磷酸二酯键之后启动RNA的合成,2B和3A参与并促进RNA早期的复制,随后在3D蛋白的作用下引起病毒RNA的复制并在3C水解酶蛋白的促进作用下可导致宿主细胞的死亡[7]。牛肠道病毒其它肠道病毒相比,在内部核糖体进入位点(IRES)与三叶草结构之间多了100个核苷酸[8]。5’UTR可通过与细胞蛋白因子的相互结合从而调控病毒基因组的合成及蛋白翻译,VPg蛋白结合在5'末端之后与聚合酶3D相互作用,启动病毒RNA的复制[6]。3'UTR末端含有的(poly A)与病毒的感染性、基因组复制及mRNA的合成效率相关[6]。
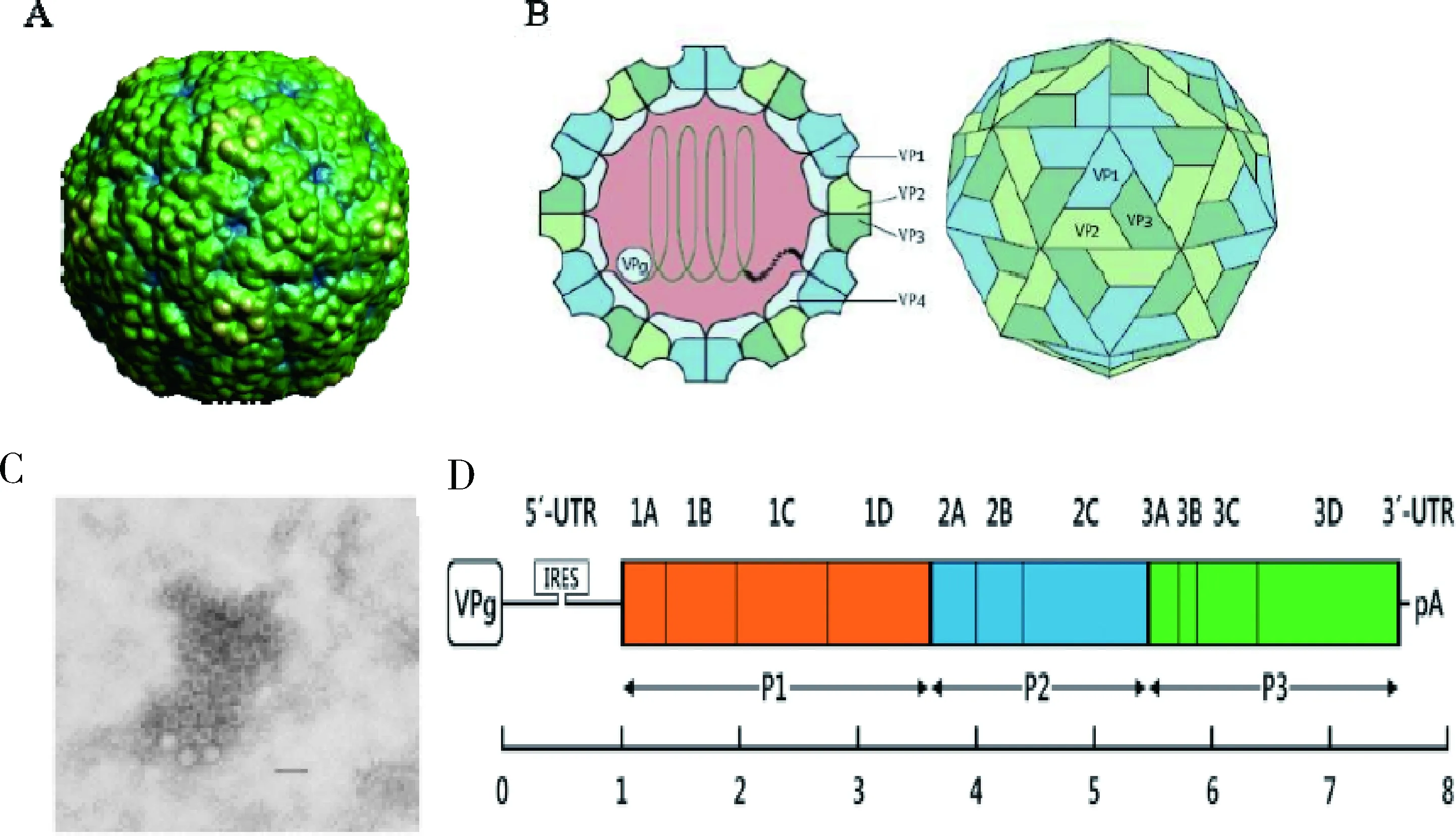
A:牛肠道病毒粒子模拟结构;B:肠道病毒粒子剖面图;C:牛肠道病毒的电镜图片(100000×);D:肠道病毒基因结构示意图图1 肠道病毒粒子及衣壳结构
BEV粒子无囊膜,故对乙醚、脱氧胆酸盐、去污剂以及氯仿有一定的抵抗力,也能抵抗蛋白水解酶的作用。此外牛肠道病毒还能抵抗弱酸条件如pH3.0、常见的消毒剂如70%乙醇和5%甲酚皂溶液等。这些理化特性决定了该病毒可在自然环境中存活数月之久。但牛肠道病毒在碱(pH10.0 以上)、高温(50 ℃以上,钙、镁离子存在时除外)和紫外线照射等条件下1 h左右便可被灭活[9]。1.2 BEV的分类及鉴定方法 BEV的分子型根据5′UTR和衣壳蛋白的结构分为BEV-E(原BEV-A)和BEV-F(原BEV-B)两大类。在BEV检测分型过程中病毒分离物或原始标本首先以5′UTR鉴定为小RNA病毒科病毒。VP1蛋白是最主要的衣壳蛋白,具有最多的型特异性中和位点,因此VP1 蛋白编码基因序列与病毒血清型具有较高的相关性,通过扩增全长VP1序列,构建系统进化树对比VP1区核酸同源性与已知血清型肠道病毒同源性<70%(氨基酸同源性<85%)则认为所测病毒与标准株不是同一血清型,为一新血清型[3]。VP1作为BEV分型的主要依据已经被普遍认可,但随着VP1序列数据的不断扩大,部分毒株的情况不符合70%/85%的分类标准,比如获得的VP1序列不完整,则要获得更多的信息如P1全序列、中和试验结果及免疫电镜图片等来进行综合判定。
BEV血清型的分型经历了几次重要的变动。1985年Knowles等[10]将BEV的分型标准确定为2个血清型(BEV-1和BEV-2)。2006年Zell等[11]通过对已知的牛肠道病毒的 5′UTR、VP1、VP2、VP3、VP4和3D 的序列进行了比对分析后确定了该病毒其实可以分为BEV-A 与 BEV-B两个大群。目前,世界公认的BEV的分类标准是在第九次病毒委员会分类报告[12]上公布的EV-E(即BEV-A)与 EV-F(即BEV-B)两种,前者包括E1~E4共四种基因型,后者分括F1~F7共六种基因型。
2 BEV的流行现状
一般认为BEV是牛肠道中原有的栖居者,在肠道中增殖,病原性并不明显。大多数人认为牛肠道病毒可引起呼吸道疾病、繁殖障碍和流产等临床症状,BEV在发生多种临床症状(呼吸系统、消化系统、繁殖障碍性疾病)牛群中都得到了分离,甚至是假定健康的牛群动物中也能够得到分离[13]。
2.1 国外的流行现状 自1959年Moll和Davis首次分离到BEV以来[14],全世界很多国家和地区先后报道了BEV的感染情况,在国外牛群中的流行率约为17.6%~80%[15]。2002年Ley等[16]研究发现与封闭牛群畜牧业相关的环境,包括加拿大白尾鹿、加拿大鹅、动物的饮用水源和牧场下游的临近河牡蛎都能够通过RT-PCR等方法检测的到BEV的存在,其中白尾鹿粪便的阳性率为38%,牛粪便的阳性率为76%。并对所检测的BEV进行序列分析发现并不完全属于同一型别。2007年Zheng等[17]从澳大利亚刷尾负鼠体内肠道内容物中分离得到两株牛肠道病毒W1与W6,均属于BEV-2型。2008年S Gur等[15]对土耳其部分地区不同物种BEV-1感染情况进行了血清学检测,其中包括244份人血清、1520份牛血清,检测结果表明人血清阳性率为30.3%,牛血清阳性率为64.8%,这是首次在人血清中检测到BEV的抗体,但其是否能够引起人类的一些临床症状尚未可知。2012年A′kos Boros等[13]从健康羊粪便样本中利用5′UTR和VP1基因序列鉴定并分离出了一株自然种间重组的猪/牛肠道病毒OEV-1,同时也首次获得了该毒株的全基因序列。2013年McClenahan等[4]在因呼吸道疾病致死的羊驼肺组织样品中首次分离得到一株BEV,经后续试验鉴定其为EV-F。2015年Sobhy等[18]从严重腹泻的牛体内分离到了一株BEV-F2型。
2.2 国内的流行现状 在国外,BEV的流行及传播比较普遍,但在中国有关BEV疾病的报告却还没有系统的数据分析。但近几年已多次报道新型BEV的发现,比如2011年李英利等[19]报道过从内蒙古地区的犊牛粪便中分离出了1株BEV,命名为BHM26,并且分析表明中国BEV分离株与国外参考毒株具有较大差异但未鉴定其基因型;2013年侯佩莉等[20]从山东地区的泌乳奶牛的腹泻粪便样本中分离出了BEV,命名为BEV-SD。同年,彭小薇等[21]从北京某牛场的泌乳奶牛的腹泻样本中分离获得1株牛肠道病毒2型,命名为BJ001,之后又对分离株进行了全基因组测序分析;2014年从吉林省一个发病率和死亡率较高的牛场分离到三株E种BEV,命名为HY12[22],从北京地区某牛场分离到一株F种BEV[23];2015年张海丽等[24]从山西某奶牛场的泌乳奶牛粪便样品中分离得到一株BEV。
从以上数据可以初步分析国内外BEV均有E型和F型的毒株出现,同时国内的分离株多数是属于新型分离株,也丰富了BEV属的种类。该病毒在国内外的流行差异主要在于感染物种的不同,国内现在仅在反刍动物体内监测的到该病毒的存在,如牛、羊等;而国外的感染谱相对比较广泛,除了牛、养外,还可在羊驼、鹿、猪、马、鹅、鼠,甚至在人类的血清中都能够监测的到该病原的存在。对此,在我国未来的牛肠道病毒的研究发展中应该扩大检测的物种范围,更加全面系统的监测BEV在我国的感染谱及流行率。
3 BEV的检测技术的发展
通常多数研究者认为牛肠道病毒的病原性不明显,广泛存在于自然界中,其宿主范围较广,能够在一定条件下如侵入其他组织或是继发感染和混合感染其他病原时引起显著的症状进而导致动物疾病的发生,造成经济损失[12],因此简便快速的检测技术对BEV的监控和防治是非常必要的。
目前已报道的牛肠道病毒的检测技术主要有传统的电镜检查、病毒分离、血清学试验和TaqMan荧光定量PCR等。2005年Jimenez-Clavero等[25]针对BEV 5′UTR基因建立了相关的RT-PCR检测方法并对西班牙的多个地区的牛、绵羊、山羊、驴、马以及地表水样进行了检测,结果数据表明,除了在驴的检测样本中未检测到,其余均有阳性样本存在;2016年Nathamon等[26]根据5′UTR基因设计引物(长约290 bp)建立了RT-PCR的方法并对泰国部分地区的本地牛、印度牛及山羊携带BEV的感染率做了一个调查和分析,结果表明,以上几种动物均有BEV的携带,其中本地牛的携带率最高(76%)。2012年我国学者吴丹等[27]根据5′UTR基因建立了检测BEV的TaqMan定量PCR方法,并对大量的临床样本进行了临床检测,检测结果表明该检测方法具有较好的特异性而且敏感性比病毒分离法高10倍。2015年朱彤[28]和2014年侯佩莉等[20]分别针对其3D 基因建立了RT-PCR和TaqMan定量PCR方法,两者所建立的该方法与引起牛的病毒性腹泻的其它几种牛的病毒(牛病毒性腹泻病毒(BVDV)、牛轮状病毒(BRV)、牛传染性鼻气管炎病毒(IBRV)、牛冠状病毒(BCoV)等)均无交叉反应,前者所建立的方法检出敏感度达7.13×10-1拷贝/μL,比常规PCR检测方法高10倍,后者所建立的检测方法的敏感度也较高,达10-1TCID50。血清学检测试验发展较缓慢,最近几年有所增多,在2014年郭金玉等[29]利用北京某牛场牛肠道病毒2型(BEV-2) 分离毒株的VP1蛋白构建了BEV抗体的间接ELISA检测方法。2015年朱彤等[28]通过扩增BEV的VP2基因并进行一系列的转化、诱导表达、纯化出重组蛋白制备了较高效价的多克隆抗体,为BEV的血清学诊断方法的建立及亚单位疫苗的研制奠定了基础。2016年邢泽黎等[30]应用大肠杆菌表达纯化的E种肠道病毒的 VP1和VP2重组蛋白制备了抗体,并建立了检测BEV病原的双抗体夹心ELISA 方法,该检测BEV的抗原方法具有特异性、灵敏度高和快速等特点,并对吉林省不同地区的牛群感染BEV进行了病原流行病学调查,发现不同地区牛群的BEV感染率为8.16%~58.7%。
4 BEV致病机制的研究现状
BEV的致病性有些争议,有认为只引起轻度腹泻或健康牛的隐性感染,致病性弱,也有认为可引起呼吸系统疾病、腹泻、产奶量下降、睾丸炎、包皮炎、黏膜炎、流产、死胎、盲肠结肠炎等临床症状[13],这可能是由于不同血清型的病毒对不同组织的嗜性不同而引起的疾病也不同,其致病性机理还不是很清楚。由于其临床症状在临床实验动物模型中很难被复制,因而BEV和机体之间的相互作用、致病机理等相关研究仍处于初步研究的阶段。
1980年Yilma等[31]研究BEV感染细胞后,通过免疫荧光抗体标记后观察病毒感染细胞的过程,来研究牛肠道病毒组装和释放的形态变化及发生过程,证实病毒通过囊泡释放到细胞外,部分也存在于细胞质泡中。2011年Blas-Machado等[32]第一次使用BEV-1株通过回归原动物实验的方法感染犊牛并观察BEV-1的致病机制,结果表明,感染组中观察到了一定程度的腹泻、呼吸窘迫及厌食等相似的临床症状,但在对照组中也出现了腹泻等现象,由于缺乏严格的对照动物导致该实验不能够准确的分析其致病机理,但其在某种程度上也可说明BEV-1株能够感染犊牛并且可能具有与其它肠道病毒类似的致病机制。2014年Zhang等[33]获得了新型BEV的分离株并利用50 μL/只的病毒量腹腔注射刚出生1d的乳鼠,结果表明可在其心、肝、肺、肠道以及排泄物中检测的到病毒的存在,说明该病毒能够引起呼吸道和消化道的感染。2015年盖小春等[34]也利用BEV-E种的HY12建立了小鼠的动物感染模型,结果表明只有IRC系小鼠能够成功感染该病毒株,但昆明小鼠和BALB/c不能。小鼠成功感染病毒后其部分实质器官和肠道组织都能够出现程度不一的组织病理学的变化,如肺脏、肝脏、脑等器官内产生了大量的炎性细胞浸润,部分组织的结构消失,肠道上皮细胞脱落,肠壁增厚等等。免疫组织化学方法在机体的实质器官和肠上皮细胞中检测到了较多的BEV阳性细胞,表明该病毒株可以通过不同途径感染IRC系小鼠,并可以在不同的器官、组织中进行病毒复制。这些试验数据结果都为研究BEV的致病机制奠定了基础。
5 小结
BEV是国内近几年新发现的一种动物疾病,给养牛业造成了严重的经济损失,我们国内的牛群中既有E种牛肠道病毒也存在F种牛肠道病毒的感染,且两种血清型之间存在较大差异,给本病的分子流行病学的调查和确诊造成了一定的困难。同时,BEV粒子的大小、形态与细小病毒的极为相似,应用电镜观察也常常造成误诊。目前关于BEV疾病的诊断和防治还是处于缺乏状态。对于新型BEV的分离鉴定确定血清之后,多数BEV的致病机制、发病机理都还处于初步研究阶段,动物模型的建立也还不够完善,比如最佳的感染途径和剂量、动物病例的完整复制、病理组织的变化分析等发病机理还不够清楚。我国虽在近几年有关注BEV的动态,但国内还没有系统的统计和相对应的管理制度,因为BEV的感染可引起的临床症状如发热、咳嗽、腹泻等,也会由于和替他病毒的混合感染引起流产、产奶量下降等;而且有的病毒株是隐性感染,但也有些毒株可引起动物的死亡,因而关于BEV的致病性也一直存在着争议,对此,我们应该在今后的研究中努力完善这些不足。
[1] Nollens H H, Rivera R, Palacios G.,etal. New recognition of Enterovirus infections in bottlenose dolphins (Tursiopstruncatus) [J]. Veterinary Microbiology, 2009, 139(1/2): 170-175.
[2] Enterovirus E[EB/OL]. http://www.picornaviridae.com/enterovirus/ev-e/ev-e.htm.
[3] Oberste M S, Maher K, Nix W A,etal. Molecular identification of 13 new enterovirus types, EV79-88, EV97 and EV100-101, members of the species human enterovirus B[J]. Virus Research, 2007, 128(1/2):34-42.
[4] McClenahan S D, Scherba G, Borst L,etal. Discovery of a bovine enterovirus in alpaca[J]. PLoS One, 2013, 8(8): e68777.
[5] Jani Ylä-Pelto, Lav Tripathi and Petri Susi. Therapeutic use of native and recombinant enteroviruses[J]. Viruses, 2016, 8:57.
[6] Tuthill T J, Groppelli E, Hogle J M,etal. Picornaviruses[J]. Curr Top Microbiol Immunol, 2010, 343:43-89.
[7] Wu K X, Ng M M, Chu J J. Developments towards antiviral therapies against enterovirus 71[J]. Drug Discov Today, 2010, 15(23/24):1041-1051.
[8] Kung Y H, Huang S W, Kuo P H,etal. Introduction of a strong temperature-sensitive phenotype into enterovirus 71 by altering an amino acid of virus 3D polymerase[J]. Virology, 2010, 396(1): 1-9.
[9] 金奇. 医学分子病毒学[M].北京: 科学出版社, 2001: 606-613.
[10]Knowles N J, Barnett I. A serological classification of bovine enteroviruses[J]. Archives of Virology, 1985, 83(3/4):141-155.
[11]Zell R, Krumbholz A, Dauber M,etal. Molecular-based reclassification of the bovine enteroviruses[J]. Journal of General Virology, 2006, 87(2):375-385.[12]King A M, Adams M J, Lefkowitz E J,etal. Virus taxonomy: classification and nomenclature of viruses[A]. Ninth Report of the International Committee on Taxonomy of Viruses[M]. Elsevier, 2012.
[13]Boros A, Pankovics P, Knowles N J,etal. Natural interspecies recombinant bovine/porcine enterovirus in sheep[J]. The Journal of General Virology, 2012, 93:1943-1951.
[14]Moll T, Davis A A. Isolation and characterization of cytopathogenic enteroviruses from cattle with respiratory disease[J]. Am J Vet Res, 1959, 20: 27-32.
[15]Gür S, Yapkic O, Yilmaz A. Serological survey of bovine enterovirus type 1 in different mammalian species in Turkey[J]. Zoonoses Public Health, 2008, 55(2) : 106-111.
[16]Ley V, Higgins J, Fayer R. Bovine enteroviruses as indicators of fecal contamination[J]. Applied and Environmental Microbiology, 2002, 68(7):3455-3461.
[17]Zheng T. Characterisation of two enteroviruses isolated from Australian brushtail possums (Trichosurusvulpecula) in New Zealand[J]. Arch Virol, 2007, 152(1):191-198.
[18]Sobhy N M, Mor S K, Mohammed M E,etal. Isolation and molecular characterization of bovine enteroviruses in Egypt[J]. The Veterinary Journal, 2015, 12, 206(3):317-321.
[19]李英利, 常继涛, 于力. 牛肠道病毒的分离鉴定[J]. 中国预防兽医学报, 2011, (5):388-390.
[20]侯佩莉, 程洪兵, 刘晓, 等. 牛肠道病毒RT-PCR检测方法的建立及初步应用[J]. 山东农业农业科学, 2014, 46(2): 13-16.
[21]彭小薇, 董浩, 吴丹, 等. 牛肠道病毒2型的分离与鉴定[J]. 畜牧兽医学报, 2013, 44(5): 823-828.
[22]Zhu L,Xing Z, Gai X,etal. Identification of novel enterovirus E isolates HY12 from cattle with severe respiratory and enteric diseases [J/OL]. PLOS One, 2014, 9(5): e97730.
[23]Peng X, Dong H, Wu Q,etal. Full genome sequence of a bovine enterovirus isolated in China [J/OL]. Genome Announc, 2014, 2(3:) e00620-14.
[24]张海丽, 周萍萍, 高明春, 等.一株牛肠道病毒的分离与鉴定[J]. 中国奶牛,2015, 5:31-34.
[25]Jimenez-Clavero M A, Escribano-Romero E, Mansilla C,etal. Survey of bovine enterovirus in biological and environmental samples by a highly sensitive real-time reverse transcription-PCR [J]. Applied and Environmental Microbiology, 2005, 71(7): 3536-3543.
[26]Natham Kosoltanapiwat, Marnoch Yindee, Irwin Fernandez Chavez,etal. Genetic variations in regions of bovine and bovine-like enteroviral 5'UTR from cattle, Indian bison and goat feces[J].Virology Journal, 2016, 13:13.
[27]吴丹, 吴涛, 张鹤晓, 等. 牛肠道病毒TaqMan荧光定量RT-PCR检测方法的建立[J]. 中国预防兽医学报, 2012, 34(11): 903-906.
[28]朱彤, 赵贵民, 沈付娆, 等. 牛肠道病毒SYBR Green I实时荧光定量PCR检测方法的建立及初步应用[J]. 病毒学报, 2015, 31(5): 487-493.
[29]郭金玉, 高志强, 张鹤晓, 等. 牛肠道病毒2型VP1+表达和间接 ELISA方法的建立与应用[J]. 中国兽医杂志, 2014, 50(11): 40-43.
[30]邢泽黎, 王新平, 朱利塞, 等. 双抗体夹心ELISA 检测牛肠道病毒抗原方法的建立及流行病学调查[J]. 中国兽医学报,2016, 4(36): 567-571.[31]Yilma T, Breese S S. Morphogenesis of the assembly and release of bovine enterovirus[J]. J Gen Virol, 1980, 49(1): 225-230.
[32]Blas-Machado U, Saliki1 J T, Sa′nchez S,etal. Pathogenesis of a bovine enterovirus-1 isolate in experimentally infected calves[J].Veterinary Pathology, 2011, 48(6): 1075-1084.
[33]Haili Zhang, Hongtao Lia, Jun Bao,etal. Characterization of an Enterovirus species E isolated from naturally infected bovine in China[J]. Virus Research, 2014, (191):101-107.
[34]盖小春,邢泽黎,朱利塞, 等. E种肠道病毒HY12小鼠感染模型的建立[A].中国畜牧兽医学会动物传染病学分会第十六次学术研讨会论文集[C].济南:中国畜牧兽医学会动物传染病学分会出版社,2015:298.
(编辑:李文平)
Research Progress on Bovine Enterovirus
HE Huan, ZHANG Bin*
(CollegeofLifeScienceandTechnology,SouthwestUniversityforNationalities,Chengdu610041,China)
Bovine enterovirus has begun to gradually popular in recent years in China. It had discovered that many novel bovine enteroviruses, meanwhile there are lots of researchs on establishing of molecules and serotypes. In this study, the basic structures, isolation, detection methods, prevalence and pathogenesis of bovine enterovirus were introduced in detail, which provide a basis for further study of the epidemic status and control measures of bovine enterovirus in China.
bovine enterovirus; isolation; detection methods; pathogenesis
四川省科技计划项目青年基金(2014JQ0044);西南民族大学中央高校基本科研业务费专项资金项目(2016NGJPY10)
何欢,硕士研究生,从事动物病原分子生物的研究。
张斌。E-mail:binovy@sina.com
2016-10-21
A
1002-1280 (2017) 01-0063-06
S852.653
