乐清湾口海域春秋季甲壳动物的群落结构特征研究
2017-01-18夏陆军周青松俞存根宋伟华毕耜瑶李德伟
夏陆军, 周青松, 俞存根,宋伟华, 郑 基, 毕耜瑶, 谢 旭, 李德伟, 叶 深
乐清湾口海域春秋季甲壳动物的群落结构特征研究
夏陆军1, 2, 周青松3, 俞存根1,宋伟华3, 郑 基1, 毕耜瑶1, 谢 旭1, 李德伟1, 叶 深1
(1. 浙江海洋大学, 浙江舟山 316024; 2. 嵊四县海洋与渔业局, 浙江嵊四 202450; 3. 国家海洋局第二海洋研究所, 浙江杭州 310012)
根据2012年春季和10月在乐清湾口海域开展渔业资源调查所获甲壳动物资料, 对乐清湾口海域的甲壳动物的种类组成、优势种、物种数量分布、生物多样性和群落结构相似性等群落结构特征及其影响因素进行了研究。结果表明, 调查海域共鉴定出34种甲壳动物, 隶属于2目17科23属, 春季优势种仅有三疣梭子蟹1种, 秋季为日本、口虾蛄、中华管鞭虾、三疣梭子蟹4种, 两季优势种更替较显著; 秋季物种数较春季丰富, 调查水域中心区域物种数较多; 秋季甲壳动物生物多样性指数高于春季; 甲壳动物群落结构与海洋环境密切相关。乐清湾口海域的甲壳动物以季节性的广温低盐种类为主, 该海域位于河口, 受瓯江径流影响较大; 春季受瓯江径流淡水水团影响, 物种较少, 而秋季受交汇水团影响, 物种丰富; 春秋季组内并无明显规律, 地形、水深和水系等外部因素对甲壳动物群落结构影响较大。
甲壳动物; 生物多样性; 群落结构; 乐清湾口海域
乐清湾位于浙江南部沿海, 瓯江口北侧, 是葫芦状的半封闭性海湾, 与象山湾、三门湾并列为浙江3大半封闭海湾。该湾口有众多岛屿作屏障, 大大减小了外海风浪对湾内的影响, 湾内具有水质肥沃, 滩涂平坦稳定等有利的地形和气候条件, 加上瓯江径流不断注入大量营养物质, 促进了饵料生物孳生, 使该海域成为海洋经济动物索饵、产卵、繁育的场所, 尤其是十足目甲壳动物虾蟹类[1]的重要分布区之一。
有关浙江近海渔场的虾、蟹类与虾蛄类的资源状况、种类组成及其生物多样性方面的研究已有不少报道[2-8], 但对于浙江近岸海湾与河口的甲壳动物资源及其生物多样性方面的研究却不多见, 已有的研究报道主要有赵蒙蒙等[9]对三门湾海域口足目和十足目虾类的种类组成、时空分布及生物多样性的分析, 徐兆礼等[10]对瓯江口海域口足目与十足目资源分布特征的研究, 柏育材等[11]对舟山渔场衢山岛海域十足目和口足目分布特征的研究, 徐开达等[12]对中街山列岛水域甲壳动物资源及其生物多样性研究方面的研究, 齐海明等[13-15]对浙江椒江口海域春季虾类资源分布、群落结构以及与环境因子间关系的研究, 但迄今尚无针对乐清湾口海域甲壳动物群落结构及生物多样性的报道。
沿岸海域既是许多经济鱼、虾、蟹类的重要栖居场所, 是人类捕捞活动的重要渔场, 也是受海洋工程建设项目影响较大的区域, 大规模的人类活动必然对海域生态环境产生重大影响, 特别是在当前, 随着社会经济的发展、用海项目的大量增加, 填海造地等海洋工程项目建设造成大量渔业生物的生存空间被挤占, 洄游通道被切断, 产卵场被破坏等海洋渔业生态损害现象越来越严重。对乐清湾口海域甲壳动物群落结构的研究具有重要的理论价值和现实意义, 一方面可为乐清湾甲壳动物资源科学利用和可持续开发提供基础数据, 同时, 也可为海洋工程建设对海洋生态环境影响评价、渔业生态损害补偿等提供本底数据资料。
1 材料与方法
1.1 数据来源
文中所用甲壳动物数据来自2012年4月(春季)和10月(秋季)在乐清湾口的大小门岛海域进行的渔业资源定点拖网调查所得, 调查共设置16个站位(图1)。
渔业资源调查方法按照《海洋渔业资源调查规范》(SC/9403-2012)[16]进行调查船为浙乐休渔122号, 渔船主机功率为184 kW, 网具规格为460目× 60mm。调查时每站拖曳约1 h, 拖曳约为3.5节, 每站拖网所获的渔获物全部取样装入样品袋, 样品袋码记入渔捞记录中, 置于放在船舱内低温冰鲜保存, 样品鉴定分析在实验室内进行, 并分别对渔获物进行生物学测定, 称量使用电子天平, 精确度为0.1g。
1.2 数据处理及分析方法
1.2.1 相对重要性指数
甲壳动物优势种的计算采用相对重要性指数, 计算公式如下:
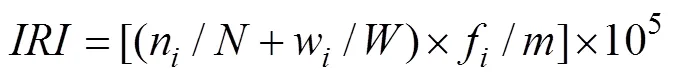
式中,n、w分别为第种生物的个体数和生物量;、分别为调查所获得的总个体数和总生物量;f 为第种生物在次取样中出现的频率;为取样次数。将相对重要性指数()大于1000者定为优势种, 在100~1000之间者定为常见种[17]。
1.2.2 群落生物多样性分析
采用Margalef种类丰富度指数()、Shannon- Wiener多样性指数()和Pielou均匀度指数()分析甲壳动物群落结构多样性, 其计算公式如下:
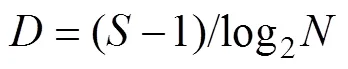
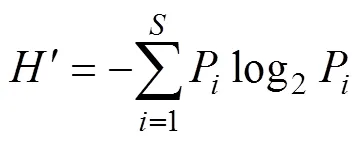
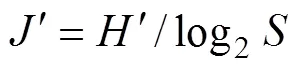
1.2.3 群落结构相似性分析
采用聚类分析方法对群落结构进行分析, 考察不同季节不同站位群落的相似性。对乐清湾口海域甲壳动物生物量进行4次方转换[14], 并删除了总体相对生物量小于1%的种, 但保留其中在任一站位相对生物量大于3%的种, 以减少机会种对群落结构的干扰, 平衡优势种和稀有种在群落中的作用, 使其更接近正态分布。在秋季调查4号站位时, 甲壳动物调查采样失败, 故不作具体分析。
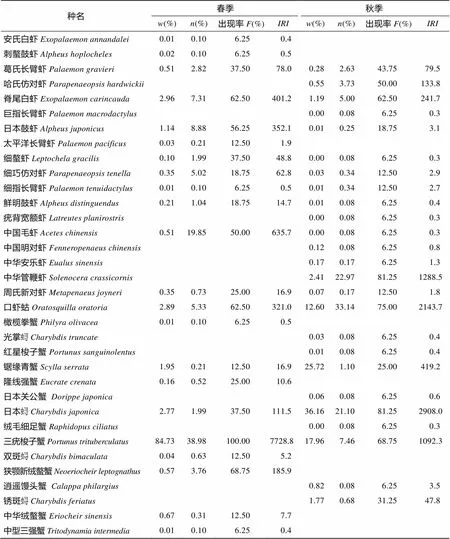
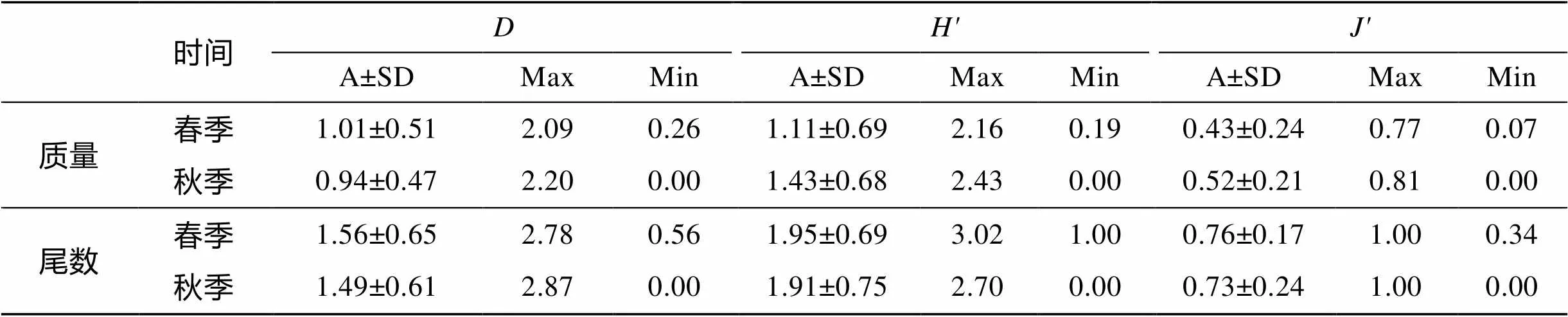
计算Bray-Curtis相似性系数矩阵, 采用非参数多维标序技术(NMDS)作二维标序与等级聚类分析(CLUSTER)来研究甲壳动物群落结构的季节变化[18]。同时用胁迫系数(stress)衡量MDS二维点阵图结果, 通常认为0.1 调查站位图、种类数和生物多样性指数平面分布图通过软件Surfer11绘制。聚类、标序分析及生物多样性分析均由PRIMER5.0软件包(Plymouth Marine Laboratory , UK )[20-22]完成。本文的甲壳动物种名主要参照《中国海洋生物名录》[23]。 2.1 种类组成和优势种 乐清湾口海域甲壳动物种类组成和优势种以2012年春、秋季2个航次大面定点拖网调查资料为基础, 渔获中甲壳动物种类数从表1可看出, 渔获中共采集甲壳动物34种, 隶属于2目(十足目与口足目)17科23属。其中, 口足目仅口虾蛄()一种, 其余33种均为十足目。虾类18种, 隶属于7科11属, 占本次调查所获甲壳动物种数的52.94%; 蟹类15种, 隶属于9科11属, 占本次调查所获甲壳动物种数的44.11%.十足目中, 长臂虾科和梭子蟹科的种数较多, 分别为6种和7种, 分别占蟹类和虾类种类数的33.33%和46.67%。 从季节变化来看, 春季甲壳动物共21种, 其中虾类12种, 隶属于5科7属, 蟹类9种, 隶属于5科8属; 秋季甲壳动物共24种, 其中虾类15种, 隶属于7科11属, 蟹类9种, 隶属于4科6属。 春季的优势种只有三疣梭子蟹()1种, 其质量占春季甲壳动物总质量的84.73%, 尾数占总尾数的38.98%, 优势度相当高, 为7728.8;秋季甲壳动物优势种有4种, 依次为日本()、口虾蛄、中华管鞭虾()、三疣梭子蟹, 它们的质量占甲壳动物总质量的69.13%, 尾数占总尾数的84.66%(表1)。 表1 乐清湾口海域甲壳动物种类组成及优势种季节变化 注: “”是该种占总质量的百分比, “”是该种占总尾数的百分比, 下同 2.2 种类数的平面分布 乐清湾口海域春秋季甲壳动物种类平均每个站位出现甲壳动物种数分别为7种和6种(图2)。春季位于调查水域中心区域的5号站位物种数最多, 为13种; 乐清湾口最西侧的11号站位最少, 为3种。秋季同样是调查水域中心区域的5号站位物种数最多, 为11种; 小门岛西侧的4号站位未捕获到甲壳动物物种数为0, 乐清湾北侧春秋季2号站位附近种类数均为8种。总体来看, 乐清湾口海域春、秋季甲壳动物种数春秋季变化较为稳定, 空间分布也较为类似。 2.3 生物多样性 由表2可知, 春季甲壳动物Margalef质量丰富度指数()平均值为1.01, 5号站位最高, 为2.09, 8号站位最低, 为0.26; 秋季质量平均值为0.94, 11号站位最高, 为2.20, 4号站位最低, 为0.00。春季尾数平均值为1.56, 5号站位最高, 为2.78, 8号站位最低, 为0.56; 秋季尾数平均值为1.49, 11号站位最高, 为2.87, 4号站位最低, 为0.00。 春季甲壳动物Shannon-Wiener质量生物多样性指数()平均值为1.11, 16号站位最高, 为2.16, 4号站位最低, 为0.19; 秋季质量平均值为1.43, 3号站位最高, 为2.43, 4号站位最低, 为0.00。春季尾数平均值为1.95, 5号站位最高, 为3.02, 9号站位最低, 为1.00; 秋季尾数平均值为1.91, 11号站位最高, 为2.70, 4号站位最低, 为0.00(表2和图3)。 表2 乐清湾口海域甲壳动物群落生物多样性指数的季节变化 春季甲壳动物Pielou质量均匀度指数(′)平均值为0.43, 8号站位最高, 为0.77, 4号站位最低, 均匀度指数为0.07; 秋季质量平均值为0.52, 3号站位最高, 为0.81, 4号站位最低, 为0.00。春季尾数平均值为0.76, 8号站位最高, 为1.00, 2号站位最低, 均匀度指数为0.34; 秋季尾数平均值为0.73, 4号站位最高, 为1.00, 4号站位最低, 为0.00(表2)。 2.4 聚类、排序结果分析 根据乐清湾口海域2个月份调查站位对甲壳动物的群落属性作聚类分析, 横轴代表聚类距离, 纵轴代表调查站位。通常情况下, MDS分析的结果与等级聚类分析的结果可以相互对照, 这样得出的分析结论更具有实际生态学意义[24](图4)。调查海域16个站位, 春季以A1~A16标注, 秋季以B1~B16标注。聚类的结果能较好地影射出调查海域甲壳动物群落结构。每个站位甲壳动物种群群落结构越相似, 其相聚的距离越近。从图4可看出, 在30%相似水平上可将各调查站位分为2个组群, 甲壳动物组成呈明显的季节变化。A1~A12聚为A组, 表明春季这16个站位内部生态属性组成较为相似。在B1~B12聚为B组, 表明秋季16个站位甲壳动物的生态属性组成较相似。两季NMDS排序的胁强系数为0.17, 为0.1~0.2, MDS二维点阵图结果是可以接受的。聚类和NMDS排序的结果都显示了乐清湾口海域甲壳动物群落季节变化趋势和区系平面分布的差异。 a、c.质量(′); b、d.尾数(′) a, c. weight (′); b, d. number (′) 3.1 水文环境特征对甲壳动物生态适宜性影响 乐清湾地属亚热带季风气候区, 受海洋性气候影四季分明, 温暖湿润, 雨量充沛。水体温度年变化较大, 乐清湾可分为雨季和干季[25], 雨季为3~5月份的春雨期和6~7月份的梅雨期及9月份的秋雨期, 其降水量分别占全年的29%~34%和18%及12%~ 13%; 干季为7月中旬至8月和10月至次年2月, 降水量分别占全年的14%~18%和18%及21%~23%。温度和降水变化均较大, 受降水影响, 乐清湾地处近岸, 加上瓯江径流影响, 盐度较低, 所以该海域春秋季甲壳动物共有种以广温低盐性种类为主, 主要种类有: 脊尾白虾()、日本鼓虾()、细螯虾()、鲜明鼓虾()、中国毛虾()、锯缘青蟹(); 受高温高盐的台湾暖流与瓯江径流、浙江沿岸流复杂水系影响, 广温广盐种类也较多, 主要种类有: 葛氏长臂虾()、哈氏仿对虾()、细巧仿对虾()、细指长臂虾()、疣背宽额虾()、中华管鞭虾、周氏新对虾()、红星梭子蟹()、日本、三疣梭子蟹; 该水域也有高温高盐性蟹类光掌()出现, 位于偏外海域6号站位, 该站位处于大门岛和鹿西岛之间, 这可能是秋季受北上台湾暖流的狭管效应, 具有较高的水温和盐度。 从群落区系特征来看, 春季乐清湾口海域甲壳动物主要以优势度较高为暖温型的日本鼓虾、鲜明鼓虾、脊尾白虾等为主, 秋季以暖水型的哈氏仿对虾、中华管鞭虾、锈斑()为主。2个季节甲壳动物群落结构差异较大, 这与NMDS结果一致, 说明该海域甲壳动物群落结构随季节不同而有较明显的变化。这与春秋季海域环境变化有关, 随着季节交替, 秋季水文环境发生改变, 暖水种数量较多, 秋季该海域更趋向于东海以南的亚热带海水温度区系, 而春季温度较低, 更适宜暖温种生长, 而暖水种数量较少, 其优势显著降低, 有的种不在出现(表1)。乐清湾口海域种类组成和分布特征与其生态类型、区系特征和水文环境变化有密切联系。 晁文春等[8]乐清湾南部相近海域南麂列岛海域进行过甲壳动物调查调查, 结果表明, 春季主要优势种为长角赤虾、鲜明鼓虾、口虾蛄、葛氏长臂虾, 夏季主要优势种有口虾蛄、中华管鞭虾、三疣梭子蟹、哈氏仿对虾。与本次调查相比, 结果相差较大, 这可能与调查时间、站位、地理位置等有关有关, 晁文春等调查布设10个站位于春、夏两季调查, 且南麂列岛地理位置特殊, 距大陆较远, 受陆地径流等影响较小, 而本次调查共布设16个站位于春、秋两季调查, 调查站位靠近大陆, 且大多站位位于乐清湾内。结合本次乐清湾调查结果, 总体来看, 口虾蛄、中华管鞭虾、三疣梭子蟹是浙江南部海域重要优势种, 对于三疣梭子蟹、中华管鞭虾这类重要经济种, 应加强保护和合理利用好资源。 3.2 种类数变化及其平面分布特征 春季乐清湾口海域甲壳动物种类数与秋季相比较少。 4月份较10份降水较多[25], 使得4月瓯江径流大于9月, 从图2春季种类数分布中可看出, 春季在瓯江口附近水域形成一舌区, 大量的径流在该区域形成一巨大冲淡水水团, 使得甲壳动物从舌区向外种类数依次增多。10月, 瓯江径流量较少, 台湾暖流北上到达乐清湾口附近, 使甲壳动物种类增多, 空间分布特征复杂, 此外, 由于江浙沿水流与台湾暖流在中部区域相交汇, 带来大量营养物质, 饵料丰富, 物种纷纷到此进行索饵, 种类数在此空间分布较多。在乐清湾北部湾顶、玉环岛西侧区域, 种类数较多, 变化较为稳定, 这是由于湾顶水流与外界接触少, 受影响较小, 加上该水域水深大于10 m的也都位于此[25], 是繁殖、索饵、生长的良好栖息场所, 种类较丰富。 3.3 主要优势种的季节变化特征 三疣梭子蟹为春秋季共同优势种, 春季三疣梭子蟹为单一绝对优势种, 其他优势种优势度不明显, 三疣梭子蟹属广温广盐性经济种, 适应能力较强, 据宋海棠等[26]研究, 4~5月份三疣梭子蟹于浙江中南部沿岸海域产卵, 而乐清湾口海域正位于此处, 数量较多的站位为2、4、10和14号站位, 这些站位质量和尾数合占总数百分比分别为83.30%和78.55%, 从三疣梭子蟹雌雄比来看, 雌蟹均占50%以上, 抱卵性腺成熟度也较高, 此外, 春季其他甲壳动物大都为产卵季节, 个体较小, 但尾数较多, 因此, 三疣梭子蟹尾数占总尾数相对较低, 到了秋季北方冷空气南下, 水温降低, 三疣梭子蟹向外侧和南部较深水域越冬, 数量减少, 所以虽然秋季三疣梭子蟹仍为优势种, 但优势度已大幅降低, 接近常见种的划分范围。此外, 有研究表明[26-27], 盐度对梭子蟹的影响较大, 秋季乐清湾口海域受高温高盐的台湾暖流影响较大, 可能导致秋季三疣梭子蟹较少。 口虾蛄为秋季第二优势种, 为亚热带地区沿海近岸性常见种, 在本次调查春秋季出现率较高, 其繁殖期为4~9月, 高峰期在5~7月, 10月份各种水系相交汇, 饵料丰富, 育肥成长, 质量和尾数较高。口虾蛄和日本类似, 喜栖于浅水泥沙或礁石裂缝内, 所以在15号质量和尾数较高, 分别占总数百分比46.88%和84.40%。 中华管鞭虾为秋季第三优势种, 中华管鞭虾为暖水性热带近岸经济种, 春季水温较低, 不适合中华管鞭虾等暖水种生长, 这可能是春季未捕获的原因。秋季3号站位质量和尾数较高, 分别占总数百分比38.14%和57.93%, 这可能与中华管鞭虾垂直分布特性有关, 在东海中华管鞭虾主要分布在20~60m水深海域[28], 而3号站位位于玉环岛西侧, 为该海域水深较深处[25], 所以3号站位的质量和尾数较高。 3.4 生物多样性变化特征 从甲壳动物生物多样性季节变化来看, 秋季无论质量生物多样性指数还是尾数生物多样性指数均高于春季。丰富度指数与瓯江径流量有一定关系, 春季受瓯江大量径流和降水影响, 乐清湾水域形成淡水团, 物种数较低; 秋季径流量减少, 瓯江径流与外海高温高盐台湾暖流交汇, 物种数增多, 丰富度指数较高。秋季大部分站位值均高于春季, 群落中生物种类增多代表了群落的复杂程度增高, 即值越大, 群落所含的信息量越大[14]。秋季乐清湾口海域甲壳的种类数和优势种的种类数均高于春季。比如, 春季三疣梭子蟹质量远远高于其他种类, 形成单一优势种格局, 而秋季优势种有4种, 当个体数量在种间分配较为均匀, 生物多样性指数′就较高, 这是秋季′值普遍高于秋季的主要原因。春秋季′相差不大, 说明乐清湾口海域甲壳群落结构比较稳定。 从甲壳动物生物多样性区域分布变化来看, 乐清湾口海域甲壳动物种数和生物多样性指数平面分布有一定的特征。从图2和图3平面分布图可看出, 分布规律相似程度较高, 均为调查海域中部和湾顶区域较高。因此可以认为, 调查海域甲壳动物种类与生物多样性指数平面分布特征与地形、水深等外部因素有密切联系。有研究也表明, 群落生物多样性指数的大小同时取决于丰富度指数及均匀度指数, 丰富度和均匀度越高, 生物多样性指数就越高[29]。 此外, 生物之间的捕食关系对乐清湾海域甲壳动物生物多样性也有一定的影响, 甲壳动物处在海洋生物食物链的较低层, 绝大多数甲壳动物是许多海洋经济动物的重要食饵, 其数量变动与高营养级的鱼类等生物存在一定的联系[30]。据闫丽娜等[31]研究, 此次调查春季鱼类优势种为刀鲚和龙头鱼, 秋季优势种仅为龙头鱼, 而刀鲚主要摄食鲜明鼓虾、安氏白虾、脊尾白虾、葛氏长臂虾、口虾蛄等[32], 在一定程度上导致春季甲壳动物种类和数量减少, 生物多样性降低, 影响乐清湾海域群落结构。 3.5 甲壳动物群落结构空间格局 乐清湾口海域甲壳动物群落结构的空间分异明显。根据等级聚类和多维度(NMDS)方法对乐清湾口海域春秋季甲壳动物资源调查资料分析的结果, 乐清湾口海域在30%相似水平上可分为2个组群, 这2个与春秋季两个季节基本对应, 这说明了甲壳动物群落结构与海洋环境密切相关[33]。在40%的相似性水平上, 等级聚类分析的结果形成5组: 第一组为A1、A2、A4~A6、A9~A11、A14~A16, 第二组为A3、A7、A8、A12、A13, 第三组为B6、B13, 第四组为B11, 第五组为B1~B3、B5、B7~B10、B12、B14~B16, 第一组和第二组为春季调查站位, 其余组为秋季站位。该海域春秋季组内并无明显规律, 可能与乐清湾口海域地形、水深的等复杂条件有关。乐清湾内散落着100多个岛屿和众多海湾, 面积大于1000m2的岛礁约30个, 湾内西北向东南倾斜, 由浅滩向深水槽, 中部地形起伏较大, 湾底潮汐汊道与舌状、连岛坝状滩地相间, 乐清湾是我国强潮海湾之一潮差、潮位变幅也较大[25], 加上浙江沿岸流及台湾暖流影响, 导致该海域甲壳动物群落结构复杂。 [1] 黄勃, 堵南山, 赖伟, 等. 河口区甲壳动物对温度、盐度及溶解氧适应能力的研究进展[J]. 海洋科学, 2000, 25(1): 21-23. Huang Bo, Du Nanshan, Lai Wei, et al. Hekou District animal carapace on temperature, salinity and dissolved oxygen ability to adapt to the progress of[J]. Marine Sciences, 2000, 25(1): 21-23. [2] 俞存根, 陈小庆, 宋海棠, 等. 春季东海海域虾类群落结构及其多样性[J]. 生态学报, 2009, 29(7): 3594- 3604. Yu Cungen, Chen Xiaoqing, Song Haitang, et al. In the East China Sea in spring shrimp community structure and diversity of[J]. Acta ecologica Sinica, 2009, 29 (7): 3594-3604. [3] 陈小庆, 俞存根, 宋海棠, 等. 东海中北部海域虾类群聚结构特征及空间分布[J]. 海洋学研究, 2010, 28(4): 50-58. Chen Xiaoqing, Yu Cungen, Song Haitang, et al. In the mid north sea shrimp population structure and spatial distribution of[J]. oceanographic research, 2010, 28(4): 50-58. [4] 陈小庆, 俞存根, 虞聪达, 等. 东海中南部外海虾类组成特征分析[J]. 浙江海洋学院学报, 2010, 29(4): 318-323. Chen Xiaoqing, Yu Cungen, Yu Congda, et al. South East China Sea shrimp characteristics analysis[J]. Journal of Zhejiang Ocean University, 2010, 29(4): 318- 323. [5] 何贤保, 俞存根, 覃涛, 等. 舟山渔场及附近海域虾类群落结构特征分析[J]. 水生生物学报, 2013, 37(3): 553-559. He Xianbao, Yu Cungen, Qin Tao, et al. Structural characteristics of sea shrimp community in Zhoushan fishing ground and near analysis[J]. Acta hydrobiologica Sinica, 2013, 37(3): 553-559. [6] 张洪亮, 王忠明, 祝增军, 等. 浙江南部沿岸产卵场春季虾类群落结构特征分析[J]. 水生生物学报, 2013, 37(4): 712-721. Zhang Hongliang, Wang Zhongming, Zhu Zengjun et al. I wish to, Zhejiang along the southern coast of the spring spawning shrimp community structure characteristics analysis[J]. Acta hydrobiologica Sinica, 2013, 37(4): 712-721. [7] 卢占晖, 薛利建, 张龙, 等. 东海大陆架虾类资源量评估[J]. 水生生物学报, 2013, 37(5): 855-862. Lu Zhanhui, Xue Lijian, Zhang Long, et al. Continental shelf of the East China Sea shrimp resource assessment[J]. Acta hydrobiologica Sinica, 2013, 37(5): 855-862. [8] 晁文春, 何贤保, 苗振清, 等. 春夏季南麂列岛海域甲壳动物种类组成及分布特征[J]. 浙江海洋学院学报, 2013, 32(3): 214-221. Chao Wenchun, He Xianbao, Miao Zhenqing, et al. The composition and distribution characteristics of[J]. Journal of Zhejiang Ocean University, spring and summer in Nanji Islands sea area crustacean animal species 2013, 32(3): 214-221. [9] 赵蒙蒙, 徐兆礼. 三门湾海域冬夏季日足目和十足目虾类的种类组成、时空分布及多样性分析[J], 动物学杂志, 2011, 46(3): 11-18. Zhao Mengmeng, Xu Zhaoli. The species composition, the Sanmen Bay in summer and winter on Isopoda and Decapoda shrimps of the temporal and spatial distribution and diversity analysis of [J], the Journal of animal, 2011, 46(3): 11-18. [10] 徐兆礼, 沈盎绿, 李新正. 瓯江口海域夏、秋季口足目和十足目虾类分布特征[J]. 中国水产科学, 2009, 16(1): 104-112. Xu Zhaoli, Shen anglu, Li Xinzheng. In the Oujiang Estuary in summer and autumn Stomatopoda and full[J]. Chinese Fisheries Science, the distribution characteristics of shrimp head 2009, 16 (1): 104-112. [11] 柏育材, 徐兆礼. 舟山渔场街山岛海域春夏季十足目和口足目的分布特征[J]. 上海海洋大学学报, 2011, 20(1): 96-101. Bai Yucai, Xu Zhaoli. The island area of Zhoushan fisheries Street in spring and summer Decapoda and foot and mouth to distribution[J]. Journal of Shanghai Ocean University, 2011, 20(1): 96-101. [12] 徐开达, 张洪亮, 谢汉阳, 等. 中街山列岛水域甲壳动物资源及其群落多样性[J]. 海洋渔业. 2012, 34(3): 308-315. Xu Kaida, Zhang Hongliang, Xie Hanyang, et al. Crustacean animal resources and community street archipelago waters diversity[J]. marine fisheries, 2012, 34(3): 308-315. [13] 齐海明, 徐兆礼. 椒江口海域春秋季虾类资源分布与环境的关系[J]. 生态学杂志, 2013, 32(8): 2062-2069. Qi Haiming, Xu Zhaoli. Relationship between[J]. Journal of ecology, Jiaojiang port distribution of waters in spring and autumn shrimp resources and environment, 2013, 32(8): 2062-2069. [14] 齐海明, 孙岳, 徐兆礼, 等. 椒江口海域春秋季虾类群落结构及其影响因素[J]. 应用生态学报, 2013, 24(12): 3546-3552. Qi Haiming, Sun Yue, Xu Zhaoli, et al. Jiaojiang estuary in spring and autumn, the community structure and influence factors of shrimp[J]. Chinese Journal of Applied Ecology, 2013, 24 (12): 3546-3552. [15] 齐海明. 椒江口海域甲壳动物群落结构和资源分布及其与环境因子的关系[D].上海: 上海海洋大学, 2014: 1-54. Qi Haiming. Community Structure and Resource Distribution and its Relationship with Environmental Factors in the Sea Area of Jiaojiang Estuary[D]. Shanghai: Shanghai Ocean University, 2014: 1-54. [16] 中华人民共和国农业部. 海洋渔业资源调查规范第5部分: 底拖网调查[M]. 北京: 中国农业出版社, 2012: 2-8. People’s Republic of China Ministry of Agriculture. Fifth parts of marine fishery resources survey: bottom[M]. Survey Beijing: China Agriculture Press, 2012: 2-8. [17] 俞存根, 宋海棠, 姚光展. 东海蟹类群落结构特征的研究[J]. 海洋与湖沼, 2005, 36(3): 213-220. Yu Cungen, Song Haitang, Yao Guangzhan. East China Sea crab community structure research[J]. oceanologia et limnologia Sinica, 2005, 36(3): 213-220. [18] 章飞军, 童春富, 张衡, 等. 长江口潮下带春季大型底栖动物的群落结构[J].动物学研究, 2007, 28(1): 47- 52. Zhang Feijun, Tong Chunfu, Zhang Heng , et al. Study on community structure[J]. animal with spring benthic animal in Yangtze Estuary Tidal 2007, 28(1): 47-52. [19] Khalaf M A, Kochzius M. Change in trophic community structure of shore fishes at an idustrial site in the Gulf of Aqaba, Red Sea[J]., 2002, (239): 287-299. [20] Brazner J C, Beak E W. Patterns in fish assemblages from coastal wetland and beach habitats in Green Bay, Lake Michigan: A multivariate analysis of abiotic and biotic forcing factors[J]., 1997, 54: 1743-1761. [21] Clarke K R. Non-parametric multivariate analysis of changes in community structures[J]., 1993, 18: 117-143. [22] Clarke KR, Warwick R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation[M]. Plymouth, UK: Primers-E Ltd, 1994. [23] 刘瑞玉. 中国海洋生物名录[M]. 北京: 科学出版社, 2008: 1-1267. Liu Ruiyu. Chinese Journal of Marine Biology[M]. Beijing: Science Press, 2008: 1-1267. [24] 厉红梅, 蔡立哲, 林丽珠, 等, 深圳湾潮间带底栖动物群落结构的等级聚类与非度量多维标度排序[J]. 厦门大学学报·自然科学版, 2001, 40(3): 735-740. Li Hongmei, Cai Lizhe, Lin Lizhu, et al. Hierarchical clustering and non metric multidimensional scaling of the benthic community structure of the intertidal zone of Shenzhen Bay[J]. Journal of Xiamen University, Natural Science Edition, 2001, 40 (3): 735-740. [25] 中国海湾志编纂委员会, 中国海湾志: 第六分册[M], 北京: 海洋出版社, 1993. Compilation Committee of bays China, Chinese bays: sixth: [M], Beijing: Ocean Press, 1993. [26] 宋海棠, 俞存根, 薛利建. 东海经济虾蟹类渔业生物学[M]. 北京: 海洋出版社, 2012: 60-67. Song Haitang, Yu Cungen, Xue Lijian. The East China Sea Economic Crab Fishery Biology[M]. Beijing: Ocean Press, 2012: 60-67. [27] 高焕, 张本厚, 陈百尧, 等. 逆境环境因素对三疣梭子蟹幼蟹致死效应分析[J]. 海洋科学, 2010, (2): 57-61. Gao Huan, Zhang Benhou, Chen Baiyao, et al. Effects of Environmental Factors Stress Analysis of [J]. Marine Sciences, three crab crab death 2010 (2): 57-61. [28] 俞存根, 宋海棠, 薛利建, 等. 东海经济虾蟹类[M]. 北京: 海洋出版社, 2006: 19-39. Yu Cungen, Song Haitang, Xue Lijian, et al. East China Economic Crustacean[M]. Beijing: Ocean Press, 2006: 19-39. [29] 程济生, 郭学武. 渤海底栖生物的种类、数量分布及其动态变化[J]. 海洋水产研究, 1998, 19(1): 31- 42. Cheng Jisheng, Guo Xuewu. The species and quantity distribution of benthic organisms and their dynamic changes in Bohai[J]. marine fisheries research, 1998, 19 (1): 31- 42 [30] 逄志伟, 徐宾铎, 陈学刚, 等.胶州湾中部海域虾类群落结构及其多样性[J]中国水产科学, 2013, 20(2): 361-371. Pang Zhiwei, Xu Binduo, Chen Xuegang, et al. In the central Jiaozhou Bay shrimp community structure and diversity of[J] China Fisheries Science, 2013, 20(2): 361-371. [31] 闫丽娜, 叶深, 李德伟, 等. 乐清湾口海域春、秋季鱼类种类组成和数量分布[J]. 海洋与湖沼, 2013, 44(4): 1062-1067. Yan Lina, Ye Shen, Li Dewei, et al. Deep leaf, Yueqing Bay Estuary in spring and autumn fish species composition and quantity distribution[J]. Oceanologia et Limnologia Sinica, 2013, 44 (4): 1062-1067. [32] 俞存根, 陈小庆, 郭远明, 等. 舟山渔场渔业生态学[M].北京: 科学出版社, 2011: 215-223. Yu Cungen, Chen Xiaoqing, Guo Yuanming, et al. Fishery Ecology of Zhoushan Fishing Ground[M]. Beijing: Science Press, 2011: 215-223. [33] Clarke K R, Warwick R. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation[M]. Plymouth, UK: Natural Environmental Research Council, 1994: 144. The study of community structure of crustaceans in Yueqing Bay during spring and autumn XIA Lu-jun1, 2, ZHOU Qing-song3, YU Cun-gen1, SONG Wei-hua3, ZHENG Ji1, BI Si-yao1, XIE Xu1, LI De-wei1, YE Shen1 (1. Zhejiang Ocean University, Zhoushan 316024, China; 2. Bureau of Marine and Fisheries of Shengsi County, Shengsi 202450, China; 3. Second Institute of Oceanography, State Oceanic Administration, Hangzhou 310012, China) Crustacean data based on comprehensive fishery resource surveys were gathered in spring and autumn of 2012 in the mouth of Yueqing Bay, and the species composition, dominance, spatial distribution of species, biological diversity, and similarities of shrimp communities were studied as well as the influential factors in the mouth of Yueqing Bay. The result showed a total of 34 species, which belonged to 23 genera in 17 families of 2 orders. The only dominant species wasin spring, and the dominant species were,andin autumn. The seasonal variation in the dominant species’ seasonal succession phenomenon was very obvious in spring and autumn: the number of species in autumn was higher than in spring; moreover, the species diversity in the center zone was higher than in other places in the survey sea. The biodiversity indices in spring were lower than in autumn, and the community structure of crustaceans had a good relation with the marine environment. Thecrustaceans species types mainly belonged to seasonal, eurythermic, and oligohalabous in the mouth of Yueqing Bay, because this area is adjacent to the Oujiang estuary, which has much running water. Crustaceans have fewer species because of the freshwater coming from Oujiang in spring; on the contrary, the species diversity is larger when the water masses flow together in autumn. External factors, such as terrain, water depth, and the water system have a greater influence on the community structure. crustaceans; biological diversity; community structure; Yueqing Bay mouth (本文编辑: 梁德海) Jan. 26, 2016 [Project of National Natural Science Foundation of China, No.31270527]; enterprise entrusted project, No.2012.03-2013.03] Q178. 1 A 1000-3096(2016)10-0033-10 10.11759/hykx20150909001 2016-01-26; 2016-04-01 国家自然科学基金项目(31270527); 企业委托项目(2012.03- 2013.03) 夏陆军(1991-), 男, 硕士研究生, 研究方向: 渔业资源, E-mail: 670496735@qq.com; 俞存根, 通信作者, 男, 教授, E-mail: cgyu@zjou.edu.cn2 结果
3 讨论
