若尔盖高寒草甸粪堆大小对牦牛粪分解速率的影响
2017-01-11何凯阳小成
何凯,阳小成
(成都理工大学材料与化学化工学院,成都610059)
□研究论文
若尔盖高寒草甸粪堆大小对牦牛粪分解速率的影响
何凯,阳小成*
(成都理工大学材料与化学化工学院,成都610059)
放牧牦牛是若尔盖高寒草甸最重要的草地利用方式,放牧牦牛过程中会产生大量的大小(重量)差异明显的牛粪堆,牛粪的及时分解与否对草地生态系统的养分循环有重要影响。为了探明粪堆大小对牛粪分解速率的影响,调查了牛粪堆大小的野外分布格局,同时,采用人为模拟粪堆大小的实验方法,设置8种不同鲜重(50、100、250、500、700、1 000、1 400和2 000 g)的牛粪堆处理,检测了牛粪的主要分解者功能群(粪甲虫)数量并测定了牛粪分解速率。方差分析结果表明,粪堆大小对牛粪分解速率和甲虫密度均有显著影响。回归分析结果显示,牛粪分解速率与粪堆大小之间呈现曲线(二次方程)关系,即中等大小(1 000~1 400 g)粪堆的分解速率最大;粪堆大小与粪甲虫密度(单位重量牛粪中的甲虫数量)之间存在相似的二次方程关系;粪甲虫密度与牛粪分解速率之间呈现线性关系。野外调查结果显示,鲜重在600~1 500 g的粪堆数量占总数的近50%。以上结果表明,粪堆大小与牛粪分解速率之间的非线性关系可能是牛粪分解者(粪甲虫)对粪堆大小长期适应的结果。
粪堆大小;牛粪分解速率;粪甲虫;中等尺寸效应;高寒草甸
草食动物排泄物(粪便)中含有大量有机质,是放牧生态系统植物生长的重要养分来源[1-4]。在高海拔高寒放牧生态系统,粪便的分解速率通常较低,有效养分回归到土壤的周期较长[5],同时,粪便堆也会直接覆盖植物[6],在一定程度上降低了这些地区的植物生产力。
粪便的分解受降雨、降雪、霜冻和温度等非生物环境条件[7-10]以及土壤动物分解者和微生物等生物因素[1、4、11-13]的影响。研究还表明,粪便的固有属性如大小(重量)、质量、含水量、形状以及产生时间等也能直接或间接地改变这些生物和非生物过程[8、14-15],进而影响粪便的分解速率。草食动物个体之间差异较大,即使是同一个体(如排便期间行走)产生的粪便堆大小也可能不同[8]。粪便堆的大小差异对其内部的理化性质会产生重要影响[14]。例如,大的粪便堆维持较高含水量的时间通常较长[16],同时产生大量异养气体[17]。这些理化性质差异可能进一步影响粪便堆内生物分解者的群落结构[18]。因此,在给定的气候条件下,粪便产生时的大小可能是影响牛粪分解速率的关键属性。
粪甲虫(Scarabaeinae)是多数放牧生态系统中粪便的主要分解者功能群[1-2、4、11-17]。研究表明,粪甲虫群落中的不同物种的取食策略不同。例如,不同物种取食食物颗粒的大小不同。蜉金龟(Aphodius)只能取食直径小于25 μm的食物颗粒[19],而蜣螂能够取食直径最大为85 μm的颗粒[11]。另外,掘洞型甲虫(Paracoprids)通常把食物(粪球)带入粪堆下面的洞中取食;滚粪球型甲虫(Telecoprids)把粪球搬离粪堆取食;粪居型甲虫(Endocoprids)则直接在粪堆中取食和繁殖[20]。粪甲虫取食策略之间的差异有利于牛粪分解,那么粪甲虫对粪堆大小如何响应呢?我们可以假设,如果粪堆中的甲虫个体大小不受粪堆大小影响,而甲虫密度随粪堆大小成比例改变,那么粪便的分解速率与粪堆大小无关;然而,如果由于长期的适应,粪甲虫偏向于在某些特定大小的粪堆中生活,那么粪便的分解速率和粪堆大小有关。目前,一些研究表明粪甲虫的物种丰富度以及粪便分解速率随粪堆大小的增加而增加[16-17],初步揭示了粪堆大小对粪便分解速率的影响。但是,这些研究所设置的粪堆大小的梯度均较小(例如,30 g和750 g[16];2 mL和200 mL[17];0.25 L、0.5 L和1.0 L[21];0.25 L、0.5 L、1.0 L和1.5 L[22]),不能真实反映自然情况下粪堆大小的分布格局。因此,粪堆大小对粪便分解速率的影响及其机制仍有待进一步研究。
若尔盖高寒草甸主要分布于甘肃省南部和四川省西北部,具有很高的生产力,是我国第五大畜牧生产基地[23]。高寒草甸的放牧系统中,牲畜(主要是牦牛)取食的植物大部分并未被吸收,而是以粪便的形式回归到地表[24]。据统计,不同大小的牦牛粪堆对草地的覆盖率在一些区域最高可达20%[4]。而且,尽管高寒草甸的降水相对充足,但年均温度较低,导致牛粪中的有机质分解缓慢,土壤有效养分较低,制约着生态系统生产力的提高[4]。因此,开展牛粪分解的相关研究十分必要。本文基于牛粪堆大小在自然情况下的分布格局,设置8种(50、100、250、500、700、1 000、1 400和2 000 g)不同大小的牛粪堆处理,通过测定牛粪分解速率、含水量以及主要分解者功能群(粪甲虫)数量等指标,旨在揭示粪堆大小对牛粪分解速率的影响。
1 研究地概况
本研究在若尔盖高寒草甸红原县(32°48′—32°52′N,102°01′—102°33′E)开展。红原县海拔约3 500 m,年均温0.9℃,最热月(7月)月均温10.9℃,最冷月(1月)月均温-10.3℃,年均降水量690 mm左右,年际波动大,其中80%的降水集中在5月至8月。植物群落以藏北嵩草(Kobresia littledalei)、芒剪股颖(Agrostis trinii)、甘青剪股颖(A.hugoniana)。鹅绒委陵菜(Potentilla anserina)、条叶银莲花(Anemone trullifolia)等为优势种[25]。生长季集中在6、7、8月3个月。生长季植被总盖度超过80%,植被平均高度约为30 cm。
放牧牦牛是研究区域草地的主要利用方式。试验区内主要放牧牦牛品种为麦洼牦牛和九龙牦牛,牧民放牧方式主要为冬春、夏秋两季放牧,即将牧场分为冬春放牧的冷季牧场和夏秋放牧的暖季牧场,同时也存在全年可放牧的常年牧场和全年不放牧的禁牧草场[4]。野外调查表明,暖季牧场内牦牛种群密度大约为10只/公顷,牦牛粪的密度最大可达5 900堆/公顷[4]。与其他放牧生态系统类似,若尔盖高寒草甸生态系统中牛粪分解者的物种种类丰富,其中,粪甲虫是最主要的分解者类群[4]。粪甲虫种类主要包括游荡蜉金龟(Aphodius erraticus)、直蜉金龟(Aphodius rectus)、雅蜉金龟(Aphodius coniugatus)、长胫嗡蜣螂(Onthophagus yubarinus)、腐嗡蜣螂(Onthophagus tabidus)等;捕食者主要包括捕食性甲虫[如隐翅虫(Philonthus purpuripennis)和藏甲(Nicrophorus japonicus)]以及蜘蛛(Araneus marmoreus)和蜈蚣(Scolopendra alternans)等;牛粪分解后期,粪堆中以蛴螬型甲虫幼虫为主。另外,一些蝇类包括黄粪蝇(Scathophaga stercoraria Linnaeus)和红头丽蝇(Calliphora vicina)等的幼虫在牛粪分解前期也会出现[4]。
2 研究方法
2.1 实验设计和取样
2009年4月在暖季牧场围封面积为100 m×100 m的样地,样地内的植被盖度超过90%。样地围封后清除样地内的所有牛粪。同年7月10日至15日,在研究区域内的草场(约1公倾)随机调查了100堆新鲜(1-2天)牛粪的鲜重、直径和高度。
野外控制实验于7月19日开始。实验用牛粪为取自牧民家牛棚中的新鲜牛粪(1-2天)。将牛粪混合均匀后,分别称取鲜重50、100、250、500、700、1 000、1 400和2 000 g的牛粪,用自制的圆形金属模型(高4 cm,直径分别为5、8、12、15、18、20、25和30 cm)将牛粪随机地放置在样地内,牛粪堆之间间隔至少5 m。实验设置8种处理,每种处理4个重复,共放置32堆不同大小的牛粪堆。9月8日实验结束时,用镊子仔细拣出牛粪内及其下面土壤中的甲虫(包括成虫和幼虫)并计数,将剩余牛粪收集在塑料自封袋中,带回实验室烘干(75℃)称重。
2.2 数据分析
参考Holter和Hendriksen[26]的方法,牛粪分解速率用相对分解速率(g g-1d-1)表示,计算公式为相对分解速率=ln(M0/Mt)/t(1)
(1)式中:M0(g)为初始牛粪干重;Mt(g)表示第t天时的剩余干重;t(d)为分解时间。甲虫密度为单位牛粪干重中的甲虫数量。由于实验结束取样时,牛粪中只有蛴螬型甲虫幼虫,并没有发现活着的成虫,因此,本文中甲虫密度指的是甲虫幼虫密度。采用单因素方差分析(One-way ANOVA)来分析粪堆大小对牛粪分解速率以及甲虫密度的影响。采用线性或二次方程回归分析牛粪大小、牛粪分解速率和甲虫密度之间的关系。所有数据处理均在STATISTICA分析软件中进行。
3 研究结果
3.1 牛粪堆大小的分布格局
野外调查结果显示(表1),质量介于600~1 500 g的粪堆的数量最多,占总数的48%;100~600 g的粪堆占总数的39%;小于100 g的粪堆的数量较少,占总数的7%;鲜重大于2 000 g的粪堆仅占总数的2%。
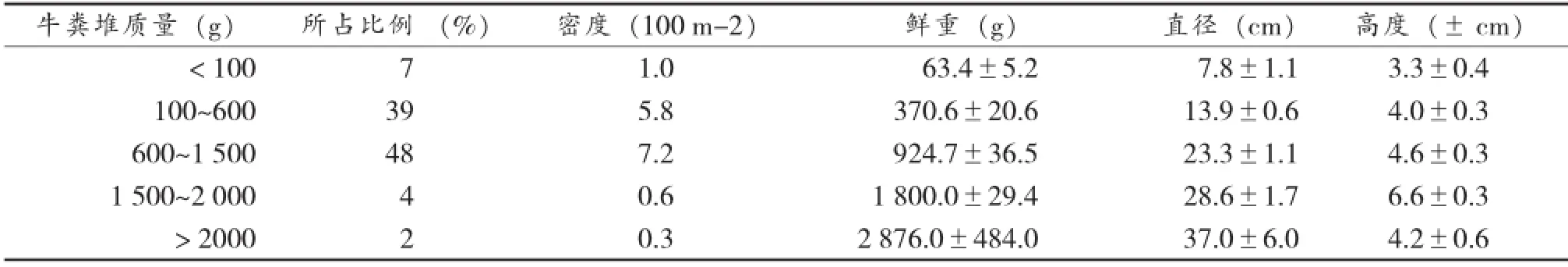
表1 自然条件下牛粪堆大小的分布状况Table 1 Distribution of yak pat size in natural conditions
3.2粪堆大小、分解速率和粪甲虫密度之间的关系
方差分析结果表明,粪堆大小对牛粪分解速率(F7,31=46.45,P<0.001)和粪甲虫密度(F7,31=16.18,P<0.001)均有显著影响。具体而言,牛粪分解速率在粪堆大小为1 400 g时最大,为0.028 g g-1d-1(图1a);粪甲虫密度在粪堆质量为1 000 g时达到最大(图1b),此外,牛粪分解速率与粪甲虫密度之间呈现线性关系(图2)。
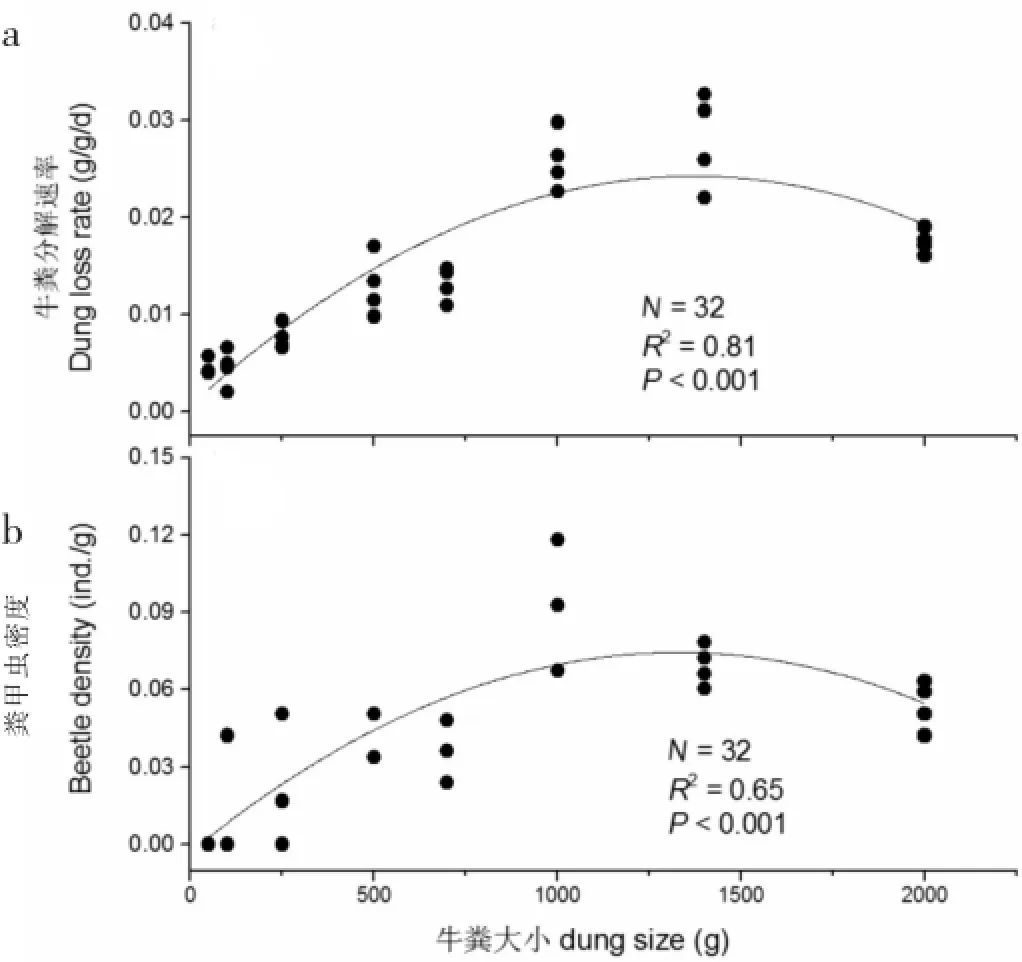
图1 粪堆质量与牛粪分解速率(a),粪堆大小与粪甲虫密度(b)之间的关系Fig.1 The relation between yak pat size and yak pat composition rate(a),beetle density(b)
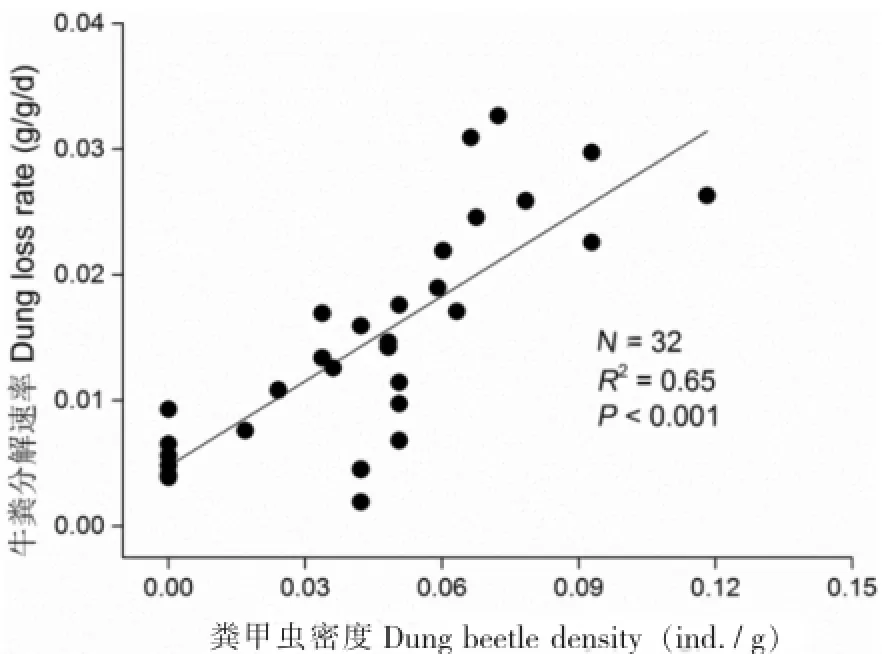
图2 粪甲虫密度与牛粪分解速率之间的关系Fig.2 The relation between dung beetle density with yak pat composition rate
4 讨论
本研究发现,粪堆大小对牛粪分解速率有显著影响,体现在中等大小粪堆的分解速率最大;粪甲虫密度在中等大小粪堆中最大;粪甲虫密度与牛粪分解速率之间呈线性关系。以上结果表明,粪堆大小能够通过影响牛粪中的粪甲虫数量,最终改变牛粪分解速率。
大的粪堆能够吸引更多粪甲虫的主要原因包括以下几个方面。首先,大的粪堆能够散发出更多的气味从而吸引更多的粪甲虫[17]。Errouissi等[16]的研究也表明大粪堆中粪甲虫的物种丰富度和多度均高于小粪堆。其次,大的粪堆能够为甲虫,特别是一些个体较大的物种提供更充足的活动空间[17,22]。例如,对于体型较大的掘洞型甲虫而言,它们掘洞时需要雄性和雌性个体的协作[27],需要更大的空间。再次,牛粪含水量是粪甲虫生存和繁殖的关键因素之一[8]。由于大粪堆的表面积/体积比相对较大,其水分蒸发相对较慢[21],因此大粪堆中的含水量明显较高。高含水量不仅有利于分解者(如粪甲虫)活动[28],而且还能提高分解者对粪便的利用率[29]。更重要的是,大的粪堆能够给粪甲虫提供更多的食物资源。粪甲虫多样化的取食策略使其能够取食大量牛粪[11],因此,充足的食物资源是其生存保障。前期研究表明,粪甲虫具有很高的分解效率(每只成虫每天能够取食0.6 g[4]),(蛴螬型)幼虫每只每天也能取食大约0.015 g牛粪[12]。由此可见,大量粪甲虫只能共存于大的粪堆中。
既然大粪堆具有相对优势,那么是否意味着粪堆越大粪甲虫密度越高呢?事实并非如此。我们发现,粪甲虫数量并没有随着粪堆大小增加而成比例增加,在粪堆大于1 500 g后,粪甲虫的密度反而下降。出现这种情况的原因可能是微生物活动减少了牛粪中的氧气含量,不利于粪甲虫生存。牛粪中尤其是中心部分,氧气浓度较低(小于2%),限制了粪甲虫的活动。同时,牛粪中大量的一氧化二氮、甲烷等气体进一步增加了对粪甲虫的生理胁迫[30-33]。Holter[32]的研究表明,甲烷的产生量与粪堆大小是成正比的,这就意味着随着粪堆大小的增加,粪甲虫需要承受更大的生理胁迫。此外,如果粪堆过大,其过高的含水量也可能会对粪甲虫形成胁迫。粪甲虫的产卵及其幼虫成活率对水分条件有较高要求,例如,蜉金龟科成虫产卵的最适宜湿度是55%~75%[29];当含水量约低于68%时,蜣螂(Euoniticellus intermedius)则不能繁殖[8]。
鉴于粪甲虫密度与牛粪分解速率之间的线性关系,我们不难理解:粪堆大小能够通过改变牛粪中的非生物(如牛粪含水量)和生物(粪甲虫密度)过程,进而影响牛粪的分解速率。需要注意的是,由于本研究实验设计的不完善(如重复数量较少),实验过程中尤其是实验前期无法统计粪甲虫成虫的物种丰富度和多度,在一定程度上限制了我们对实验结果更深入的解释。即使如此,我们认为幼虫密度是可以反映成虫密度的。而且,由于生长发育速度较快,粪甲虫幼虫自身能够取食成虫不能取食的大颗粒和纤维食物,具有较高的取食效率[34]。
综上所述,粪堆大小和牛粪分解速率之间的非线性关系可能是青藏高原气候条件下放牧牦牛和粪甲虫长期协同进化的结果。该推测与野外粪堆大小的分布情况一致,即大多数的粪堆重量在600~1 500 g之间。本结果对于研究青藏高原高寒草甸放牧牦牛与粪甲虫之间的长期协同进化与适应具有一定的参考价值。
[1]MOHR C O.Cattle Droppings as Ecological Units[J].Ecological Monographs,1943,13(3):275-298.
[2]BANG H S,LEE J H,KWON O S,at al.Effects of Paracoprid Dung Beetles(Coleoptera:Scarabaeidae)on the Growth of Pasture Herbage and on the Underlying Soil[J].Applied Soil Ecology,2005,29(2):165-171.
[3]何奕忻,孙庚,罗鹏,等.牲畜粪便对草地生态系统影响的研究进展[J].生态学杂志,2009,28(2):322-328.
[4]WU X,SUN S.Roles of Beetles and Flies in Cattle Dung Removal in an Alpine Meadow of Eastern Qinghai-Tibetan Plateau[J].E-coscience,2010,17(2):146-155.
[5]何奕忻,孙庚,刘琳,等.牦牛粪便对川西北高寒草甸土壤养分的影响[J].应用与环境生物学报,2009,15(5):666-671.
[6]姜世成,周道玮.牛粪堆积对草地影响的研究[J].草业学报,2006,15(4):30-35.
两组患儿诊疗护理2周后观察其临床治疗效果,具体情况(见表1),实验组患儿治愈率、有效率均明显高于对照组。
[7]WHITE E.The Distribution and Subsequent Disappearance of Sheep Dung in the Pennine Moorland[J].Jourhal of Animal Ecology,1960,29(2):243-250.
[8]EDWARDS P B.Seasonal Variation in the Dung of African Grazing Mammals,and Its Consequences for Coprophagous Insects[J]. Functioual Ecology,1991,5(5):617-628.
[9]MENENDEZ R,GUTIERREZ D.Altitudinal Effects on Habitat Selection of Dung Beetles(Scarabaeoidea:Aphodiidae)in the Northern Iberian Peninsula[J].Ecography,1996,19(3):313-317.
[10]吴新卫,李国勇,孙书存.降水量对川西北高寒草甸牦牛粪分解速率的影响[J].生态学报,2011,31(23):7013-7021.
[11]HOLTER P,SCHOLTZ C H.Are Ball-rolling(Scarabaeini,Gymnopleurini,Sisyphini)and Tunnelling Scarabaeine Dung Beetles E-qually Choosy about the Size of Ingested Dung Particles[J].Ecological Entomology,2005,30(6):700-705.
[12]HOLTER P.An Experiment on Dung Removal by Aphodius larvae(Scarabaeidae)and Earthworms[J].Oikos,1977,28(1):130-136.
[13]HOLTER P.Resource Utilization and Local Coexistence in a Guild of Scarabaeid Dung Beetles(Aphodius spp.)[J].Oikos,1982,39(2):213-227.
[15]DDORMONTL,RAPIOR S,MCKEY DB,LUMARET JP.Influence of Dung Volatiles on the Process of Resource Selection by Coprophagous Beetles[J].Chemoecology,2007,17(1):23-30.
[16]ERROUISSI F,HALOTI S,JAY-ROBERT P,et al.Effects of the Attractiveness for Dung Beetles of Dung Pat Origin and Size A-long a Climatic Gradient[J].Envioronmental Entomology,2004,33(1):45-53.
[17]PECK S B,HOWDEN H F.Response of a Dung Beetle Guild to Different Sizes of Dung bait in a Panamanian Rainforest[J].Biotropica,1984,16(3):235-238.
[18]SEVENSTER J G,ALPHEN J V.Aggregation and Coexistence.II.A Neotropical Drosophila Community[J].Jourhal of Animal Ecology,1996,65(3):308-324.
[19]HOLTER P.Particle Feeding in Aphodius Dung Beetles(Scarabaeidae):Old Hypotheses and New Experimental Evidence[J]. Founctional Ecology,2000,14(5):631-637.
[20]DOUBLE B M.A Functional Classification for Analysis of the Structure of Dung Beetle Assemblages[J].Ecological Entomology,1990,15(4):371-383.
[21]GITTINGS T.The Community Ecology of Aphodius Dung Beetles[D].National University of.Ireland:Cork,1994.
[22]FINN J A,GILLER P S.Patch Size and Colonisation Patterns:an Experimental Analysis Using North Temperate Coprophagous Dung Beetles[J].Ecography,2000,23(3):315-327.
[23]XIANG S,GUO R,WU N,et al.Current Status and Future Prospect of Zoige Marsh in Eastern Qinghai-Tibet Plateau[J].Ecological Engineering,2009,35(4):553-562.
[24]CHEW R M.Consumers as Regulators of Ecosystems:an Alternative to Energetics[J].The Ohio Journal of Science,1974,74(6):359-370.
[25]ZHAO C,GRIFFIN J N,WU X,et al.Predatory Beetles Facilitate Plant Growth by Driving Earthworms to Lower Soil;ayers[J]. Jourhal of Animal Ecology,2013,82(4):749-758.
[26]HOLTER P,HENDRIKSEN N B.Respiratory Loss and Bulk Export of Organic Matter from Cattle Dung Pats:a Field Study[J]. Ecography,1988,11(11):81-86.
[27]SOWIG P.Habitat Selection and Offspring Survival Rate in Three Paracoprid Dung Beetles:the Influence of Soil Type and Soil Moisture[J].Ecography,1995,18(2):147-154.
[28]DICKINSON C H,CRAIG G.Effects of Water on the Decomposition and Release of Nutrients from Cow Pats[J].New Phytologist,1990,115(1):139-147.
[29]LUMARET J P,KIRK A.Ecology of Dung Beetles in the French Mediterranean Region(Coleoptera:Scarabaeinae)[J].Acta Zoologica Mexicana,1987(NS),24:1-55.
[30]HOLTER P.Concentration of Oxygen,Carbon Dioxide and Methane in the Air Within Dung Pats[J].Pedobiologia,1991,35(6):381-386.
[31]HOLTER P.Tolerance of Dung Insects to Low Oxygen and High Carbon Dioxide Concentrations[J].European Journal of Soil Biology,1994,30(4):187-193.
[32]HOLTER P.Methane Emissions From Danish Cattle Dung Pats in the Field[J].Soil Biology&Biochemistry,1997,29(1):31-37.
[33]YAMULKI S,JARVIS S C,OWEN P.Nitrousoxide Emission from Excreta Applied in a Simulated Grazing Pattern[J].Soil Biology&Biochemistry,1998,30(4):491-500.
[34]HALFFTER G,MATTHEWS E G.The Natural History of Dung Beetles of the Subfamily Scarabaeinae(Coleoptera:Scarabaeidae)[J]. Folia Entomologica Mexicana,1969,45(2):426-427.
[责任编辑:向晚]
Impacts of Dung Pat Size on the Decomposition Rate of Yak Dung in a Zogie Alpine Meadow
HE Kai,YANG Xiao-cheng*
(College of Materials and Chemistry&Chemical Engineering,Chengdu University of Technology,Chengdu 610059,China)
There is a large variation in dung pat size even from a single yak species in the Zogie Alpine Meadow,a typical yak grazing grassland.Thus,dung decomposition rate is important to the nutrient cycling in this grazing ecosystem.To examine the size effect on decomposition rate of yak dung,we conducted a field experiment i nvolving eight size classes of circular dung pats differing in fresh weight(i.e.50、100、250、500、700、1000、1 400 and 2 000 g),each class having four replicates.At the end of the experiment,we examined dung loss rate and beetle density for each dung pat.The relative dung loss rate(g g-1d-1)showed a curvilinear(quadratic)relationship with dung pat size,with the highest rate being in the middle-sized dung pats(1 000-1 400 g).Similar relationships were observed between beetle density(individuals per dung mass)and dung pat size.Significant positive linear relationships were observed between beetle density and dung loss rate.In addition,an independent field survey showed that about 50%of yak dung pats ranged from 600 g to 1 500 g in fresh mass in the study alpine pasture. These results suggest that the non-linear relationship between dung pat size and dung loss rate might be due to the long-history evolution and adaptation of the within-dung detritivores(dung beetles)to the frequency distribution of dung pat sizes.
dung pat size;dung loss rate;dung beetle;middle-size effect;alpine meadow
S154.3
A
2096-2347(2016)02-0025-06
10.19478/j.cnki.2096-2347.2016.02.04
2016-04-29
国家自然科学基金项目(31000232,31200375)。
何凯(1990—),男,四川绵阳人,硕士研究生,E-mail:hekai007@sina.cn
*[通讯作者]阳小成(1965—),男,重庆长寿人,博士,教授,硕士生导师,主要从事植物生态学研究。E-mail:yxc39@163.com
