地形对七姊妹山自然保护区植物丰富度及分布格局的影响
2017-01-04熊斌梅汪正祥李亭亭熊秀海李中强彭宗林
熊斌梅,雷 耘,汪正祥,3,4*,李亭亭,熊秀海,李中强,3,4,张 娥,彭宗林
(1 湖北大学 资源环境学院,武汉430062;2 华中师范大学 生命科学学院,武汉 430070;3 湖北省生物资源绿色转化协同创新中心,武汉 430062;4 区域开发与环境响应湖北省重点实验室,武汉 430062;5 武汉大学 资源环境学院,武汉430072;6 七姊妹山自然保护区管理局,湖北 宣恩 445500)
地形对七姊妹山自然保护区植物丰富度及分布格局的影响
熊斌梅1,雷 耘2,汪正祥1,3,4*,李亭亭1,熊秀海5,李中强1,3,4,张 娥1,彭宗林6
(1 湖北大学 资源环境学院,武汉430062;2 华中师范大学 生命科学学院,武汉 430070;3 湖北省生物资源绿色转化协同创新中心,武汉 430062;4 区域开发与环境响应湖北省重点实验室,武汉 430062;5 武汉大学 资源环境学院,武汉430072;6 七姊妹山自然保护区管理局,湖北 宣恩 445500)
该研究以七姊妹山自然保护区40个(20×20 m2)植物群落调查样方为基础,并采用回归分析和典型对应分析(CCA)的方法研究该区地形对植物物种丰富度及植物分布格局的影响,以明确海拔、坡度、坡向、坡位等地形因子的相对重要性,为该区植物多样性的保护和管理提供理论依据。结果表明:(1)七姊妹山自然保护区40个调查样地共有植物633种,隶属133科,316属,其中乔木118种,灌木150种,草本365种。(2)曲线回归方程拟合结果显示,七姊妹山自然保护区植物物种丰富度分别与海拔、坡度具有显著相关性,物种丰富度沿海拔梯度升高而增大,沿坡度梯度先减少后增大之后又减小。(3)从植物的生活型来看,在所有海拔段,乔木物种丰富度始终低于灌木和草本植物;在低、中低海拔地带,灌木物种丰富度均高于乔木和草本植物;而在中、高海拔地带草本植物物种丰富度较大且高于乔木和灌木。(4)CCA排序结果表明,地形因子对植物物种的分布具有显著影响按其影响强度排序为海拔>坡度>坡位>坡向,说明海拔是影响该区植物物种分布最重要的地形因子。
地形;物种丰富度;物种分布;典型对应分析;七姊妹山自然保护区
山地植物群落物种多样性及空间分布受地形因子(海拔、坡度、坡向、坡位)的影响较大,一直是生态学研究的热点领域[1-2]。地形因子通过控制太阳辐射和降水的空间分布间接对物种丰富度及分布格局产生显著的影响[3-4],地形因子中海拔梯度包含光照、热量、水分等的梯度变化,对物种丰富度分布格局具有决定性作用[5-6]。坡度影响土壤厚度、土壤水分、养分等从而影响植物的生长,但坡度对物种丰富度的影响并没有统一的结论,相关研究表明,缓坡和陡坡物种丰富度最高[3],也有研究证明中等坡度物种丰富度最高[7],但其他相关研究发现,坡度与物种丰富度无显著相关性[8]。不同的研究区海拔、坡度等地形因子对物种丰富度及分布影响的相对重要性不同,因此对典型区域的研究尤为重要。七姊妹山自然保护区属于典型的亚热带山地,当前研究者们主要对该区植被类型、植物区系、植物资源的开发与利用、保护策略、群落物种多样性特征等方面进行研究[9-13],但对于该区地形对物种丰富度及分布影响的研究尚未见报道。本研究以七姊妹山自然保护区40个调查样方为基础,分析了地形因子对物种丰富度及分布的影响,并确定了海拔、坡度、坡向、坡位等地形因子的相对重要性,为该区植物多样性的保护和管理提供理论依据。
1 研究区概况
七姊妹山自然保护区位于湖北省恩施土家族苗族自治州宣恩县东部(109°38′30″~109°47′00″E,29°39′30″~30°05′15″N),该区属于中亚热带季风湿润型山地气候,气候垂直差异明显。海拔800 m以下的低山带年均温15.8 ℃,无霜期在294 d,年降水量1 491.3 mm,年日照时数1136.2 h;海拔在800~1 200 m的山地,年平均气温13.7 ℃,无霜期263 d,年降水量1 635.3 mm,年日照时数1 213 h;海拔1 200 m以上的高山地带年均温8.9 ℃,无霜期203 d,年降水量1 876 mm,年日照时数1 519.9 h。该地域的岩层主要由石英页岩、砂质层岩组成,土壤主要由黄壤、黄棕壤、棕壤等种类组成[9]。
七姊妹山自然保护区地处中亚热带和北亚热带的过渡地带,是中国三大特有现象中心之一的“川东-鄂西特有现象中心”的核心地带,属于泛北极植物区,中国-日本森林植物亚区,华中地区的北缘,正处于华中植物区系、西南植物区系和华南植物区系的交汇处。区内火烧堡是最高峰(2 014.5 m),最低海拔为650 m。该区由于地理位置独特,垂直高差明显,具有丰富的植物资源,共有维管束植物183科,752属,2 027种,包含温性针叶林、暖性针叶林、常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、灌丛、灌草丛、温性竹林、苔藓沼泽9个植被类型[9]。
2 研究方法
2.1 野外调查
在七姊妹山自然保护区沿海拔梯度选择具有代表性的植物群落设置40个面积为20 m×20 m调查样方,设置的样方尽可能包含不同坡度、坡向、坡位。采用植物社会学方法对乔木和灌木样地进行植被调查,记录乔木层、灌木层及草本层的种名、多度、盖度;同时测量并记录地形因子(海拔、坡度、坡向、坡位)、经纬度、土壤、林分、干扰等情况。
2.2 数据分析
2.2.1 物种丰富度 物种丰富度是按样地面积为400 m2的物种数计算。用SPSS19.0软件对地形因子与物种丰富度进行回归分析。
2.2.2 排序 对七姊妹山自然保护区物种-样地进行除趋势对应分析(detrended correspondence analysis,DCA),DCA排序结果显示,最长梯度值为7.6,故本研究采用典型对应分析(canonical correspondence analysis,CCA)研究地形因子对植物物种分布格局的影响。CCA排序所用的植物数据是物种的多度,组成(样方×物种)40×634的植被数据矩阵;地形因子包括海拔、坡度、坡向、坡位,组成(样方×地形因子)40×4的地形因子数据矩阵。为了便于定量分析及建立矩阵,把各地形指标加以量化:海拔、坡度直接用实测数据;坡位赋值,1代表下坡、2代表中坡、3代表山(顶)脊侧坡、4代表山(顶)脊;坡向赋值1~8分别代表北坡、东北坡、东坡、东南坡、南坡、西南坡、西坡和西北坡[3,14]。
用Canoco for Windows 4.5进行 CCA排序,用CanocoDraw for Windows 4.1完成图。排序图中仅显示物种适合度在10%~100%,权重在5%~100%间的物种,图中数字代表物种。
3 结果与分析
3.1 物种丰富度与地形因子关系
3.1.1 物种组成 在40个调查样地中,共有植物633种,隶属133科,316属(其中包含苔藓植物泥炭藓1科1属1种,其余为维管束植物),其中乔木118种,灌木150种,草本365种。主要由多脉青冈(Cyclobalanopsismultinervis)、耳叶杜鹃(Rhododendronauriculatum)、泥炭藓(Sphagnumpalustre))、珙桐(Davidiainvolucrata)、吉祥草(Reineckeacarnea)、腊莲绣球(Hydrangeastrigosa)、亮叶水青冈(Faguslucida)、杉木(Cunninghamialanceolata)、细叶水芹(Oenanthedielsii)、小叶青冈(Cyclobalanopsismyrsinifolia)、宜昌木姜子(Litseaichangensis)、四照花(Dendrobenthamiajaponicavar.chinensis)、三桠乌药(Linderaobtusiloba)等种类组成。
3.1.2 物种丰富度与海拔、坡度的关系 图1和图2分别显示了物种丰富沿海拔和坡度梯度的变化趋势。物种丰富度与海拔和坡度之间存在显著非线性关系(P<0.05)。图1中物种丰富度随海拔的升高大致呈递增的趋势,在海拔为1 500 m左右达到峰值,这与该地降水随海拔的变化趋势一致。图2中物种丰富度随坡度的变化大致呈现先减小后增大再减小的趋势,即在缓坡和较陡坡物种丰富度较大。
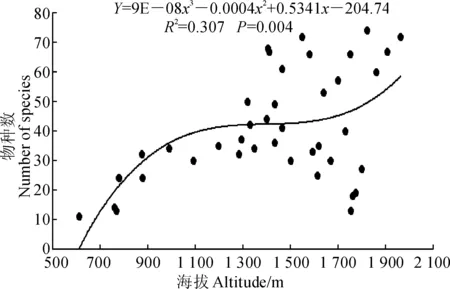
图1 物种丰富度与海拔的关系Fig.1 Relationship between species richness and altitude

图2 物种丰富度与坡度的关系Fig.2 Relationship between species richness and slope

图3 乔木、灌木和草本植物物种丰富度随海拔梯度的变化Fig.3 Species richness of trees, shrubs and herbs vary along the altitudinal gradient and ordination summary

表1 环境因子与CCA前4轴排序的相关系数及排序概要
*P<0.05;**P<0.01
3.2 海拔对不同生活型植物物种丰富度的影响
图3是乔、灌、草3种生活型植物物种丰富度随海拔梯度的变化趋势。由图3可知,乔木物种丰富度随海拔梯度的变化趋于平缓,而灌木和草本变化幅度较大。在各个海拔段,乔木物种丰富度始终低于灌木和草本植物,在1 600 m左右的海拔段乔木物种丰富度达到峰值,随后逐渐降低直至零,乔木物种丰富度整体呈现单峰格局;在低海拔(600~1 300 m)段,灌木物种丰富度高于乔木和草本植物,但当海拔大于1 300 m时,草本植物物种丰富度迅速增大并且高于灌木和乔木;总体来说,随着海拔升高,灌木和草本植物物种丰富度呈增加的趋势,乔木物种丰富度在中海拔段出现峰值后逐渐减少。乔木、灌木和草本3种生活型植物物种丰富度沿海拔梯度变化大致趋势与该区植物总丰富度沿海拔分布格局基本一致。
3.3 地形因子与CCA排序轴的关系
对七姊妹山自然保护区植物物种与地形因子进行CCA排序,经蒙特卡罗检验结果显示,所有典范轴都极显著(P= 0.002<0.01),说明地形因子对七姊妹山自然保护区植物的分布格局具有显著影响。CCA排序4个轴累积解释了物种-环境关系方差的100%,说明CCA排序的4个轴较好地解释了物种-地形因子的关系。CCA排序的4个轴代表不同的地形因子梯度变化,根据地形因子与植物排序轴的相关性的大小(表1),海拔、坡向、坡位与CCA 1轴呈极显著负相关,而与坡度呈极显著正相关,按相关性大小排序为海拔>坡位>坡度>坡向,表明第1轴主要反映海拔、坡度的梯度变化;海拔、坡度与CCA 2轴呈极显著负相关,而与坡向呈极显著正相关,按地形因子与CCA 2轴相关性大小为坡度>海拔>坡向>坡位,说明CCA 2轴主要反映坡度的梯度变化;坡位、坡度与CCA 3轴呈极显著正相关,而与海拔、坡向无显著相关性,第3轴主要反映坡位、坡度的梯度变化;除坡向与CCA 4轴相关性系数达0.70外,其余地形因子均无显著相关性,说明第4轴主要反映坡向的梯度变化。上述情况说明,海拔梯度是影响七姊妹山自然保护区植物分布格局最重要的地形因子,其次为坡度、坡位、坡向。
3.4 地形因子对物种分布格局的影响
CCA排序图把植物与地形因子相结合,能综合反映群落物种分布与地形因子之间的关系。图4中箭头代表地形因子,箭头连线的长度代表植物群落的分布与该环境因子相关性大小,连线越长表示对你植物的分布影响越大;箭头连线与排序轴的斜率表示地形因子与排序轴相关性大小[15]。

Elve. 海拔;Slop. 坡度;Aspe. 坡向;Posi. 坡位;21.薄叶山矾;31.藏刺榛;37.常春藤;58.粗边扁蕾;65.大理苔草;74.大羽贯众;80.淡红忍冬;83.灯笼花;86.点叶落地梅;98.椴树;101.多齿长尾槭;105.鹅掌柴;110.鄂西绣线菊;111.耳叶杜鹃;113.繁缕;129.革叶耳蕨;132.珙桐;133.钩栲;140.贯众;166.红毛七;170.湖北海棠;185.华西花楸;194.黄连木;197.黄水枝;209.戟叶蓼;222.箭竹;238.荩草;247.蕨;261.腊莲绣球;270.棱果海桐;277.亮鞘苔草;288.六叶葎;296.鹿蹄草;299.落新妇;301.绿叶甘橿;303.马尾松;304.马银花; 307.满山红;313.芒萁;315.猫儿屎;322.茅栗;338.泥炭藓;346.泡叶栒子;358.荞麦;363.青冈;377.日本苔草;389.三桠乌药;401.山合欢;402.山胡椒;403.山鸡椒;414.杉木;424.肾唇虾脊兰;428.石松;440.水青树;442.四川冬青;448.穗序鹅掌柴;475.卫矛;497.细叶水芹;518.小灯芯草;519.小果南烛;524.小叶石楠; 550.一把伞南星;553.宜昌木姜子;560.异叶榕;561.异叶蛇葡萄;573.油茶;592.长裂葛萝槭;613.周毛悬钩子;620.紫萼;630.棕榈图4 物种与地形因子的CCA双序图Elve. Altitude;Slop. Slope;Aspe. Aspect;Posi. Slope position;21.Symplocos anomala;31.Corylus ferox var. thibetica;37.Hedera nepalensis var. sinensis;58.Gentianopsis scabromarginata;65.Carex taliensis;74.Cyrtomium maximum;80.Lonicera acuminata;83.Agapetes lacei;86.Lysimachia punctatilimba;98.Tilia tuan;101.Acer caudatum;105.Schefflera octophylla;110.Spiraea veitchii;111.Rhododendron auriculatum;113.Stellaria media;129.Polystichum neolobatum;132.Davidia involucrata;133.Castanopsis tibetana;140.Cyrtomium fortunei;166.Caulophyllum robustum;170.Malus hupehensis;185.Sorbus wilsoniana;194.Pistacia chinensis;197.Tiarella polyphylla;209.Polygonum thunbergii;222.Fargesia spathacea;238.Arthraxon hispidus;247.Pteridium aquilinum;261.Hydrangea strigosa;270.Pittosporum trigonocarpum;277.Carex fargesii;288.Galium asperuloides;296.Pyrola calliantha;299.Astilbe chinensis;301.Lindera neesiana;303.Pinus massoniana; 304.Rhododendron ovatum; 307.Rhododendron mariesii;313.Dicranopteris dichotoma;315.Decaisnea insignis;322.Castanea seguinii;338.Sphagnum palustre;346.Cotoneaster bullatus;358.Fagopyrum esculentum;363.Cyclobalanopsis glauca;377.Carex japonica;389.Lindera obtusiloba;401.Albizia kalkora;402.Lindera glauca;403.Litsea cubeba;414.Cunninghamia lanceolata;424.Calanthe brevicornu;428.Lycopodium japonicum;440.Tetracentron sinense;442.Ilex szechwanensis;448.Schefflera delavayi;475.Euonymus alatus;497.Oenanthe dielsii;518.Juncus bufonius;519.Lyonia ovalifolia var. elliptica;524.Photinia parvifolia; 550.Arisaema erubescens;553.Litsea ichangensis;560.Ficus heteromorpha;561.Ampelopsis heterophylla;573.Camellia oleifera;592.Acer grosseri var. hersii;613.Rubus amphidasys;620.Teucrium tsinlingense;630.Trachycarpus fortunei Fig.4 CCA biplot of species and topographic factors
本研究CCA排序的前两轴累积物种-环境关系方差所占比重近60%,说明前两轴集中了大部分的信息反映地形对植物物种分布格局的影响,排序结果见图4。从排序图中可以看出,物种排列在第1和第2排序轴平面的不同位置,反映了该物种适宜的生境条件。海拔、坡位沿CCA 1轴从左向右逐渐降低。坡度沿CCA 2轴从上到下逐渐增大。杉木(Cunninghamialanceolata)、钩栲(Castanopsistibetana)、马尾松(Pinusmassoniana)、油茶(Camelliaoleifera)、薄叶山矾(Symplocosanomala)、青冈(Cyclobalanopsisglauca)等物种分布在排序轴的右上角,说明这些物种主要分布在海拔较低、坡度较缓的地段;珙桐(Davidiainvolucrata)、绿叶甘橿(Linderaneesiana)、水青树(Tetracentronsinense)、椴树(Tiliatuan)等物种分布在CCA 2排序轴的下部,其分布受坡度的影响较大,即在坡度较陡的地段分布较多;茅栗(Castaneaseguinii)、箭竹(Fargesiaspathacea)、宜昌木姜子(Litseaichangensis)、腊莲绣球(Hydrangeastrigosa)等物种受海拔和坡位的影响较大,主要出现在高海拔、高坡位的区域;泥炭藓(Sphagnumpalustre)、大理苔草(Carextaliensis)、湖北海棠(Malushupehensis)、石松(Lycopodiumjaponicum)、戟叶蓼(Polygonumthunbergii)、紫萼(Teucriumtsinlingense)等物种分布在CCA 1排序轴的左端,其分布受海拔、坡位、坡向的影响较大,主要分布在海拔较高、坡度较缓、较冷湿的山顶台地。
4 讨 论
物种多样性及分布与环境因子的关系受研究尺度的影响较大,在大尺度上,气候是主导因子,在较小尺度上,地形因子起主要作用[3,6]。本研究发现,海拔、坡度等地形因子与物种丰富度具有显著相关性(P<0.05),海拔作为间接的环境因子通过影响水分、温度等的空间分布从而影响物种丰富度分布。许多研究表明,物种丰富度随海拔梯度的变化先增大后减小的单峰格局[5,16-17]或随海拔升高呈递减的趋势[18-19],而本研究中物种丰富度随海拔的升高呈递增的趋势,该研究结果与前人研究的一般规律不同。通过野外调查发现,该区高海拔地段(约1 800~2 000 m)分布大面积泥炭藓湿地,湿地是陆生生态系统和水生生态系统之间的过渡地带,陆生和湿生植物生长旺盛,是生物多样性最丰富的地区之一[20],因此湿地的分布是导致该区高海拔地区物种丰富度较高的原因。坡度则是水分和土壤养分流动的驱动力,对土壤厚度、土壤理化性质等具有重要影响[3],常用来解释物种丰富度的分布格局[6],但研究区不同得出的结论也有差异,而本研究结果显示,植物物种丰富度随坡度的增加呈先减小后增大再减小的趋势,在缓坡和较陡坡物种丰富度最高,这一规律与张昌顺等[3]和Pinder等[21]的研究结果一致。实地调查发现,在七姊妹山自然保护区缓坡地带大部分是泥炭藓湿地分布的山顶台地,植物种类较丰富;较陡坡由于地势陡峻,人为干扰较小,原生植被保存较好,植物丰富度较高。
研究表明,不同生活型植物对海拔的敏感度不同,因而其物种丰富度可能具有不同的海拔梯度格局[5,22]。通常情况,低海拔段,植物生长季节所需热量充足而水分不足,高海拔段,水分充足而热量可能成为限制性因子,高大乔木生长较少,低矮灌、草物种生长旺盛,而中海拔段水热组合较好,最适宜植物生长[23]。本研究七姊妹山自然保护区3种生活型植物物种丰富度表现出同样的规律,在低海拔段,乔、灌、草3种生活型植物丰富度都较低但灌木物种丰富度高于乔木和草本,到达中海拔段,3种生活型植物物种丰富度都较高,而当到高海拔段时,草本植物物种丰富度迅速增大且高于灌木和乔木。
CCA排序的第1轴和第2轴主要反映海拔和坡度梯度的变化,第3轴和第4轴主要反映坡位和坡向的梯度变化,4轴共解释了物种-环境关系方差的100%,这可能与自然保护区的核心区森林群落中,人为干扰较少,土壤因子含量趋于稳定,海拔、坡度等地形因子对物种的分布起主要作用有关[24]。地形因子包括海拔、坡度、坡向、坡位等它们共同影响水分、温度、养分、光照等的空间分布,从而影响物种的分布格局[3]。本研究结果表明,海拔是影响该区物种分布最重要的地形因子,其次为坡度,因此,按地形因子对七姊妹山自然保护区植物分布影响力的大小排序为海拔>坡度>坡位>坡向,这与李帅锋等[1]对云南思茅松林群落、邵方丽等[15]对北京山区防护栏优势种群、刘秋峰等[25]对中条山混沟地区影响群落物种分布的主要地形因子的研究结果相似。
[1] 李帅锋,苏建荣,刘万德,等. 云南省思茅松林群落数量分类及物种多样性与自然环境的关系[J].生态学杂志,2013,32(12):3 152-3 159.
LI S F, SU J R, LIU W D,etal. Quantitative classification ofPinekesiyavar.langbianensiscommunities and their species richness in relation to the environmental factors in Yunnan Province of Southwest China [J].ChineseJournalofEcology, 2013, 32(12):3 152-3 159.
[2] 许 涵,李意德,骆土寿,等. 海南尖峰岭不同热带雨林类型与物种多样性变化关联的环境因子[J]. 植物生态学报,2013,37(1):26-36.
XU H, LI Y D, LUO T S,etal. Environmental factors correlated with species diversity in different tropical rain forest types in Jianfengling, Hainan Island, China[J].ChineseJournalofPlantEcology, 2013, 37(1):26-36.
[3] 张昌顺,谢高地,包维楷,等. 地形对澜沧江源区高寒草甸植物丰富度及其分布格局的影响[J]. 生态学杂志,2012,31(11):2 767-2 774.
ZHANG C S, XIE G D, BAO W K,etal. Effects of topographic factors on the plant species richness and distribution pattern of alpine meadow in source region of Lancang River, Southwest China[J].ChineseJournalofEcology, 2012, 31(11):2 767-2 774.
[4] CANTON Y, BARRIO G D, SOLE-BENET A,etal. Topographic controls on the spatial distribution of ground cover in the Tabernas badlands of SE Spain[J].Catena, 2004, 55(3):341-365.
[5] 王志恒,陈安平,朴世龙,等. 高黎贡山种子植物物种丰富度沿海拔梯度的变化[J].生物多样性,2004,12(1):82-88.
WANG Z H, CHEN A P, PIAO S L,etal. Pattern of species richness along an altitudinal gradient on Gaoligong Mountains, Southwest China [J].BiodiversitySince, 2004, 12(1):82-88.
[6] 任学敏,杨改河,朱 雅,等. 环境因子对太白山高山植被物种组成和丰富度的影响[J].生态学报,2014,34(23):6 993-7 003.
REN X M, YANG G H, ZHU Y,etal. Effect of environmental variable on species composition and richness of alpine vegetation in Taibai Mountain [J].ActaEcologicaSinica, 2014, 34(23):6 993-7 003.
[7] RAHBEK C, GRAVES G R. Multiscale assessment of patterns of avian species richness [J].ProceedingsoftheNationalAcademyofSciencesoftheUnitedStatesofAmerica, 2001, 98(8):4 534-4 539.
[8] TAMBE S, RAWAT G S, HURNI H,etal. The alpine vegetation of the Khangchendzonga landscape, Sikkim Himalaya: community characteristics, diversity, and aspects of ecology [J].MountainResearch&Development, 2015, 30(3):266-274.
[9] 刘胜祥,瞿建平,汪正祥,等. 湖北七姊妹山自然保护区科学考察与研究报告[R]. 武汉: 湖北科学技术出版社,2006: 2-3.
LIU S X, QU J P, WANG Z X,etal. Hubei Qizimeishan Nature Reserve scientific survey and research report[R]. Wuhan: Hubei Science and Technology Press, 2006, 2-3.
[10] 满金山,彭宗林,方元平,等.七姊妹山国家级自然保护区野生植物资源及其开发利用[J].安徽农业科学,2008,36(12):5 119-5 120+5 123.
MAN J S, PENG Z L, FANG Y P,etal. Study on the wild plant resource in Qizimeishan Nature Reserve and its exploitation[J].JournalofAnhuiAgricultureScience, 2008, 36(12):5 119-5 120+5 123.
[11] 满金山,彭宗林,方元平,等. 七姊妹山国家级自然保护区菊科植物区系及资源植物分析[J]. 生态科学,2008,27(1):38-41.
MAN J S, PENG Z L, FANG Y Y,etal. Analysis of flora and resources plants on compositae in Qizimeishan Nature Reserve [J].Ecologicalscience, 2008, 27(1):38-41.
[12] 方元平,刘胜祥,汪正祥,等. 七姊妹山自然保护区野生保护植物优先保护定量研究[J]. 西北植物学报,2007, 27(2):348-355.
FANG Y P, LIU S X, WANG Z X,etal. Quantitative assessment of priority for conservation of the National protected plant in Qizimei Mountain Nature Reserve[J].ActaBotanicaBoreali-OccidentaliaSinica, 2007, 27(2):348-355.
[13] 何 俊,汪正祥,雷 耘,等. 七姊妹山自然保护区台湾水青冈林群落学特征研究[J]. 华中师范大学学报(自然科学版), 2008,42(2):272-277.
HE J, WANG Z X, LEI Y,etal. The study on coenological characteristics ofFagushayataecommunity in Qizimei Mountain Nature Reserve [J].JournalofHuazhongNormalUniversity(Nat,Sci.),2008, 42(2):272-277.
[14] 沈泽昊,方精云. 基于种群分布地形格局的两种水青冈生态位比较研究[J]. 植物生态学报,2001,25(4):392-398.
SHEN Z H, FANG J Y. Niche comparison of twoFagusspecies based on the topographic patterns of their populations[J].ActaPhytoecologicaSinica, 2001, 25(4):392-398.
[15] 邵方丽,余新晓,郑江坤,等. 北京山区防护林优势树种分布与环境的关系[J]. 生态学报,2012,32(19):6 092-6 099.
SHAO F L, YU X X, ZHENG J K,etal. Relationships between dominant arbor species distribution and environmental factor of shelter forests in the Beijing mountain area[J].ActaEcologicaSinica, 2012, 32(19):6 092-6 099.
[16] 王志恒, 陈安平, 方精云. 湖南省种子植物物种丰富度与地形的关系[J]. 地理学报,2004,59(6):889-894.
WANG Z H, CHEN A P, FANG J Y. Richness of seed plant in relation with topography in Hu’nan province[J].ActaGeographicaSinica, 2004, 59(6):889-894.
[17] 池秀莲, 唐志尧. 面积、温度及分布区限制对物种丰富度海拔格局的影响:以秦岭太白山为例[J]. 植物生态学报,2011,35(4):362-370.
CHI X L, TANG Z Y. Effects of area, temperature and geometric constraints on elevational patterns of species richness: a case study in the Mountain Taibai, Qinling Mountains, China[J].ChineseJournalofplantEcology, 2011, 35(4):362-370.
[18] 唐志尧,方精云,张 玲. 秦岭太白山木本植物物种多样性的梯度格局及环境解释[J]. 生物多样性,2004,12(1):115-122.
TANG Z Y, FANG J Y, ZHANG L. Patterns of woody plant species diversity along environmental gradients on Mt. Taibai, Qinling Mountains[J].BiodiversityScience, 2004, 12(1):115-122.
[19] OHSAWA M. Latitudinal comparison of altitudinal changes in forest structure, leaf-type, and species richness in humid monsoon Asia[J].GlobalChange&TerrestrialEcosystemsinMonsoonAsia, 1996, 33(1-2):3-10.
[20] 杨文斌,刘 坤,周守标.安徽新安江干流滩涂湿地草本植物区系及物种多样性[J].生态学报,2013,33(5):1 433-1 442.
YANG W B, LIU K, ZHOU S B. The flora and diversity of herbaceous seed plants in wetlands along the Xinanjiang river from Anhui[J].ActaEcologicaSinica, 2013, 33(5):1 433-1 442.
[21] III J E P, KROH G C, WHITE J D,etal. The relationships between vegetation type and topography in Lassen Volcanic National Park [J].PlantEcology, 1997, 131(1):17-29.
[22] WANG G, ZHOU G, YANG L,etal. Distribution, species diversity and life-form spectra of plant communities along an altitudinal gradient in the northern slope of Qilianshan Mountains, Gansu, China[J].PlantEcology, 2003, 165(2):169-181.
[23] 王国宏. 祁连山北坡中段植物群落多样性的垂直分布格局[J]. 生物多样性,2002,10(1):7-14.
WANG G H. Species diversity of plant communities along an altitudinal gradient in the middle section of Northern slope of Qilian Mountains, Zhangye, Gansu, China[J].BiodiversityScience, 2002, 10(1):7-14.
[24] 白玉宏,张钦弟,毕润成. 山西接骨木群落分布及其与环境因子的关系[J]. 林业科学,2013,49(12):18-24.
BAI Y H, ZHANG Q D, BI R C. Relation betweenSambucuswilliamsiicommunity distribution and the environmental factors in Shanxi[J].ScientiaSilvaeSinicae, 2013,49(12):18-24.
[25] 刘秋锋,康慕谊,刘全儒.中条山混沟地区森林乔木种的数量分类与环境解释[J].植物生态学报,2006,30(3):383-391.
LIU Q F, KANG M Y, LIU Q R. Quantitative classification and environmental interpretation of forest tree species in Hungou, Zhongtiao Mountain[J].JournalofPlantEcology, 2006, 30(3):383-391.
(编辑:潘新社)
Effect of Topography on Plant Species Richness and Distribution in Qizimei Mountains Nature Reserve
XIONG Binmei1, LEI Yun2, WANG Zhengxiang1,3,4*, LI Tingting1,XIONG Xiuhai5, LI Zhongqiang1,3,4, ZHANG E1, PENG Zonglin6
(1 College of Resources and Environmental Science, Hubei University, Wuhan 430062, China; 2 College of Life Sciences, Central Normal University, Wuhan 430070, China; 3 Hubei Collaborative Innovation Center for Green Transformation of Bio-Resources, Wuhan 430062, China; 4 Hubei Province Key Laboratory of Regional Development and Environment Response, Wuhan 430062, China; 5 School of Resource and Enviromental Sciences, Wuhan University, Wuhan 430072, China; 6 Area Management Bureau of the Qizimei Mountains Nature Reserve, Xuan’en, Hubei 445500, China)
Species richness and distribution are closely related to topography factors (altitude, slope grade, slope aspect, slope position), and study their relationship of Qizimei Mountains Nature Reserve is important to maintain and restore the plant diversity and the management of forest. Phytosociological method was used to investigate plant communities in Qizimei Mountains Nature Reserve. Furthermore, the regression analysis and canonical correspondence analysis were used to study the effect of topography on species richness and distribution pattern. The results showed that: (1) there were 633 plant species and it was belonging to 133 families and 316 genera. And it contained 118 species of trees, 150 species of shrubs and 365 species of herbs in 40 sample plots(20×20 m2). (2) The results of curve fitting indicated that the species richness was highly significantly correlated to altitude and slope (P<0.05), the species richness increase with increasing altitude and had a tendency with slope gradient that was after reducing first increase and then decrease. (3) The plant life form showed, at all altitudes, tree species richness was consistently lower than that of shrubs and herbs. Moreover, shrub species richness was higher than that of trees and herbs in low and mid-low altitude zones. Besides, species richness of herbs in middle and high altitude areas was larger and higher than that of trees and shrubs. (4) Canonical correspondence analysis showed that the topographic factors had a great effect on species distribution (P= 0.002<0.01) , and according to the impact intensity ranked as altitude>slope>slope position>slope aspect, which indicated that altitude is the most significant factor for affecting the distribution of species in Qizimei Mountains Nature Reserve.
topography; species richness; species distribution; canonical correspondence analysis (CCA); Qizimei Mountains Nature Reserve
1000-4025(2016)11-2307-07
10.7606/j.issn.1000-4025.2016.11.2307
2016-08-10;修改稿收到日期:2016-08-29
国家自然科学基金(41471041);七姊妹山自然保护区管理局保护研究专项
熊斌梅(1990-),女,硕士研究生,主要从事生物多样性的保护与管理研究。E-mail:1014132088@qq.com
*通信作者:汪正祥,教授,博士生导师,主要从事植被生态、生物多样性的保护、受损生态系统的诊断与恢复研究。E-mail:wangzx66@hubu.edu.cn
Q948.114
A
