冠层高度对毛竹叶片光合生理特性的影响
2017-01-04曹永慧周本智王小明顾连宏
曹永慧,周本智*,王小明,顾连宏
(1 中国林业科学研究院亚热带林业研究所,国家林业局钱江源森林生态系统定位观测研究站,浙江富阳 311400; 2 美国橡树岭国家实验室环境科学部,田纳西州,橡树岭 37831)
冠层高度对毛竹叶片光合生理特性的影响
曹永慧1,周本智1*,王小明1,顾连宏2
(1 中国林业科学研究院亚热带林业研究所,国家林业局钱江源森林生态系统定位观测研究站,浙江富阳 311400; 2 美国橡树岭国家实验室环境科学部,田纳西州,橡树岭 37831)
借助LI-6400便携式光合作用系统,研究了冠层高度对不同林龄毛竹(Phyllostachyspubescens)叶片光合生理特性和水分利用效率(WUE)的季节性影响,为促进毛竹林碳汇能力和生产力提升的林分结构调整等可持续栽培技术提供理论依据。结果表明:(1)出笋期,不同竹龄毛竹叶片净光合速率(Pn)和蒸腾速率(Tr)的日均值呈现出冠层上部小于冠层下部的梯度变化趋势,且2a生毛竹不同冠层Pn日均值大于3a生毛竹;孕笋行鞭期,不同林龄毛竹各时间点Pn值和日均值、以及2年生毛竹各时间点的Tr值均为冠层上部大于冠层下部。各生长季节,不同林龄毛竹个体叶片的气孔导度(Gs)均与Tr的变化趋势一致。(2)2年生毛竹各季节仅冠层上部叶片会出现“光合午休”,而3年生毛竹仅于出笋期时各冠层叶片出现“光合午休”现象。(3)出笋期毛竹叶片WUE日均值随着冠层高度增加而增加,这种变化趋势不受竹龄影响;而孕笋行鞭期,仅2年生毛竹叶片WUE日均值随着冠层高度增加而下降。不同冠层高度的孕笋行鞭期毛竹叶片WUE日均值都显著高于出笋期;冠层高度对毛竹叶片气体交换特性和WUE的影响受生长发育关键期的季节因素影响,且毛竹叶片WUE与Gs之间存在负相关关系,其不受毛竹个体年龄和叶片冠层高度影响。(4)不同生长季节各冠层叶绿素a/b值均随着冠层高度下降而降低,不同林龄毛竹叶片叶绿素含量基本随着冠层自上而下呈逐渐增加的趋势。各生长季节,不同林龄个体叶片氮素含量、比叶重随冠层高度垂直变化趋势与叶片Pn日均值的垂直变化趋势一致。研究认为,毛竹不同冠层部位叶片通过改变形态、氮素含量来适应不同生长季节生长环境的变化,以便充分利用光能提高光合能力。
毛竹;光合作用;蒸腾作用;水分利用效率;冠层高度;季节差异;日变化
竹子广泛分布于东南亚、非洲和拉丁美洲。竹林具有重要的生态系统服务功能,在陆地生态系统碳循环中发挥着重要作用。中国是竹林分布面积最大、竹资源种类最丰富的国家之一,在竹子生产和利用方面有着悠久的历史[1]。中国约有500个竹种,从竹子分布、竹材价值和其他经济价值方面来论,毛竹(Phyllostachyspubescens)是最重要的竹种[2-3]。近年来,随着全球气候变化和森林生态系统的碳循环领域研究的深入,毛竹经营面积迅速增加,毛竹产业实现蓬勃发展。中国现有毛竹经营面积超过400万hm2,占世界总量的90%以上[4-7]。毛竹林碳储量达到约611.15 Tg,约占全国森林总碳储量的7.8%[3]。
毛竹作为重要固碳树种,生长迅速,每年的生物量积累非常巨大。其生长很独特,从幼笋出土至长成幼竹只需短短几个月的时间即完成秆行生长,可迅速生长达到树冠(通常>10 m)[8];这是因为竹丛具有发达的鞭根系统,这使新老竹之间可通过鞭根系统实现碳和养分运输互连[9]。我们早期研究表明,在9月份不同的光合有效辐射(PAR)条件下,3年生竹子冠层上部和中部叶片的净光合速率(Pn)显著高于冠层下部的[10]。然而,在重要的生长时期,毛竹叶片固碳能力、蒸腾速率和水分利用效率的时空动态变化仍在很大程度上存在不确定性,这在出笋期和茎秆伸长期尤其明显[11]。毛竹发笋成竹主要在3月底到5月初这个阶段完成。在毛竹出笋后的幼竹生长发育过程中,其叶片尚未展开,叶片光合功能尚未完善,此时光合同化物主要靠母竹供给,可见这段时间内毛竹母竹个体的光合作用能力大小直接影响了笋期幼竹快速生长、持续获取养分的水平。孕笋行鞭期也是竹子生长发育的重要阶段,一般从10月下旬开始至12月中旬结束。此时,毛竹发达鞭根系统对营养物质和水分需求旺盛,对土壤条件依赖比较强。竹林地形复杂,灌溉困难,因此,在秋季和冬季出现的季节性干旱会阻碍竹子生长,在有限的水资源条件下,提高竹林水分利用效率是提高竹林生产力水平的关键。
太阳光辐射通量从植物冠层上部到下部是指数式衰减,引起了冠层空气温度和湿度的相应变化。叶片的形态结构、叶绿素含量、氮素含量以及光合速率等光合生理特性会对不同冠层高度外界环境条件的改变发生响应。因此,太阳光辐射通量势必会造成植物叶片水分利用效率的昼夜和季节变化。迄今为止,很少有学者针对不同年龄毛竹或不同冠层高度毛竹叶片的光合固碳能力和水分利用效率开展研究,对于毛竹关键生长发育期间冠层水平叶片光合作用的研究报道更少[4,11-12]。有研究表明毛竹碳固定峰值分别出现在6月和11月[11],然而目前对毛竹关键生长期出笋期(5月)和孕笋行鞭期(12月)的碳固定能力研究较少。因此,本试验借助毛竹林内自行设计搭建的30 m高的综合观测铁塔,以毛竹不同年龄个体为研究对象,在毛竹关键发育阶段(5月上旬和12月上旬),对其不同冠层高度水平的叶片光合作用、水分利用效率和叶片形态、生理特性进行了原位比较研究,旨在阐明冠层高度对毛竹叶片光合生理特性潜在影响及其叶片水分利用效率对气孔导度的响应特性,为毛竹林林分结构调整、高效栽培等科学经营方式的深入研究提供理论依据。
1 材料和方法
1.1 研究地区自然概况
本研究在国家林业局钱江源森林生态系统定位研究站庙山坞辅站进行,其位于浙江杭州西郊富阳区境内(119°56′~120°02′ E,30°03′~30°06′N)。该区域地形属浙西低山丘陵区天目山系余脉,属于中亚热带季风气候区,年平均气温16.1 ℃(最高气温40.2 ℃,最低气温-14.4 ℃),多年平均降水量1441.9 mm。土壤属于酸性红壤。
1.2 试验材料与样地设置
试验区毛竹林为20世纪60年代种植,处于粗放经营的自然更新林。本试验在自然生长毛竹人工林内构建一座30 m高的综合观测塔。2年生和3年生毛竹个体随机分布在铁塔周围。本研究在观测铁塔周围随机选取3块面积为400 m2(20 m×20 m)的人工毛竹林标准样地,样地坡度20°左右,坡向正南,海拔169 m。采用每木检测的方法分别测定样地内竹株的胸径和竹高。样地内竹林密度为3 875株·hm-2,郁闭度0.95,竹株胸径分布在4.0~13.6 cm之间,平均胸径为9.8 cm,平均竹高13.2 m。林下几乎没有灌木和草本,但地表覆盖一定的凋落物,平均厚度约2.0 cm。每2年劈山1次,不施肥,不翻耕,只采挖春笋。
1.3 测定指标及方法
1.3.1 光合气体交换参数 本试验分别于不同年龄毛竹各选取3株个体进行气体交换参数日变化进程测定。测定和采样时间分别为2013年5月上旬(出笋期)和12月上旬(孕笋行鞭期)。测定选择天气晴朗的条件下进行,每个样株分别在不同冠层高度各随机选取3片长势相同、充分展开的向阳叶片(每个叶片重复测定3个数据,最终取平均值)。采用 LI-6400 便携式光合测定系统(LI-COR, Lincoln, USA)从 8:00 ~18:00 每2 h测定1次,每个测定重复3次。为了使测定具有可比性,所有毛竹样株的叶片气体交换参数测定尽量在同一天内同一时间范围内完成;测定好一个样株的不同冠层部位的第一片样叶后,再测定另外样株的不同冠层部位第一片样叶;然后马上依次开始各样株不同冠层部位的第二和第三片样叶的测定,最终取各参数重复测定的平均值作为该参数各时间点值。不同参数各时间点的测定值取平均值作为该参数日平均值(表1)。仪器自动记录净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)等数值。光合参数测定时,CO2浓度以当地环境CO2浓度(400 μmol·mol-1)为基准,气体流量控制在0.5 L·min-1,相对湿度为60%±4%,样品室叶片温度在出笋期(5月上旬)和孕笋行鞭期(12月上旬)分别控制在(20± 3)℃和(15± 3)℃。然后利用公式(Pn/Tr)计算瞬时水分利用效率(WUE)。
1.3.2 光合色素含量和叶形态参数 每次光合参数测量结束后,测定所用叶片做标记后放入存储袋带回室内用于叶绿素含量、叶氮含量和叶片面积测定。其中,叶片面积测定利用扫描仪(CB50EJ,Cambridge,UK)扫描后再用Delta-T Scan(CB50EJ,Cambridge,UK)软件计算实际的叶面积,最后按照实际叶面积计算样品实际的各项生理指标;光合色素含量按照李合生的方法[13]测定,每个处理取叶片样品0.1~0.2 g,按照丙酮和乙醇1∶1的比例混合浸泡提取,重复3次;提取叶片色素后,用分光光度计测定吸光度值,计算叶绿素(Chl a、Chl b)含量和类胡萝卜素(Cars)的含量,最终取3次重复测定数据的平均值。从同样的每株植物上按照不同冠层部位,分别采集30片完全展开的健康叶片,分为3份,分别用千分尺测量叶片厚度并称取鲜重,用扫描仪CB50EJ测定叶片面积,然后带回实验室用烘箱烘干(105 ℃下杀青30 min,然后80 ℃连续烘干至恒重),称量烘干重,然后计算比叶干重(LMAdw,叶干重/叶面积)和比叶鲜重(LMAfw,叶鲜重/叶面积),取3次重复测定的平均值。试验同时,不同年龄植株另外分别在冠层部位取若干数量叶片带回实验室,用烘箱烘干并磨碎过100目筛,然后经浓硫酸-过氧化氢溶液消煮,消煮液采用Foss 凯氏定氮仪叶片全氮含量(Nmass)。
1.4 数据处理
毛竹叶片的光合色素、叶形态参数和光合参数之间的差异,采用Excel 2007进行数据处理,各参数以平均值加减标准误(mean±SE)表示。气体交换参数的计算运用Photosynthesis Assistants软件。
2 结果与分析
2.1 冠层高度对毛竹个体光合气体交换参数和水分利用效率的影响
2.1.1 净光合速率 毛竹个体净光合速率(Pn)日变化存在季节差异和冠层高度差异(图1)。出笋期,冠层上部叶片Pn日变化在不同年龄毛竹个体间均呈明显的“双峰”曲线,第一个峰值3a生个体比2a生个体提早在10:00左右出现,第二个峰值均出现在15:00左右;冠层下部叶片Pn日变化在2a生毛竹个体呈明显的“单峰”曲线,而在3a生个体呈明显的“双峰”曲线(图1,A);同时,不同竹龄个体叶片Pn日均值均表现出冠层上部<冠层下部的垂直变化趋势,即Pn日均值随着冠层高度增加而下降(表1),且2a生毛竹个体不同冠层Pn日均值显著大于3a生毛竹个体(P<0.05)。孕笋行鞭期,3a生毛竹个体不同冠层叶片Pn日变化均呈明显的“单峰型”曲线,而2a生毛竹个体冠层上部叶片Pn日变化为“双峰型”曲线,下部叶片Pn日变化基本为“单峰型”曲线;在这个生长季节,3a和2a生毛竹个体冠层叶片Pn峰值多在12:00左右出现,2a生毛竹个体冠层上部叶片“光合午休”现象亦在12:00左右出现(图1,B);与出笋期不同,孕笋行鞭期不同年龄(无论竹龄大小)毛竹一天内各时间点冠层上部叶片Pn值均大于冠层下部叶片,即Pn日均值随着冠层高度增加而增加(表1),且相同冠层3年生毛竹个体叶片的Pn日均值较高。方差分析结果(表1)显示,无论那个生长季节,毛竹个体叶片Pn日均值在相同竹龄个体的不同冠层之间以及相同冠层部位不同竹龄个体之间差异均达显著水平(P<0.05)。
2.1.2 蒸腾速率 图2,A显示,在出笋期,2a、3a生毛竹个体冠层上部和下部叶片蒸腾速率(Tr)日变化亦均表现出明显的“双峰型”曲线,2a生个体各冠层叶片Tr值第一次峰值均出现在12:00左右,3a生个体第一次峰值则比2a生个体提早出现;2a、3a生毛竹个体叶片各时间点Tr值和Tr日均值同样表现出冠层上部小于冠层下部的垂直变化趋势,即不同竹龄个体叶片Tr日均值随着冠层高度增加而下降,且Tr日均值随冠层垂直梯度变化比Pn日均值更显著(表1)。方差分析结果(表1)显示,出笋期毛竹个体叶片Tr日均值在相同竹龄不同冠层之间差异达显著水平(P<0.05)。从图2,B可知,在孕笋行鞭期,3a生毛竹个体冠层上部和下部叶片Tr日变化均呈明显的“单峰型”曲线,峰值点在12:00左右出现,各时间点Tr值在冠层叶片间差异较小;同期2a生毛竹个体冠层下部叶片Tr日变化为“单峰型”曲线,冠层上部叶片则为“双峰型”曲线,且冠层上部叶片各时间点Tr值大于冠层下部叶片;在孕笋行鞭期,2年生毛竹个体Tr日均值随着冠层高度增加而增加,而3年生毛竹个体随冠层增加略微下降(表1)。Tr日均值在相同冠层部位不同竹龄个体之间差异均达显著水平(P<0.05)。

3a-d 和3a-u分别表示3年生个体冠层的下部和上部叶片,而2a-d 和2a-u分别表示2年生个体冠层的下部和上部叶片;下同图1 出笋期(A)和孕笋行鞭期(B)毛竹不同竹龄和冠层叶片净光合速率(Pn)日变化特征3a-d and 3a-u stand for lower (d) and upper (u) canopy leaves of 3 years old (3a) plant ,while 2a-d and 2a-u stand for leaves of 2 years old (2a) plant; The same as below.Fig.1 The diurnal and seasonal change of net photosynthetic rate (Pn) in leaves of P.pubescens during the culm elongation period(A) and shoot development period(B)

图2 出笋期(A)和孕笋行鞭期(B)毛竹不同冠层叶片蒸腾速率(Tr)日变化Fig.2 The diurnal and seasonal transpiration rate (Tr) change for different canopy leaves of P.pubescens during the culm elongation period (A) and shoot development period (B)
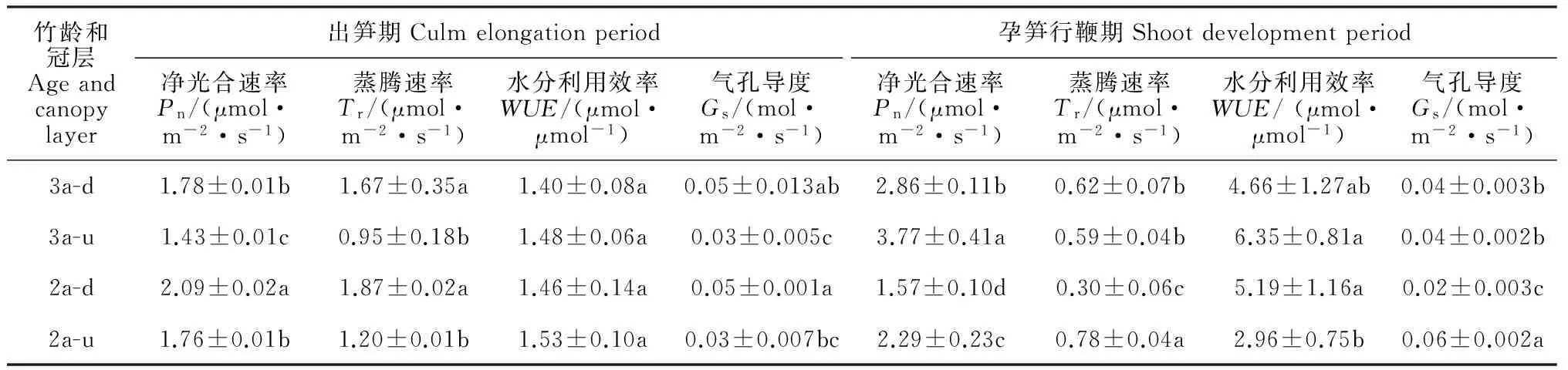
表1 不同冠层高度毛竹个体叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和瞬时水分利用效率(WUE)日均值比较
注:表中数值为平均值±标准偏差,同列不同字母表示不同竹龄和冠层处理下某指标在0.05水平差异显著(P<0.05);下同
Note: Data in the table are means ± SD. The results with same normal letters in the same column are not significantly different at 0.05 level among different canopies and ages of bamboo. The same as below

图3 出笋期(A)和孕笋行鞭期(B)毛竹不同冠层叶片气孔导度(Gs)日变化Fig.3 Stomatal conductance (Gs) for 1 year-old leaves of 2 and 3 year-old P. pubescens among different canopy layers during the culm elongation period(A) and shoot development period(B)
2.1.3 气孔导度 在出笋期,不同竹龄毛竹不同冠层叶片气孔导度(Gs)日变化趋势与其Tr日变化一致;3a、2a生毛竹冠层下部和上部叶片Gs日变化均呈明显的“双峰型”曲线,峰值出现时间与蒸腾速率一致,且冠层下部叶片Gs值各时间点均高于冠层上部(图3,A)。不同竹龄个体Gs日均值冠层下部均显著高于上部(P<0.05)(表1)。图3,B显示,在孕笋行鞭期,2a生个体不同冠层Gs日变化均呈“双峰型”曲线,峰值均在10:00和14:00左右出现,且其冠层上部叶片各时间点Gs值均大于冠层下部叶片;与2a生个体不同,3a生个体不同冠层叶片Gs日变化为“单峰型”曲线,峰值在12:00左右出现,其各时间点Gs值在冠层之间差异较小。但2a生个体Gs日均值为冠层上部显著大于下部,且相同冠层不同竹龄个体之间差异显著(P<0.05)(表1)。综上所述,无论哪一个生长季节,不同竹龄毛竹个体叶片Gs与Tr变化趋势一致;出笋期毛竹冠层上部太阳辐射和温度增加迅速,导致叶片自我保护性关闭,进而降低气孔导度。
2.1.4 水分利用效率 在出笋期,不同竹龄个体一天内各时间点叶片水分利用效率(WUE)值均随着时间进程逐渐下降,傍晚时回升;各时间点WUE值基本表现出冠层上部高于下部的垂直变化趋势,且冠层差异在午后时间段更为明显;相同冠层高度,叶片各时间点WUE表现为3年生高于2年生个体的趋势(图4,A)。同时,不论竹龄大小,毛竹WUE日平均值随着冠层高度增加而增加(表1)。这是由于出笋期不同竹龄个体叶片的Pn和Tr日均值随着冠层高度增加而下降,且Tr日均值随冠层垂直梯度变化比Pn日均值更显著,因此在Pn和Tr的双重影响下,WUE日均值随着冠层高度增加而增加,但冠层之间差异较小。与出笋期WUE日变化明显不同,孕笋行鞭期各时间点WUE值随着时间进程逐渐上升,3年生毛竹个体各时间点WUE值为冠层上部略高于下部,而2年生毛竹正好相反(图4,B)。同时,3年生毛竹个体WUE日均值随冠层高度增加而增加,但差异不显著,而2年生毛竹WUE值随着冠层高度增加而下降,且冠层差异较显著;在相同冠层高度,冠层上部3年生毛竹个体的WUE值明显高于2年生毛竹个体的,而冠层下部则相反(表1)。这是因为孕笋行鞭期,与Pn日均值垂直梯度变化不同,2年生毛竹个体Tr日均值受冠层高度影响显著,而3年生毛竹个体则冠层之间差异较小,即Pn和Tr日均值将限制瞬时WUE值变化。另外,不论冠层高度如何,毛竹各生长季节叶片WUE日均值都为孕笋行鞭期显著高于出笋期。气孔导度(Gs)表示植物气孔的开张程度,它反映植物蒸腾耗水的大小。对叶片WUE日均值与Gs日均值之间相关性分析结果(表5)表明,不论哪个生长季节,不同竹龄个体不同冠层的叶片WUE与Gs之间存在显著负相关系。

图4 出笋期(A)和孕笋行鞭期(B)毛竹不同冠层叶片水分利用效率(WUE)日变化Fig.4 Water use efficiency (WUE) for 1 year-old leaves of 2 and 3 year-old P. pubescens among different canopy layers during the culm elongation period(A) and shoot development period(B)

图5 出笋期(A)和孕笋行鞭期(B)毛竹不同冠层叶片气孔导度(Gs)与瞬时水分利用效率(WUE)相关性Fig.5 The linear relationship between stomatal conductance (Gs) and WUE of P. pubescens among different canopy layers during the culm elongation period (A) and shoot development period (B)
2.2 冠层高度对毛竹个体叶片光合色素含量的影响
叶绿素含量可以作为反映植物衰老状况和光合能力的一个重要指标,在一定程度上叶绿素含量的高低会直接影响植物叶片捕获光的能力。如表2所示,在出笋期间,3年生和2年生毛竹个体不同冠层叶片叶绿素a/b值基本都随着冠层高度下降而降低;但叶绿素含量随冠层高度变化受竹龄影响,即3年生毛竹个体叶绿素a、叶绿素b和叶绿素总含量均随着冠层高度下降而呈增加趋势,但2年生毛竹个体叶绿素a、叶绿素b和叶绿素总含量则随着冠层高度下降呈降低的趋势;不同林龄毛竹叶绿素含量不同,其中3年生毛竹个体冠层上部叶片除了叶绿素a/b值外,其叶片叶绿素a、叶绿素b、叶绿素总含量均不同程度低于2年生毛竹个体,而其冠层下部叶片叶绿素含量均高于2年生个体,但差异不显著(表2)。由于叶绿素a/b值能反映植物耐阴能力大小,而3年生毛竹个体冠层上部叶片的叶绿素a/b值明显高于2年生的,说明出笋期3年生毛竹个体冠层上部叶片利用弱光的能力强于2年生毛竹。
同时,表2还显示,在孕笋行鞭期,不同林龄毛竹个体叶片叶绿素含量随着冠层高度的垂直变化而不同,3年生和2年生毛竹个体冠层上部叶片叶绿素a、叶绿素b、叶绿素(a+b)含量均不同程度低于冠层下部叶片,即随着冠层自上而下逐渐增加;与出笋期一致,叶绿素a/b值随冠层高度自上而下逐渐降低。综上所述,各生长季节,不同林龄毛竹叶绿素a/b值随冠层高度自上而下逐渐降低,而叶绿素a、叶绿素b、叶绿素(a+b)含量则基本随冠层高度下降而增加;说明各生长季节,毛竹个体不同冠层部位叶片捕获光能的能力基本表现出冠层上部低于下部的趋势,但其冠层上部叶片利用弱光的能力高于冠层下部叶片。
2.3 冠层高度对毛竹个体叶片比叶重和叶氮含量的影响
出笋期不同林龄毛竹个体叶片比叶重大小(LMA)基本表现出冠层下部高于冠层上部的趋势,仅2年生个体冠层差异显著(P<0.05,图6,A);而孕笋行鞭期则表现出冠层上部高于冠层下部的相反趋势,且相同年龄个体的冠层差异较显著,不同年龄个体的相同冠层之间差异也较明显(P<0.05,图6,B)。不同生长季节均是3年生毛竹个体各冠层叶片LMA值较高。同时,在出笋期,不同林龄毛竹个体单位重量叶片氮素含量(Nmass)与LMA值一致,亦表现出冠层下部高于冠层上部趋势,冠层差异显著(P<0.05,图7,A);在孕笋行鞭期,2年生毛竹个体叶片Nmass冠层变化趋势同出笋期,即表现出冠层下部高于冠层上部趋势,而3年生毛竹个体叶片Nmass与LMA变化类似,亦随冠层高度增加而增加,即冠层下部显著低于冠层上部(P<0.05,图7,B)。另外,与LMA变化不同,在不同生长季节,毛竹不同林龄个体各冠层叶片Nmass值均以2年生个体较高。以上结果说明出笋期,毛竹叶片通过增加冠层下部叶片比叶重和单位重量氮素含量来提高冠层下部的光合速率;孕笋行鞭期,冠层上部叶片叶绿素含量较低,其通过增加单位面积叶片重量和单位重量氮素含量来增加光能吸收,增加净光合速率,进而提高水分利用效率。

表2 出笋期和孕笋行鞭期毛竹不同冠层叶片叶绿素含量垂直变化

图6 出笋期(A)和孕笋行鞭期(B)2年生和3年生毛竹不同冠层叶片比叶重(LMA)的变化Fig.6 Leaf mass per area (LMA) for 1 year-old leaves of 2 and 3 year-old P. pubescens among different canopy layers during the culm elongation period (A) and shoot development period (B)

图7 出笋期(A)和孕笋行鞭期(B)2年生和3年生毛竹不同冠层叶片氮素含量(Nmass)的变化Fig.7 Leaf nitrogen content per mass (Nmass) for 1 year-old leaves of 2 and 3 year-old P. pubescens among different canopy layers during the culm elongation period (A) and shoot development period (B)
3 讨 论
3.1 冠层高度对毛竹个体光合气体交换参数和水分利用效率的影响
碳增益与水分损耗的比率(水分利用效率)是生态系统功能的一个重要特征,而生态系统功能又是全球水、能量和碳循环的核心[14-15]。因此,提高植物水分利用效率及其植物对水分可利用效率下降的应对能力已经成为重要的科学议程[16]。气孔最终控制了植物叶片和环境之间所有气体通量的95%。由此可见,气孔在减少水分流失中发挥重要调节作用。叶片与大气之间的气体交换是通过气孔控制吸收二氧化碳进行光合作用和蒸腾作用,进而决定了植物生产力及其水分利用效率[17]。太阳光辐射通量从植物冠层上部到下部是指数式衰减,引起了冠层空气温度和湿度的相应变化,从而导致不同冠层水平叶片净光合速率的显著差异。因此,太阳光辐射通量势必会造成植物(竹子)叶片水分利用效率的昼夜和季节变化。
毛竹是重要的固碳树种,经营面积迅速增加。近年来,随着全球气候变化和森林生态系统的碳循环领域越来越多的研究,毛竹林生态系统碳循环领域研究日益丰富。冠层导度是确定冠层蒸腾作用和光合作用的关键参数,为了探索由于竹林扩张造成的陆地碳水循环的潜在变化,日本学者将冠层导度这个参数用于日本西部地区毛竹的研究,发现与具有高叶面积指数的针叶林相比,毛竹林具有更高的蒸腾速率和光合作用能力,因此竹林替代针叶林后会使森林生态系统碳水循环发生较大变化[18]。然而,他们的研究依赖于林冠层蒸腾作用间接评价的液流通量方法。本研究采用了LI-6400便携式光合作用测定系统,借助30 m梯度观测铁塔直接测定分析评估毛竹林冠层光合作用和蒸腾作用的昼夜与季节变化,以及气孔导度和水分利用效率之间的关系,获取的数据更直接,更接近竹林自然生长状态。
Wen 等[11]研究报道5年生毛竹个体固碳能力显著高于3年生和当年生新竹,并研究和比较了3个林龄段毛竹个体叶片的最大净光合速率,但没有报道不同冠层叶片气孔导度和蒸腾速率对叶片水分利用效率的影响。本研究在对不同林龄毛竹个体叶片净光合速率、蒸腾速率和水分利用效率日变化和季节变化比较分析的基础上,探讨了在不同冠层高度毛竹叶片气孔导度对叶片水分利用效率的影响。
不同于落叶树种,毛竹不是年年换叶,而是每2年换1次叶(新竹在出笋当年完成放叶和展叶)。通常,每年换叶期有2个峰值,分别发生在春季(4月至5月)和秋末(11月)[19]。Ge 等[20]也报道毛竹林大小年的年总凋落物含量和年内凋落物第一次峰值均存在显著差异。本研究中试验毛竹林年龄结构组成主要以2年生和3年生毛竹个体为主。因此,在毛竹出笋期和孕笋行鞭期这两个重要生长期,2年生和3年生个体的叶片水分利用效率和气孔导度的变化格局从林地生产力提升和碳固定能力方面值得研究和探讨。本研究通过比较毛竹个体叶片水分利用效率的季节变化发现,无论冠层高度,孕笋行鞭期毛竹叶片水分利用效率日均值显著高于出笋期;在孕笋行鞭期,3年生毛竹个体叶片日平均水分利用效率随着冠层高度增加而增加,而2年生毛竹个体变化则相反,且这种水分利用效率随冠层梯度的变化格局不同于出笋期。这表明不同生长季节毛竹不同冠层高度叶片具有不同的水分利用策略,尤其在毛竹换叶期间;孕笋行鞭期的毛竹个体年龄结构组成将对叶片光合作用和水分利用效率大小有较大限制作用。然而,目前毛竹叶片水分利用效率受生长季节和竹龄影响的机制尚且不清楚,今后仍需进一步研究。
气孔调节对植物功能维持至关重要,尤其在水分供应有限时[16]。一般研究认为,具有较低气孔导度的叶片往往具备较高的水分利用效率,但这是以降低净光合速率和植物缓慢生长为代价的[17]。本研究也发现不论毛竹个体年龄和冠层高度,其叶片水分利用效率与气孔导度之间均存在较显著的负相关性;同时,在毛竹不同的生长季节,不同林龄毛竹个体可以通过调整不同林冠层叶片的气孔导度和蒸腾速率来提高叶片水分利用效率,从而有利于高效的光合作用和干物质积累。在毛竹孕笋行鞭期,降水量较低,温度适中,这些环境条件有利于气孔调节,降低蒸腾作用,提高净光合速率,达到较高的水分利用效率。然而,毛竹在出笋期表现出较低的水分利用效率和净光合速率,表明了水分胁迫可能造成毛竹叶片光合能力明显下降,而气孔控制是影响其光合作用的主要因素。另外,与前人研究结果[21]相似,本研究也发现出笋期毛竹冠层上部叶片气孔导度的下降会导致叶片蒸腾减少。由此可见,在此生长旺季为毛竹提供及时灌水可能会增加其笋产量。
早期的试验表明,没有其它限制因素时,植物叶片光合速率与气孔导度密切相关[22]。另有研究强调了气孔导度对木材产量的影响[17]。较低的气孔导度通过限制二氧化碳扩散到叶片而限制叶片同化速率。一般情况下,植物由于具有较高的气孔导度和较大的同化速率,在最佳生长条件下生长较快,但它们通常具有较低的水分利用效率[17]。本研究发现,在不同生长期,毛竹冠层叶片平均净光合速率始终随着气孔导度增加而增加。这意味着生产实践中我们可根据竹林年龄结构组成对毛竹林分密度进行合理调整,获得更好的冠层结构和冠长,以更好捕获光照,提高叶片光合作用效率,进一步提高林分生产力。
Wen 等[11]发现毛竹个体碳固定峰值一般在9~11月,因此延缓竹子钩梢至12月,在减少雪压的同时可以增加竹子碳汇能力。然而,我们的调查数据显示,12月初毛竹个体叶片仍具有较高的碳吸收和水分利用效率。
迄今为止,评估气孔行为对光合碳增益影响的研究大多数主要集中在影响光合作用的气孔导度的稳态测量方面。然而,这些研究没有考虑到野外现场动态情况,其野外光波动水平是影响气孔行为主要变化的环境因素之一。在本研究中,我们试图通过水分利用效率的季节变化和梯度变化来探讨毛竹叶片气孔行为对不同冠层梯度内光环境变化的应对能力,旨在确定和探讨不同生长季节不同年龄个体毛竹叶片的气孔行为变化及其对冠层高度的响应特征,为在不影响同化速率的情况下进一步提高水分利用效率奠定理论基础。
3.2 冠层高度对毛竹个体叶片光合色素含量的影响
叶片中光合色素含量是反映植物光合能力的一个重要参数,环境因子的改变会引起光合色素含量的变化,进而引起光合性能的改变。光合色素尤其是叶绿素,在植物的光合作用中对光能的吸收、传递和转化起着极为重要的作用。叶绿素含量的多少在一定程度上影响植物的光合速率。不少学者研究了竹类植物叶绿素含量的动态变化,并发现其与光合作用的变化规律具有密切的相关性[23-24]。受太阳活动影响,树冠不同部位叶片接受到的光照存在差异,使得叶绿素含量等叶片生理特性发生改变。何春霞等[25]研究发现,树冠受光最弱的东向方位叶片叶绿素含量最高。本研究发现,不同林龄毛竹个体叶片叶绿素含量随冠层高度的垂直变化特征亦存在差异,且冠层差异受生长季节影响。在孕笋行鞭期,不同林龄毛竹叶片叶绿素a、叶绿素b、叶绿素(a+b)含量均随着冠层自上而下呈现逐渐增加的垂直变化趋势,与何春霞等[25]研究结论一致,反映出不同冠层部位毛竹叶片捕获光能的能力呈现冠层上部低于冠层下部的趋势。而在出笋期,3年生个体其含量随着冠层高度下降而增加,2年生个体反之,表明该季节毛竹叶片叶绿素含量随着冠层高度垂直变化受竹龄影响。研究发现叶绿素a/b值基本都随着冠层高度下降而降低,说明不同生长季节不同林龄毛竹个体利用弱光能力均以冠层上部叶片较强。可见,毛竹不同年龄个体在不同生长季节可通过调节不同冠层部位叶片叶绿素含量来提高叶片捕获光能的能力,进而有利于各生长季节光合作用的正常、高效进行,促进干物质的积累。
3.3 冠层高度对毛竹个体叶片比叶重和叶氮含量的影响
比叶重(LMA)是重要的叶片碳经济性状,显著地影响着叶片光合能力。多数情况下,LMA随光强的增加而增大[25]。与荫蔽环境下叶片相比,处于较高光照条件下的叶片具有较大的比叶重和单位面积光合能力[26]。随林冠层密度增加,受上部叶片影响,下部叶片变得更加的荫蔽,植株表现出适应性反应,即分配较少干物质和较小的光合能力给下部叶片,相对较多干物质和光合能力分配给光照条件比较好的上部叶片[27]。何春霞等[25]研究发现,LMA大小与受光强大小一致,即南向方位叶片LMA最大,而东向方位叶片LMA最小。但本研究发现毛竹冠层叶片LMA大小随冠层高度的变化受生长季节影响,即出笋期冠层上部叶片LMA小,而孕笋行鞭期冠层上部叶片LMA则高于下部的。
另外,氮素是植物生长的主要限制因子之一。大量研究表明,叶氮含量(Nmass)的高低直接决定着叶片光合能力高低[28]。冠层叶片氮含量的垂直梯度分布是植物冠层重要、普遍特征之一。单位面积叶片氮素的垂直分布梯度变化,被看作是碳同化过程中植物优化氮素利用的可塑性反应。目前对毛竹冠层高度对叶片氮素分配影响研究报道较少。本研究发现,无论生长季节,不同林龄毛竹个体叶片氮素含量随冠层高度变化与比叶重梯度变化趋势一致;且不同林龄毛竹个体叶片氮素含量和比叶重随冠层高度垂直变化趋势基本与叶片净光合速率日均值的冠层梯度变化趋势较为一致。上述结果说明毛竹不同冠层部位叶片通过改变形态、氮素含量来适应不同生长季节生长环境的变化,以便充分利用光能提高光合能力。
综上所述,毛竹冠层高度对叶片的光合生理特性的影响有显著差异,且受生长季节的影响。冠层顶部叶片在出笋期受高温、强光照的环境影响,其叶片Tr减慢,但其WUE增加。毛竹出笋期冠层下部叶片比叶重、叶氮素含量、叶绿素含量均较高,因此其冠层下部光合能力也较高;而在孕笋行鞭期,毛竹冠层下部叶片在比叶重、叶氮素含量较低的情况下,则通过增加下部叶片叶绿素含量来保持该季节光合能力。这些差异在一定程度上是毛竹对外界变化环境适应的结果。迄今为止,对于毛竹关键生长发育期间冠层水平叶片光合作用的研究报道较少。因此,今后有待加强毛竹冠层内叶片生理特性指标梯度分布及其对毛竹冠层光合能力和生产力提升的潜在影响研究。
[1] SONG X Z, ZHOU G M, JANG H,etal. Carbon sequestration by Chinese bamboo forests and their ecological benefits: assessment of potential, problems, and future challenges[J].EnvironmentalReviews, 2011,19(NA): 418-428.
[2] WANG B, WEI W J, LIU C J,etal. Biomass and carbon stock in Moso bamboo forests in subtropical China: characteristics and implications[J].JournalofTropicalForestScience, 2013,25(1): 137-148.
[3] LI P H, ZHOU G M, DU H Q,etal. Current and potential carbon stocks in Moso bamboo forests in China[J].JournalofEnvironmentalManagement, 2015, 156: 89-96.
[4] SHI J M, GUO Q R & YANG G Y. Study on the photosynthetic dynamic variation ofPhyllostachyspubescens[J].ForestResearch, 2005,18: 551-555.
[5] 周国模,姜培坤.毛竹林的碳密度和碳贮量及其空间分布[J].林业科学,2004,40(6):51-55.
ZHOU G M , JIANG P K. Density, storage and spatial distribution of carbon inPhyllostachyspubescensforest[J],ScientiaSilvaeSinicae, 2004,40(6): 20-24.
[6] 周国模,吴家森,姜培坤.不同管理模式对毛竹林碳储量的影响[J].北京林业大学学报,2006,28(6):51-55.
ZHOU G M, WU J S , JIANG P K. Effects of different management models on carbon storage inPhyllostachyspubescensforests[J].JournalofBeijingForestryUniversity, 2006,28(6):51-55.
[7] ZHOU B Z, LI Z C, WANG X M,etal. Impact of the 2008 ice storm on moso bamboo plantations in southeast China[J].JournalofGeophysicalResearch:Biogeosciences, 2011,116, doi:10.1029/2009JG001234
[8] ISAGI Y. Ecological study of bamboo communities: a resource-based approach[M]. Bull. Fac. Int. Arts Sci. Hiroshima Univ. 1995,IV 21: 235-238 (in Japanese).
[9] LIN Y M, PENG Z Q , LIN P. Dynamics of leaf mass, leaf area and element retranslocation efficiency during leaf senescence inPhyllostachyspubescens[J].ActaBotanicaSinica, 2004,46: 1 316-1 323.
[10] CAO Y H, ZHOU B Z, WANG X M,etal. The Photosynthetic characteristics of Moso bamboo (Phyllostachyspubescens) for different canopy leaves[J].AdvancedMaterialsResearch, 2013,726-731: 4 274-4 279.
[11] WEN G S, ZHANG L Y, ZHANG R M, . Temporal and spatial dynamics of carbon fixation by Moso bamboo (Phyllostachyspubescens) in subtropical China[J].BotanicalReview, 2011,77: 271-277.
[12] 黄启民,杨迪蝶,高爱新,等.不同条件下毛竹光合作用的研究[J].竹类研究,1989,8(2):8-16.
HUANG Q M, YANG D J , GAO A X,etal. Study on the photosynthetic ofPhyllostachyspubescensunder different conditions[J].BambooResearch, 1989,8: 8-16.
[13] 李合生.植物生理生化实验原理和技术(面向21世纪课程教材)[M]. 北京:高等教育出版社, 2000.
[14] 林琼影,陈建新,杨淑贞,等. 毛竹气体交换特征[J].浙江林学院学报,2008,25(4):522-526.
LIN Q Y, CHEN J X, YANG S Z,etal. Gas exchange withPhyllostachyspubescenson Mount Tianmu, Zhejiang Province[J].JournalofZhejiangForestryCollege, 2008,25(4): 522-526.
[15] KEENAN T F, HOLLINGER D Y, BOHRER G,etal. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise[J].Nature, 2013,499: 324-327.
[16] KLEIN T, SHPRINGER I, FIKLER B,etal. Relationships between stomatal regulation, water-use, and water-use efficiency of two coexisting key Mediterranean tree species[J].ForestEcologyandManagement, 2013,302: 34-42.
[17] LAWSON T, BLATT M R. Stomatal size, speed, and responsiveness impact on photosynthesis and water use efficiency[J].PlantPhysiology, 2014,164: 1 556-1 570.
[18] KOMATSU H, ONOZAWA Y K, KUME T,etal. Canopy conductance for a Moso bamboo (Phyllostachyspubescens) forest in western Japan[J].AgriculturalandForestMeteorology, 2012,156: 111-120.
[19] FU M Y, FANG M Y, XIE J Z,etal. Nutrient cycling in bamboo stands. Ⅰ. Leaf litter and its decomposition in purePhyllostachyspubescensstands[J].ForestResearch, 1989,2(3): 207-213.
[20] GE X G, ZHOU B Z, TANG Y L. Litter production and nutrient dynamic on a Moso bamboo plantation following an extreme disturbance of 2008 ice storm[J].AdvancesinMeteorology, 2014,8: 1-10.
[21] BERNACCHI C J, KIMBALL B A, QUARLES D R,etal. Decreases in stomatal conductance of soybean under open-air elevation of CO2are closely coupled with decreases in ecosystem evapotranspiration[J].PlantPhysiology, 2007,143: 134-144.
[22] WONG S C, COWAN I R , FARQUHAR G D. Stomatal conductance correlates with photosynthetic capacity[J].Nature, 1979,282: 424-426.
[23] 宋艳冬,金爱武,金晓春,等.施肥对毛竹叶片光合生理的影响[J].浙江林学院学报,2010,27(3): 334-339.
SONG Y D, JIN A W, JIN X C,etal. Physiology of leaf photosynthesis with fertilization inPhyllostachyspubescens[J],JournalofZhejiangForestryCollege, 2010,27(3): 334-339.
[24] 李 凯,余学军,董大川,等. 绿竹滩涂地造林的快速叶绿素荧光动力学分析[J]. 竹子研究汇刊,2012,31(4): 21-27.
LI K, YU X J, DONG D C,etal. An analysis of fast chlorophyll fluorescence induction dynamics ofBambusaoldhamion intertidal zone[J],JournalofBambooResearch, 2012,31(4): 21-27.
[25] 何春霞,李吉跃,张燕香,等.5种绿化树种叶片比叶重、光合色素含量和δ13C的开度与方位差异[J].植物生态学报,2010,34(2): 134-143.
HE C X, LI J Y , ZHANG Y X,etal. Differences in leaf mass per area, photosynthetic pigments and δ13C by orientation and crown position in five greening tree species [J].ChineseJournalofPlantEcology, 2010,34(2): 134-143.
[26] KULL O. Acclimation of photosynthesis in canopies: models and limitations[J].Oecologia, 2002, 133: 267-279.
[27] BOONMAN A, PRINSEN E, GILMER F,etal. Cytokinin import rate as a signal for photosynthetic acclimation to canopy light gradients[J].PlantPhysiology, 2007, 143: 1 841-1 852.
[28] FIELD C , MOONEY H A. Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a California shrub[J].Oecologia. 1983,56: 348-355.
(编辑:裴阿卫)
Effects of Canopy Height on Photosynthetic Physiology Characteristics ofPhyllostachyspubescensLeaves
CAO Yonghui1, ZHOU Benzhi1*, WANG Xiaoming1, GU Lianhong2
(1 Research Institute of Subtropical Forestry, Chinese Academy of Forestry, Qianjiangyuan Forest Ecosystem Research Station, State Forestry Administration, Fuyang, Zhejiang 311400, China; 2 Environmental Sciences Division, Oak Ridge National Laboratory, Oak Ridge, TN, USA 37831)
The carbon sequestration ability, water use efficiency (WUE) and other photosynthetic physiology characteristics for different aged ofPhyllostachyspubescensindividuals were analyzed at two canopy layers with a LI-6400 portable photosynthesis system. The results showed that: (1) the daily average net photosynthetic rate (Pn) and the transpiration rate (Tr) ofP.pubescenswere lower for upper canopy leaves than those of lower canopy leaves regardless of bamboo age during the culm elongation period. During the shoot development period, the upper canopy leaves had a higherPnvalue at each time point and a higher daily averagePnthan that of the lower canopy leaves regardless of bamboo age. Meanwhile, theTrvalue for upper canopy leaves of 2 year-old bamboo was higher at each time point than that for lower canopy leaves. The vertical change of stomatal conductance (Gs) for different age groups was consistent with that ofTr, regardless of the growing season. (2) A reduction of photosynthesis at noon was observed at different canopy layers of 3 year-old bamboo and the upper canopy of 2 year-old bamboo during the culm elongation period, while it only occurred in the upper canopy of 2 year-old bamboo during the shoot development period. (3) The daily average ofWUEin the culm elongation period increased along with canopy height regardless of age. While, the daily average ofWUEdecreased with increasing canopy height during the shoot development period for 2 year-old individuals. So, the canopy height had obvious effects on bamboo leaf photosynthesis andWUE, which also impacted by growing seasons. There was negative relationship betweenGsandWUEfor bamboo with all ages and canopy layers. We wanted to explore the variable stomata behavior of moso bamboo at different ages during its important growing seasons and in order to improvingWUEand lay the foundation for further mechanistic research on water use strategy for bamboo. (4) The chlorophyll a/b values decreased with the decrease of canopy height in different growing seasons. While the chlorophyll content of all ages of bamboo was increased with the decrease of canopy height. The vertical variation of leaf nitrogen content and leaf mass per area within canopy was consistent with that of leaf net photosynthetic rate regardless of the growing seasons. The results indicated that the leaf morphology and nitrogen content in the different canopies could change with the growth environment in different growing seasons, and make full use of light energy to improve the photosynthetic capacity. Our data could provide a scientific basis for developing sustainable cultivation technologies and enhancing moso bamboo timber production and carbon sequestration.
Phyllostachyspubescens; photosynthesis; transpiration; water use efficiency; canopy height; seasonal variation; diurnal variation.
1000-4025(2016)11-2256-11
10.7606/j.issn.1000-4025.2016.11.2256
2016-05-09;修改稿收到日期:2016-11-16
浙江省自然科学基金项目(LY13C160002);国家自然科学基金(31600503);国家林业局948项目(2014-4-57);中央级公益性科研院所基本科研业务费专项(RISF2013002);中国林业科学研究院基本科研业务费专项资金项目——海内外杰出学者讲习计划(CAFYBB2011007)
曹永慧(1977-),女,博士,助理研究员,主要从事植物生理生态学研究。E-mail:fjcyh77@sina.com
*通信作者:周本智,研究员,博士生导师,主要从事森林生态结构与功能研究。E-mail:benzhi_zhou@126.com
Q945.78
A
