塔里木河下游胡杨生物量及其空间分布特征
2017-01-04王希义徐海量潘存德凌红波苑塏烨
王希义,徐海量,潘存德,凌红波,苑塏烨
(1 新疆农业大学,乌鲁木齐 830052;2 中国科学院新疆生态与地理研究所,乌鲁木齐 830011)
塔里木河下游胡杨生物量及其空间分布特征
王希义1,徐海量2*,潘存德1,凌红波2,苑塏烨1
(1 新疆农业大学,乌鲁木齐 830052;2 中国科学院新疆生态与地理研究所,乌鲁木齐 830011)
该研究采用野外样地调查法,以塔里木河下游地区的胡杨为研究对象,以胡杨地上生物量与易测因子(胸径、树高)之间的关系模型为基础,依据监测样带胡杨状况,对研究区内胡杨生物量分布状况进行分析,并结合区域内胡杨林面积对胡杨总生物量进行了评估。结果表明:(1)胡杨生物量模型B=128.09(D2h)0.6318(R2=0.892),精度较高,并且验证的实测值与模拟值之间差异不显著(P> 0.05),此模型可以用来对胡杨生物量进行评估。(2)随着离河道距离的增加,胡杨生物量呈减少趋势;胡杨生物量主要分布在离河道700 m以内的范围,占总生物量的91.37%。(3)塔里木河下游胡杨单位面积生物量为416.613 kg·hm-2,总生物量为2.474×104t。
胡杨;地上生物量;空间分布;塔里木河下游
森林是陆地生态系统的主体[1],与其他生态系统相比具有更为复杂的层次结构和更长的生命周期,是陆地生物光合产量的主体[2-3]。森林生物量指个体、种群或群落生物质数量的多少,是生态系统特征的重要指标之一,也是森林系统环境质量的综合体现[4]。生物量不仅反映了生态系统生产力水平的高低,也是植被固碳能力的重要标志和评估植被碳收支的重要参数[5-6],其测定在碳循环及气候变化等研究中具有重要意义[7-8]。
自国际生物学计划(IBP)实施以来,生物量一直是生态学研究的重点[9-10]。在国外,一些学者针对森林生物量与生产力进行了研究,建立了生物量与碳循环之间的关系[11-13],并对不同区域的生物量进行了评估[14-15];国内针对热带雨林、亚热带常绿阔叶林、温带针叶林等森林系统的生物量进行了探讨,分析了森林生物量的分布规律与变化特征[16-19]。然而这些研究大都集中于东部湿润区或是热带亚热带地区,有关干旱区森林生物量的研究较为少见。
胡杨是塔里木河下游的植被建群种,对于区域内环境保护和植被恢复程度具有最直接的指示作用[20]。针对塔里木河下游胡杨的研究主要集中于胡杨与水资源的关系[21-22]、胡杨生态分布[23-25]与胡杨生理特征[26-28]等方面,而有关生物量模拟评估的研究较少。本研究通过查询生物量模拟方法,结合调查资料,探讨了塔里木河下游胡杨生物量的空间分布特征,并对胡杨总生物量进行了评估,以便于人们对胡杨在生活与生产中的作用有一个恰当认识,为塔里木河下游植被保护和恢复以及准确估算区域胡杨生物量和碳储量提供科学依据。
1 研究区概况
塔里木河下游是指从大西海子水库至台特玛湖河段(图1)。本河段位于库鲁克沙漠与塔克拉玛干沙漠之间,区域内的荒漠河岸林形成了南北长约400 km的绿色走廊。塔里木河下游属温带干旱荒漠气候,年降水量仅为20~50 mm,而潜在蒸发量却高达2 500~3 000 mm[29]。
由于人类不合理的水土资源开发和利用,使下游的河道曾经长期断流,生态严重退化,植被长势极度衰败,区域内建群植物——胡杨基本为过熟林,中幼龄林基本枯死[30]。基于此,中国政府于2000年5月组织实施了以生态恢复和环境保护为目标的塔里木河下游生态输水工程。在实施生态输水的过程中,地下水位不断抬升,地下水的响应宽度不断扩大[31];另外,河道两岸出现了大范围河水漫溢现象,激活了土壤种子库,在漫溢区萌生了大量幼苗[32],增加了生态系统的稳定性,在一定程度上缓解了区域内的生态退化过程。塔里木河下游乔木主要是胡杨(Populuseuphratica),灌木主要包括柽柳(Tamarixchinensis)、黑刺(Lyciumruthenicum)、铃铛刺(Halimodendronhalodendron)等,草本植物包括芦苇(Phragmitescommunis)、鹿角草(GlossogynetenuifoliaCass)、骆驼刺(Alhagisparsifolia)、花花柴(Kareliniacaspica)、盐生草(Halogetonglomeratus)等。
2 资料来源与研究方法
2.1 资料来源
于2016年6月上中旬至7月上旬在塔里木河下游进行胡杨样地调查。选取塔里木河下游英苏、喀尔达依、阿拉干、依干不及麻4个断面。对于每一个监测断面,选择具有代表性、未受人类破坏的地段,沿垂直于河道方向设置一个长2 000 m、宽100 m的样带,然后将此样带平均分成20个100 m×100 m的大样方进行监测。首先统计各个大样方内胡杨的数量,然后对每个大样方内的胡杨进行每木检尺(DBH≥4 cm),记录其胸径、树高、冠幅等,对于DBH<4 cm的幼苗及幼树,记录其数量和高度。
由于以前没有开展胡杨生物量的研究,因此在2016年野外调查时,为了保证实验的合理性与数据的均匀性,并兼顾保护植被稀疏地带的胡杨不再遭受破坏,本研究在胡杨分布最多的英苏和阿拉干断面对胡杨进行取样。在对胡杨的取样过程中,首先选取胡杨分布密集且均匀的地区,一共标记60株幼中龄的、长势良好的标准木,先记录这些胡杨的胸径、树高、冠幅等,然后在地面根基部位锯断伐倒并称重,记录这60株胡杨的地上生物量。
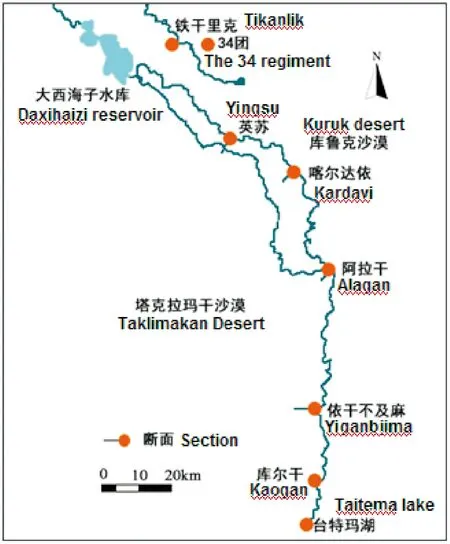
图1 塔里木河下游各监测断面状况Fig.1 Distribution of monitoring sections in the lower reaches of Tarim River
2.2 研究方法
本研究的关键方法是构建胡杨生物量估算模型。本文采用唐守正[33]建立的生物量回归模型计算胡杨单株生物量。
B=a(D2h)b(D为胸径,h为树高)
(1)
首先用30株伐倒胡杨(编号1,2,……,30)的相关数据对a与b进行参数拟合,计算出另外30株(编号31,32,……,60)胡杨生物量,并用T检验方法,通过计算P值分析模拟值与实测值之间差异的显著性,若P< 0.05,则模拟值与实测值差异显著;若P> 0.05,则模拟值与实测值之间不显著。然后计算样方内胡杨总生物量,进而推算胡杨单位面积生物量,最后结合胡杨林总面积计算总生物量。
本研究分析和作图软件用SPSS16.0与EXCEL。
3 结果与分析
3.1 胡杨生物量模型建立与变量统计
3.1.1 胡杨生物量模型构建 利用伐倒30株胡杨建立回归模型(方程(2)),且拟合度达到显著水平。
B= 128.09(D2h)0.6318(R2= 0.892;F= 285.35)
(2)
之后利用剩余30株胡杨特征数据进行验证,对实测值与模拟值进行对比分析(图2)。由图2可知,实测值与模拟值之间没有显著差异(P> 0.05),拟合方程合理。因此可以用方程(2)计算胡杨的生物量。
3.1.2 胡杨地上变量统计分析 结合胡杨生物量估算模型(1),重点对所调查胡杨的胸径和树高进行统计分析(表1)。由表1可知,胡杨胸径的最小值和最大值都出现在英苏断面。由于胡杨的胸径反映出了树龄的大小[34],因此英苏断面的胡杨树龄宽度最大。依干不及麻断面胡杨胸径平均值最低,高度平均值也显著低于其他3个断面,这导致胡杨单株平均生物量显著低于其他断面。另外,胡杨单株生物量最小值和最大值均出现在英苏断面,表明在英苏断面胡杨龄级跨度大;依干不及麻断面胡杨生物量最小值与最大值差异最小,表明该断面胡杨龄级跨度小。
3.2 胡杨地上生物量空间分布特征
3.2.1 各断面胡杨单位面积生物量横向分布特征 生物量横向分布特征即为随着离河距离的增加,生物量的变化趋势。本文将4条样带内不同离河距离胡杨生物量特征作图讨论(图3)。
由图3可知,在离河道700 m以内胡杨分布较多,生物量总量较大,占总生物量的91.37%;700 m以外生物量分布较少。这主要是由于随着离河距离的增加,河水漫溢作用减弱,地下水补给量逐渐减少,水位降低,逐渐邻近甚至低于胡杨生长的胁迫水位,导致胡杨生长萎靡,生物量降低。对于各个断面而言,近河岸胡杨生物量高于离河较远的地区,最为明显的是英苏断面,其次是依干不及麻断面。喀尔达依断面的胡杨生物量在离河900 m附近出现一个小高峰,阿拉干断面在离河600~700 m处胡杨的生物量也相对较高,造成这种现象的原因也许与微地形或是样带的随机设置有关,具体原因需要进一步探讨。

图2 胡杨实测值与模拟值对比Fig.2 Comparison of measured values and simulation values of P. euphratica

表1 胡杨地上因子统计

图3 各断面不同离河距离上胡杨单位面积生物量分布特征Fig.3 The distribution characteristics of P. euphratica biomass per unit area in different distances from river to each section

表2 各监测样带内胡杨单位面积生物量
3.2.2 胡杨生物量纵向分布特征 生物量纵向分布规律即在沿河道方向上从上游到下游的变化特征。本文将4条样带的胡杨总生物量进行了对比分析(图4)。
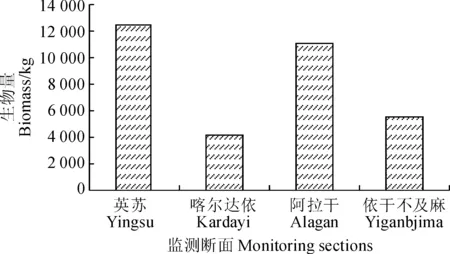
图4 自上游到下游胡杨生物量特征Fig.4 The distribution characteristics of P. euphratica biomass from upstream to downstream
各样带中,英苏的胡杨生物量最高,阿拉干次之,喀尔达依和依干不及麻的胡杨生物量最小。因为英苏离放水源最近,断面内河水对地下水和土壤水的转化量较大,可以较好地满足胡杨的生长需求,因此胡杨的数量较多,长势较好;另外,阿拉干断面位于其文阔尔河与老塔里木河的交汇处,这使得阿拉干断面水源也很充足,胡杨生物量也较高;在喀尔达依与依干不及麻断面,河道的过水量较少,导致河水对地下水和土壤水的转化量较低,断面内的水分条件限制了胡杨的生长,使得胡杨的数量少,长势差,生物量较低。
3.3 塔里木河下游胡杨生物量估测
3.3.1 胡杨单位面积生物量 由于样带设置的随机性,样方内所取胡杨特征也具有随机性。在本研究中,每个样带设置了20个大样方,但并非所有的样方内都有胡杨。例如阿拉干断面,在样带内,离河道1 100 m以内有胡杨,远处没有胡杨;在样带之外离河道1 100 m以外依然有胡杨分布,直至到2 000 m以外的区域都有胡杨。因此本研究中,计算样带内胡杨平均生物量(表2)依然使用整条样带的面积,而非仅有胡杨分布区域的面积。
由表2可知,英苏断面胡杨单位面积生物量最高,阿拉干次之,喀尔达依断面最小,这与图4相对应。另外,塔里木河下游胡杨单位面积平均生物量取4个断面单位面积生物量的平均值,即416.613 kg/hm2。
3.3.2 区域内胡杨总生物量 本研究利用计算得出的胡杨单位面积生物量,然后结合塔里木河下游胡杨林总面积,可得塔里木河下游胡杨总生物量。在本研究中,根据2013年的调查结果,塔里木河下游林地总面积约为593.8 km2[35],即59 380 hm2,得胡杨总生物量为2.474×107kg(416.613 kg/hm2×59 380 hm2)。
4 讨 论
乔木生物量估测是森林生物量评估中重要组成部分,其模型中变量的选择有多种形式,其中最常用的是以胸径和高度作为模型的变量;另外董道瑞[36]建立了冠幅与生物量之间的关系模型,曾慧卿[37]以冠幅和高度之积为自变量拟合了乔木生物量估测模型,精度均较高,都在0.8左右。本研究中,以胸径平方与树高之积作为变量,与胡杨的地上生物量建立了关系模型,精度高于冠幅与生物量间关系模型,并且利用此模型计算的估测值接近实测值,这与Lyon[38]揭示的体积与生物量的关系比面积与生物量关系更为紧密的结论相一致。在实施生态输水以前,塔里木河下游各个断面胡杨都十分衰败,自开展生态输水以来,胡杨大面积复苏[39]。英苏断面离放水地点较近,地下水位较高,并且河水漫溢作用较强,大大促进了各龄级胡杨的生长发育,胡杨平均生物量也最高。另外,塔里木河下游生态输水是其文阔尔河和老塔里木河双河道输水,两个河道在阿拉干断面汇合,使得阿拉干断面水量也较多,胡杨长势较好,平均生物量也较高,这与本研究成果相吻合。依干不及麻断面胡杨龄级跨度小,长势较差,这是由于依干不及麻断面离水源较远,地下水位低,漫溢作用弱,抑制了胡杨的生长发育[40]。
植被的生长发育与生态环境之间存在着一定的关系,生态环境的变化影响着植被的组成、结构、形成以及动态分布等,而植被的变化又会反过来作用于生态环境[41]。在干旱区,植被地上生物量与地下水间的关系十分显著[42-43]。在离河道近处,地下水埋深较浅,植物根系与地下水的接触面积相对较大,水分较为充足,漫溢作用较强,植被长势较好,单位面积生物量较大;随着与河距离的增加,地下水埋深加大,植物根系与地下水的接触面积越来越小,水分条件逐渐变差,植被长势逐渐衰弱,单位面积生物量减小,这正与本研究的结果相符合。本研究所布设的样带内,胡杨大都分布在离河道1 000 m的范围内,1 000 m以外胡杨分布很少。在离河道700 m以内胡杨分布较多,生物量总量较大,700 m以外生物量分布较少。另外,具体到每一个监测样带,胡杨生物量分布没有明显规律,如喀尔达依断面胡杨生物量在离河900 m出现一个小高峰,阿拉干断面在离河600~700 m处胡杨的生物量也较高,这也许与胡杨在荒漠环境中呈斑块状分布[44]有关,或是受到微地形或样带随机设置的影响,具体原因需要进一步探讨。
塔里木河下游荒漠河岸林衰败、老化与退化现象严重[45],胡杨平均生物量仅为416.613 kg·hm-2,远远低于中国东部地区,如贡嘎山区[46]森林单位面积生物量最高达568 t·hm-2,是塔里木河下游的十几倍;又如浙江林分单位面积生物量虽远低于全国平均水平[47],但是依然高于塔里木河下游。这一方面说明塔里木河下游胡杨生物量与生产力都很低,另一个方面也反映了胡杨恢复还有很大发展潜力。但是,在所设立的样带内,不仅仅分布有胡杨,还有柽柳、骆驼刺、花花柴、鹿角草等灌木和草本植物,胡杨的生长发育与这些林下植被有密切关系,如胡杨根系提水作用促进灌草的生长[48],而灌草的物种多样性维持了生态系统的稳定[49]等,以后应加强此方面的研究。另外,就本研究的取样方式而言,也存在着一些误差和不确定性,如取样选取的是幼中龄胡杨并建立模型,用此模型模拟老龄胡杨生物量时就存在误差。本研究采用每木检尺、实地采伐测量的方式,从理论上来讲,对于所取的胡杨样品实测的各项数据应该没有误差。但是在实际的操作过程中,由于受到人为、仪器等因素的影响,计算出的结果也存在着一定误差。但是与机械角规样地实测法、样圆实测法、角规典型选样调查法等方法相比,精度是最高的[50],结果可信。因此,在以后的研究中应深入分析模型误差的范围、来源及影响因素,提高建模过程中各个环节的精确度。
综上所述,本研究构建了胡杨生物量估测模型:B= 128.09(D2h)0.6318(R2= 0.892;F= 285.35),且实测值与模拟值之间没有显著差异(P> 0.05);英苏断面胡杨树龄跨度大,各龄级胡杨分布多,依干不及麻断面胡杨树龄跨度小。胡杨大都分布在离河道1 000 m的范围内,1 000 m以外胡杨分布很少;在离河道700 m以内胡杨生物量占总生物量的91.37%;700 m以外生物量较少。根据调查结果,塔里木河下游胡杨单位面积生物量约为416.613 kg·hm2,胡杨总生物量约为2.474×104t。
[1] LYON L J. Estimating twig production of serviceberry from crown volumes[J].JournalofWildlifeManagement, 1968,32(1):115-119.
[2] CONSTANZA R, RUDOLFDE G. The value of the world ecosystem service and natural capital [J].Nature, 1997,387:253-260.
[3] NELSON R, KRABILL W, TONELLI J. Estimating forest biomass and volume using airbone laser data[J].RemoteSensingofEnvironment, 1988, 24(88):247-267.
[4] 张远彬.CO2浓度升高对红桦幼苗生理与生长的影响[D].北京:中国科学院大学,2007.
[5] 奥得姆.生态学基础[M].北京:人民教育出版社,1971.
[6] 王 昆.基于Quickbird和极化雷达数据的森林生物量估测方法研究[D].北京:北京林业大学,2013.
[7] 杨永恬.基于多源遥感数据的森林蓄积量估测方法研究[D].北京:中国林业科学研究院,2010.
[8] 韦龙斌.九万山自然保护区森林生物量及碳贮量研究[D].长沙:中南林业科技大学,2015.
[9] 翟晓江.基于遥感的陕北黄龙山森林生物量模型[D].陕西杨陵:西北农林科技大学,2013.
[10] KARL M G, HEITSCHMIDT R K, HAFERKAMP M R. Vegetation biomass dynamics and patterns of sexual reproduction in a Northern Mixed-Grass Prairie[J].AmericanMidlandNaturalist, 2013, 141(Apr 1999):227-237.
[11] MUNEEPEERAKUL R, RINALDO A, RODRIGUEZ I I. Effects of river flow scaling properties on riparian width and vegetation biomass[J].WaterResourcesResearch, 2007, 43(12):55-60.
[12] RYAN J G, FYFE C T, MCALPINE C A. Biomass retention and carbon stocks in integrated vegetation bands: a case study of mixed-age brigalow-eucalypt woodland in southern Queensland, Australia[J].RangelandJournal, 2015, 37(3):261-271.
[13] SIMPSON L G, BOTKIN D B. Vegetation, the Global Carbon Cycle, and Global Measures[C].1992:413-425.
[14] ADAMS J M, PIOVESAN G. Uncertainties in the role of land vegetation in the carbon cycle[J].Chemosphere, 2002, 49(8):805-819.
[15] SINGH T P, DAS S. Predictive analysis for vegetation biomass assessment in Western Ghat Region (WG) using geospatial techniques[J].JournaloftheIndianSocietyofRemoteSensing, 2014, 42(3):549-557.
[16] ISABIRYE M, VERBIST B, MAGUNDA M K,etal. Tree density and biomass assessment in agricultural systems around Lake Victoria, Uganda[J].AfricanJournalofEcology, 2008, 46(Supplement s1):59-65.
[17] 杨存建,刘纪远,黄 河,等.热带森林植被生物量与遥感地学数据之间的相关性分析[J].地理研究,2005,24(3):473-479.
YANG C J, LIU J Y, HUANG H,etal. Correlation analysis of the biomass of the tropical forest vegetation, meteorological data and topographical data[J].GeographicalResearch, 2005,24(3):473-479.
[18] 邢艳秋,王立海.基于森林调查数据的长白山天然林森林生物量相容性模型[J].应用生态学报,2007,18(1):1-8.
XING Y Q, WANG L H. Compatible biomass estimation models of natural forests in Changbai Mountains based on forest inventory[J].ChineseJournalofAppliedEcology, 2007,18(1):1-8.
[19] 范文义,张海玉,于 颖,等.三种森林生物量估测模型的比较分析[J].植物生态学报,2011,35(4):402-410.
FAN W Y, ZHANG H Y, YU Y,etal. Comparison of three models of forest biomass estimation[J].ChineseJournalofPlantEcology, 2011,35(4):402-410.
[20] 徐 婷,曹 林,申 鑫,等.基于机载激光雷达与Landsat 8 OLI数据的亚热带森林生物量估算[J].植物生态学报,2015,39(4):309-321.
XU T, CAO L, SHEN X,etal. Estimates of subtropical forest biomass based on airborne LiDAR and Landsat 8 OLI data[J].ChineseJournalofPlantEcology, 2015,39(4):309-321.
[21] 徐海量,宋郁东,王 强.胡杨生理指标对塔里木河下游生态输水的响应[J].环境科学研究,2003,16(4):24-27.
XU H L, SONG Y D, WANG Q. Response of Physiological Indexes ofPopuluseuphraticato Ecological Water Transport in the Lower Reaches of the Tarim River[J].ResearchofEnvironmentalSciences, 2003,16(4):24-27.
[22] 邓潮洲,张希明,李 利,等.河道输水对塔里木河下游胡杨生长状况的影响[J].中国沙漠,2010,30(2):312-318.
DENG C Z, ZHANG X M, LI L,etal. Effects of replenishing water into river on the growth ofPopuluseuphraticain Lower Reaches of Tarim River[J].JournalofDesertResearch,2010,30(2):312-318.
[23] 涂文霞,叶 茂,徐海量,等.塔里木河不同河段胡杨径向生长及径流的影响差异[J].干旱区研究,2014,31(3):508-515.
TU W X, YE M, XU H L,etal. Radial growth ofPopuluseuphraticaand the effect of runoff in the Tarim River Basin[J].AridZoneResearch, 2014,31(3):508-515.
[24] 蔡富艳,玉米提·哈力克,艾尔肯·艾白不拉,等.塔里木河下游阿拉干断面胡杨树高的分布和密度[J].生态环境学报,2008,17(3):1 086-1 090.
CAI F Y, UMUT H, ARKIN A,etal. Tree height distribution and density ofPopuluseuphraticaforest along the Argan section in the lower reaches of Tarim River[J].EcologyandEnvironment, 2008,17(3):1 086-1 090.
[25] 陈亚宁,陈亚鹏,李卫红,等.塔里木河下游胡杨脯氨酸累积对地下水位变化的响应[J].科学通报,2003,48(9):958-961.
CHEN Y N, CHEN Y P, LI W H,etal. Response of proline accumulation ofPopuluseuphraticato the changes of groundwater level in the Lower Reaches of Tarim River[J].ChineseScienceBulletin, 2003,48(9):958-961.
[26] 张绘芳,李 霞.塔里木河下游胡杨种群空间分布格局分析[J].西北植物学报,2006,26(10):2 125-2 130.
ZHANG H F, LI X. Spatial distribution pattern ofPopuluseuphraticapopulations in the Lower Reaches of Tarim River[J].ActaBotanicaBoreali-OccidentaliaSinica, 2006,26(10):2 125-2 130.
[27] 安红燕,叶 茂,徐海量,等.塔里木河下游胡杨径向生长量对生态输水的响应[J].中国沙漠,2011,31(4):957-962.
AN H Y, YE M, XU H L,etal. Response of radial increment ofPopuluseuphraticato ecological water conveyance in Lower Reaches of the Tarim River[J].JournalofDesertResearch, 2011,31(4):957-962.
[28] 朱绪超,袁国富,邵明安,等.塔里木河下游河岸带植被的空间结构特征[J].植物生态学报,2015,39(11):1 053-1 061.
ZHU X C, YUAN G F, SHAO M A,etal. Spatial pattern of riparian vegetation in desert of the lower Tarim River basin[J].ChineseJournalofPlantEcology, 2015,39(11):1 053-1 061.
[29] 苏里坦,李玉生,艾尔肯·艾比布拉,等.塔里木河下游河岸胡杨(Populuseuphratica)林耗水过程模拟[J].中国沙漠,2015,35(2):380-384.
SULITAN, LI Y S, AERKEN A,etal. Simulation of water consumption ofPopulouseuphraticaforests in the Lower Reaches of the Tarim River[J].JournalofDesertResearch, 2015,35(2):380-384.
[30] 张青青,徐海量,安红燕,等.塔里木河下游河水漫溢后胡杨幼苗分布格局初探[J].西北植物学报,2011,31(11):2 298-2 303.
ZHANG Q Q, XU H L, AN H Y,etal. Distribution patterns ofPopuluseuphraticaseedlings after river overflowing in the Lower Reaches of Tarim River[J].ActaBotanicaBoreali-OccidentaliaSinica, 2011,31(11):2 298-2 303.
[31] 杨鹏年,邓铭江,李 霞,等.塔里木河下游间歇输水下地下水响应宽度——以塔里木河下游英苏断面为例[J].干旱区研究,2008,25(3):331-335.
YANG P N, DENG M J, LI X,etal. Respond width of groundwater level after conveying stream water to the Lower Reaches of the Tarim River, Xinjiang——A case study along the Yengisu Section in the Lower Reaches of Tarim River[J].AridZoneResearch, 2008,25(3):331-335.
[32] 徐海量,宋郁东,王 强,等.塔里木河中下游地区不同地下水位对植被的影响[J].植物生态学报,2004,28(3):400-405.
XU H L, SONG Y D, WANG Q,etal. The effect of groundwater level on vegetation in the Middle and Lower Reaches of The Tarim River, Xinjiang, China[J].ChineseJournalofPlantEcology, 2004,28(3):400-405.
[33] 赵振勇,王让会,孙洪波,等.塔里木河下游输水廊道植被恢复的生态学评价[J].科学通报,2006,51(S1):31-35.
ZHAO Z Y, WANG R H, SUN H B,etal. Ecological evaluation of vegetation restoration in water delivery corridor in The Lower Reaches of Tarim River[J].ChineseScienceBulletin, 2006,51(S1):31-35.
[34] 唐守正,张会儒,胥 辉.相容性生物量模型的建立及其估计方法研究[J].林业科学,2000,36(S1):19-27.
TANG S Z, ZHANG H R, XU H. Study on establish and estimate method of Compatible Biomass Model[J].ScientiaSilvaeSinicae, 2000,36(S1):19-27.
[35] 顾亚亚,张世卿,李先勇,等.濒危物种胡杨胸径与树龄关系研究[J].塔里木大学学报,2013,25(2):66-69.
GU Y Y, ZHANG S Q, LI X Y,etal. Relationship between diameter at breast height and age of endangered speciesPopulusEuphraticaOliv[J].JournalofTarimUniversity, 2013,25(2):66-69.
[36] 白 元,徐海量,刘新华,等.塔里木河干流景观格局梯度分析[J].干旱区研究,2013,30(6):1 064-1 072.
BAI Y, XU H L, LIU X H,etal. Gradient change of landscape pattern in the Mainstream Area of the Tarim River[J].AridZoneResearch, 2013,30(6):1 064-1 072.
[37] 董道瑞,李 霞,万红梅,等.塔里木河下游柽柳灌丛地上生物量估测[J].西北植物学报,2012,32(2):384-390.
DONG D R, LI X, WAN H M,etal. Aboveground biomass estimation ofTamarixramosissimaShrub in the Lower Reaches of Tarim River[J].ActaBotanicaBoreali-OccidentaliaSinica, 2012,32(2):384-390.
[38] 赵万羽,陈亚宁,周洪华,等.塔里木河下游生态输水后衰败胡杨林更新能力与条件分析[J].中国沙漠,2009,29(1):108-113.
ZHAO W Y, CHEN Y N, ZHOU H H,etal. Reproductive ability and relative environment factors of degradedPopuluseuphraticaforest in ecological water delivery project at Lower Reaches of Tarim River[J].JournalofDesertResearch, 2009,29(1):108-113.
[39] 曾慧卿,刘琪璟,马泽清,等.基于冠幅及植株高度的檵木生物量回归模型[J].南京林业大学学报(自然科学版),2006,30(4):101-104.
ZENG H Q, LIU Q J, MA Z Q,etal. The Regression Model ofLoropetalumchinensebiomass based on canopy diameter and plant height[J].JournalofNanjingForestryUniversity(Natural Sciences Edition), 2006,30(4):101-104.
[40] 叶 茂,徐海量,龚君君,等.不同胸径胡杨径向生长的合理生态水位研究[J].地理科学,2011,31(2):172-177.
YE M, XU H L, GONG J J,etal. Rational ecological groundwater level ofPopuluseuphraticawith different diameter in Lower Reaches of Tarim River[J].ScientiaGeographicaSinica, 2011,31(2):172-177.
[41] 田 雨.森林群落的水分自营作用及其定量模式研究[D].北京:中国科学院研究生院,2012.
[42] 赵文智,常学礼,李启森,等.荒漠绿洲区芦苇种群构件生物量与地下水埋深关系[J].生态学报,2003,23(6):1 138-1 146.
ZHAO W Z, CHANG X L, LI Q S,etal. Relationship between structural component biomass ofreedpopulation and ground water depth in desert oasis[J].ActaEcologicaSinica, 2003,23(6):1 138-1 146.
[43] 王希义,徐海量,潘存德,等.塔里木河下游地下水埋深对草本植物地上特征的影响[J].生态学杂志,2015,34(11):3 057-3 064.
WANG X Y, XU H L, PAN C D,etal. The influence of groundwater depth on aboveground herbaceous characteristics in the lower reaches of Tarim River[J].ChineseJournalofEcology, 2015,34(11):3 057-3 064.
[44] 白 元.漫溢干扰对胡杨种群空间分布格局的影响[D].北京:中国科学院大学,2013.
[45] 徐海量,李吉玫,张占江,等.塔里木河下游退化荒漠河岸林地上植被与土壤种子库关系初探[J].中国沙漠,2008,28(4):657-664.
XU H L, LI J M, ZHANG Z J,etal. Study on relation between soil seed bank and standing vegetation along different degradation levels of desert riverside forest in Lower Reaches of Tarim River[J].JournalofDesertResearch, 2008,28(4):657-664.
[46] 罗 辑,杨 忠.贡嘎山森林生物量和生产力的研究[J].植物生态学报,2000,24(2):191-196.
LUO J, YANG Z. A study on the biomass and production of forest on the Gongga Mountain[J].ChineseJournalofPlantEcology, 2000,24(2):191-196.
[47] 张茂震,王广兴,刘安兴.基于森林资源连续清查资料估算的浙江省森林生物量及生产力[J]. 林业科学,2009,45(9):13-17.
ZHANG M Z, WANG G X, LIU A X. Estimation of forest biomass and net primary production for Zhejiang Province based on continuous forest resources inventory[J].ScientiaSilvaeSinicae, 2009,45(9):13-17.
[48] 苏 芮.胡杨的水力提升作用及功能研究[D].北京:中国科学院研究生院,2011.
[49] 江小雷,岳 静,张卫国,等.生物多样性,生态系统功能与时空尺度[J].草业学报,2010,19(1):219-225.
JIANG X L, YUE J, ZHANG W G,etal. Biodiversity, ecosystem functioning and spatio-temporal scales[J].ActaPrataculturaeSinica, 2010,19(1):219-225.
[50] 梁 倍,邸 利,赵传燕,等.祁连山天老池流域灌丛地上生物量空间分布[J].应用生态学报,2014,25(2):367-373.
LIANG B, DI L, ZHAO C Y,etal. Spatial distribution of aboveground biomass of shrubs in Tianlaochi catchment of the Qilian Mountains[J].ChineseJournalofAppliedEcology, 2014,25(2):367-373.
(编辑:裴阿卫)
Spatial Distribution ofPopuluseuphraticaBiomass in the Lower Reaches of Tarim River
WANG Xiyi1,XU Hailiang2*,PAN Cunde1,LING Hongbo2,YUAN Kaiye1
(1 Xinjiang Agricultural University, Urumqi 830052; 2 Xinjiang Institute of Ecology and Geography,Chinese Academy of Sciences,Urumqi 830011,China)
Based on sample-plot survey to thePopuluseuphraticain the lower reaches of Tarim River, we established the model between biomass and factors such as DBH and height; Besides, the distribution of biomass ofP.euphraticawas analyzed in the district; Finally, the total biomass was assessed based on the area ofP.euphraticaforest in the lower reaches of Tarim River. The results are as follows: (1) biomass model ofP.euphratica:B=128.09(D2h)0.6318(R2=0.892),which was high in precision, and the discrepancy between measured value and simulated value was non-significant (P>0.05), thus the model could be used to assess the biomass ofP.euphratica. (2) The biomass presented a trend that the biomass declining was relevant to the distance from river;P.euphraticadistributed within 700 m of river; and the biomass of this area accounted for 91.37% of total. (3) The biomass per unit area was 416.613 kg·hm-2in the lower reaches of Tarim river and the total biomass was 2.474×104t.
Populuseuphratica;biomass;spatial distribution;lower reaches of Tarim River
1000-4025(2016)11-2314-08
10.7606/j.issn.1000-4025.2016.11.2314
2016-07-27;修改稿收到日期:2016-09-21
国家自然科学基金(31370551,41471099, 31400466);中国科学院“西部之光”人才培养计划(XBBS-2014-13);中国科学院特色研究所主要服务项目2课题2(TSS-2015-014-FW-2-2)
王希义(1987-),男,在读博士研究生,主要从事生态需水与恢复生态学研究。E-mail:binzhouwxy@163.com.
*通信作者:徐海量,博士,研究员,硕士生导师,主要从事恢复生态学研究。E-mail:xuhl@ms.xjb.ac.cn
Q948.11;Q948.15+6
A
