四合木根部及茎部导管分子比较解剖学研究
2017-01-04刘果厚刘冠志李红颖
兰 庆,刘果厚*,刘冠志,李红颖
(内蒙古农业大学 草原与资源环境学院,呼和浩特 010011)
四合木根部及茎部导管分子比较解剖学研究
兰 庆1,刘果厚1*,刘冠志1,李红颖1
(内蒙古农业大学 草原与资源环境学院,呼和浩特 010011)
采用显微照相技术对四合木根、茎部导管分子形态特征及数量特征进行比较研究,并运用SMA分析导管分子长度与直径之间的异速生长关系,探讨四合木的次生木质部导管分子对草原化荒漠地区的适应特性。结果表明:(1)四合木根部与茎部导管分子均以孔纹、网纹和两端无尾类型为主,具单穿孔板。(2)四合木根部与茎部导管分子在长度上均为短级水平,直径上均为小级水平,且宽窄导管并存;茎部导管分子较根部导管分子更加“小型化”。(3)不论根部或茎部,其导管分子的长度与直径之间均表现出异速生长的关系,且导管分子长度的增长速率大于直径的增长速率。这些结果有助于从细胞学尺度上了解荒漠区木本植物对草原化荒漠地区的适应策略。
四合木;导管分子;异速生长;荒漠植被
四合木属(TetraenaMaxim.)是蒺藜科(Zygophyllaceae)的单种属,是一个较为进化的属[1],具有重要的科学研究价值。四合木(TetraenamongolicaMaxim.)现已被列为国家二级[2]和内蒙古一级[3]重点保护珍稀濒危植物,主要分布于鄂尔多斯高原西缘至黄河东岸的狭长地带,该地区多为低山丘陵,地貌类型多样,是中国荒漠化草原向草原化荒漠过渡的典型区域,自然条件相对恶劣,具有降水少、风沙大、光照足、无霜期短、昼夜温差大、蒸发强烈等显著的气候特点[4]。目前,关于四合木的形态解剖已做过一些研究[5-8],但关于根、茎次生木质部的结构报道较少。本实验研究了四合木根、茎部导管分子形态特征及数量特征的差异,并对导管分子长度与直径之间的异速生长关系及这种关系在根、茎不同部位间的差异进行了分析,探讨四合木对草原化荒漠地区的适应性。
1 材料和方法
于2015年7月植物生长最旺盛的季节进行材料的采集,随机选取生长状况较好,冠幅、高度相近的四合木植株5株,在直径约0.5 cm枝条上截取1 cm茎段若干,并挖取直径约0.5 cm的根,截取1 cm根段若干,放入FAA固定液中保存带回实验室。将采集的材料分为两部分,一部分使用双面刀片做徒手横切片,厚度大约20 μm,用番红-固绿染色,制成永久装片;另一部分去掉韧皮部后取靠近形成层的边材,置于离析液(过氧化氢∶冰醋酸=1∶1,V/V)内,在60 ℃下离析48 h后去离析液,加蒸馏水,用玻璃棒将材料捣碎至絮状,静置10 min,重复数次,材料用梯度乙醇脱水后,用吸管吸取少量材料,滴在载玻片上,并用番红染色,制成临时装片。
采用Olympus BX51光学显微镜拍照,并利用Motic Images Plus 2.0 ML软件随机选取100个导管分子进行观察,记录导管分子类型,计算各类型所占比例。测量导管长度、直径、端壁角度、管壁厚度等指标,进行3次重复。采用单因素方差分析对其各性状差异进行分析,用Excel 2013、SAS9.0软件进行分析处理。
对导管分子长度与直径异速生长关系的分析,采用Y=βXα来描述,线性转化为:logY=log β + α logX,其中,X与Y代表导管长度和直径,α为方程的斜率,即异速生长指数,log β代表两性状关系的截距。斜率值决定了相关性是等速或异速关系,若α=l(近似等于),表示两性状间呈等距或等速生长,若α≠1(显著偏离),则表示两性状间为异速生长。采用第Ⅱ类回归 (Model Type Ⅱ regression analyses) 来估计异速生长方程的参数,使用标准化主轴估计 (SMA) 的方法来估算方程的斜率[9]。
2 结果与分析
2.1 导管分子形态特征
从四合木根、茎的离析材料观察得出,根部与茎部导管分子均为单穿孔(图版Ⅰ,1~9)。从细胞壁木质化加厚的方式来看(图1),根部导管分子的类型中孔纹导管(图版Ⅰ,1、3)的比例显著大于网纹导管(图版Ⅰ,2)和梯纹导管(图版Ⅰ,4,P<0.05),网纹导管与梯纹导管的比例无显著性差异(P>0.05);茎部导管分子的类型中,网纹导管(图版Ⅰ,7、9)的比例显著大于孔纹导管(图版Ⅰ,6)、梯纹导管(图版Ⅰ,8)和螺纹导管(图版Ⅰ,5)(P<0.05),孔纹导管的比例与梯纹导管、螺纹导管之间均具有显著性差异(P<0.05),梯纹导管与螺纹导管的比例无显著性差异(P>0.05)。根部与茎部导管相比,孔纹导管、网纹导管和梯纹导管的比例均无显著性差异(P>0.05)。根部和茎部导管分子管壁间纹孔排列方式为互列式(图版Ⅰ,3、7)和对列式(图版Ⅰ,2、8)。
从导管分子的具尾情况来看(图2),四合木根部与茎部的导管分子为两端具尾(图版Ⅰ,2、5),一端具尾(图版Ⅰ,4、6)和两端无尾(图版Ⅰ,1、3、7、8)3种类型。根部和茎部导管分子两端无尾的比例分别显著大于各自两端具尾和一端具尾的比例(P<0.05)。其中茎部导管分子两端无尾的比例与根部的相差19%,具显著性差异(P<0.05),其他两种类型无差异。
2.2 导管分子数量特征
表1显示,四合木根部导管分子的平均长度为110.25 μm,茎部为100.64 μm,根据国际木材解剖学家协会理事会(1937年)对导管分子长度的等级规定[10],二者的平均长度均为短级水平。根部导管分子的平均长度显著大于茎部(P<0.05)。根部50个最长导管分子的平均长度为129.72 μm,茎部为118.85 μm,相差10.87 μm,二者差异显著(P<0.05)。并且根部导管分子平均长度远小于50个最长导管分子的平均长度,相差19.47 μm,茎部相差18.21 μm。从四合木根、茎部导管分子长度范围分布情况可以看出(图3),四合木根部导管分子长度主要分布在90~130 μm,占62.05%;茎部导管分子长度主要分布在70~110 μm,占66.67%。

不同小写字母表示不同部位同一指标差异显著(P<0.05)图1 四合木根部和茎部导管分子纹孔类型百分比Values with different small letters indicate significantly differences between different parts of the same index (P<0.05)Fig.1 The percentage of pit types of root and stem vessel elements of T. mongolica
四合木根部导管分子的平均直径为37.87 μm,茎部的为31.33 μm,根据国际木材解剖学家协会理事会(1939年)对导管分子直径的等级规定[10],二者的平均直径均为小级水平。根部导管分子的平均直径与茎部相差6.54 μm,存在显著差异(P<0.05)。
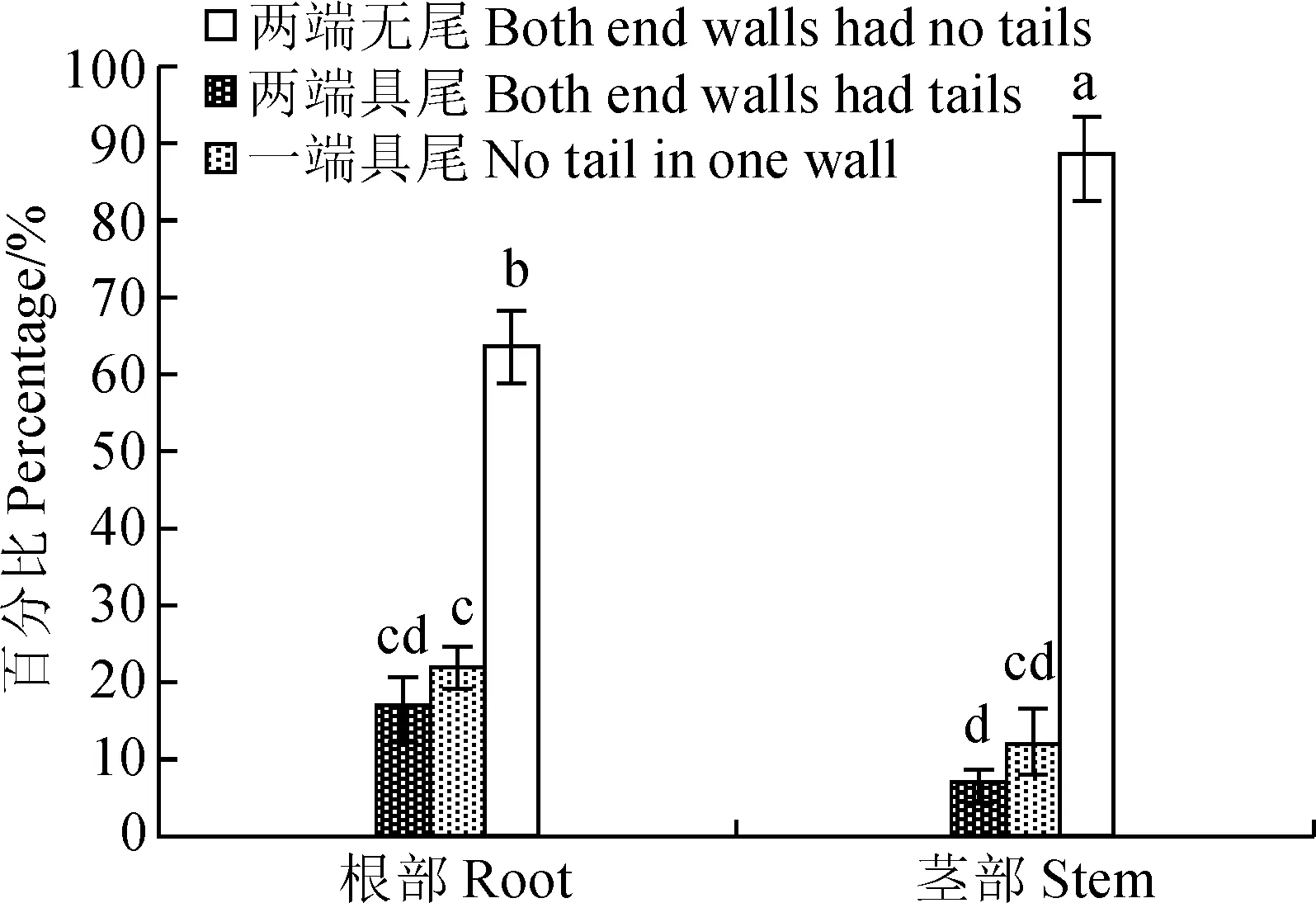
不同小写字母表示不同部位同一指标差异显著(P<0.05)图2 四合木根部和茎部导管分子具尾类型百分比Values with different same normal letters indicate significantly differences between different parts of the same index (P<0.05)Fig.2 The percentage of root and stem vessel elements with tail types of T. mongolica
根部50个最大导管分子的平均直径为52.87 μm,茎部为39.54 μm,相差13.33 μm,同样存在显著差异(P<0.05)。并且根部和茎部的导管分子平均直径远小于各自的50个最大导管分子的平均直径。从四合木根、茎部导管分子直径范围分布情况可以看出(图4),四合木根部导管分子直径主要分布在20~60 μm,占67.5%;茎部导管分子直径主要分布在20~40 μm,占65.28%。
四合木根部导管分子端壁角度为22.18°,茎部为25.04°,无显著差异(P>0.05)。从四合木根、茎部导管分子端壁角度分布(图5)可看出,四合木根部和茎部导管分子的端壁角度都主要分布在0°~40°。
四合木根部导管分子的管壁厚度为2.15 μm,茎部为1.48 μm,相差0.67 μm,有显著差异(P<0.05)。从四合木根、茎导管分子管壁厚度分布(图6)可看出,四合木根部导管分子管壁厚度主要分布在1.5~3 μm,占86.66%;四合木茎部导管分子管壁厚度主要分布在1.0~2.0 μm,占86.67%。
2.3 导管分子长度与直径之间的关系
从表2可以看出,四合木根部导管分子的长度与直径之间的异速生长指数为-0.44(95%置信区间CI=-0.55 ~-0.35),与1.0具有极显著差异(P<0.001),说明其根部导管分子长度与直径之间存在异速生长关系,即导管分子长度与直径间呈非等比例增长,导管分子长度的增长速率大于直径。四合木茎部导管分子的长度与直径之间的异速生长指数为-0.70(95%置信区间CI=-0.87~-0.57),与1.0的差异极显著(P<0.001),说明其茎部导管分子长度与直径之间也存在异速生长关系,导管分子长度的增长速率大于直径。四合木根部导管分子长度、直径与其茎部的导管分子长度、直径之间表现为显著的相关关系,其异速生长指数为-0.56,与1.0具有极显著差异(P<0.001),表现为异速生长关系。

图3 四合木根部和茎部导管分子长度范围Fig.3 The length range of root and stem vessel elements of T. mongolica

表1 四合木根部和茎部导管分子数量特征(平均值±标准差)
注:同行数字后不同字母表示差异显著(P<0.05)
Note: Different letters indicate significant difference in the same row(P<0.05)

表2 四合木根部和茎部导管分子长度与直径的SMA分析结果

图4 四合木根部和茎部导管分子直径范围Fig.4 The diameter range of root and stem vessel elements of T. mongolica

图5 四合木根部和茎部导管分子端壁角度范围Fig.5 The end wall’s angle of root and stem vessel element of T. mongolica

图6 四合木根部和茎部导管分子管壁厚度范围Fig.6 The vessel wall thickness of root and stem vessel element of T. mongolica
3 讨 论
3.1 导管分子的进化特征
四合木被认为旱生植物中较为进化的类群,在茎次生木质部中,其进化特征表现在导管和纤维上[6]。从本研究结果来看,茎部导管分子的形态特征具有原始性。由于发育的先后及细胞壁木质化加厚的方式不同,形成了5种类型的导管,按原始到进化的顺序依次为环纹导管→螺纹导管→梯纹导管→网纹导管→孔纹导管,其中后三者的生理功能效率显著高于前两者,是较为进化的类型[11]。而本研究中,四合木茎部除了存在孔纹导管、网纹导管和梯纹导管外,还存在螺纹导管,此结果与前人[5]得出的一致。从导管分子具尾情况来看,无尾较有尾更为进化[11]。本研究中,根部和茎部导管分子两端无尾的比例分别显著大于各自两端具尾和一端具尾的比例,因此从具尾情况推断四合木次生木质部导管分子较为进化。四合木根、茎次生木质部导管分子的平均长度小于同科植物白刺(NitrariatangutorumBobr.)和霸王(Zygophyllumxanthoxylon(Bge.) Maxim.)次生木质部导管分子的平均长度[5]。3种极具有代表性的旱生植物梭梭(Haloxylonammodendron(C. A. Mey.) Bunge)、无叶假木贼(AnabasisaphyllaL.)和多枝柽柳(TamarixramosissimaLedeb.)次生木质部导管分子平均直径[12]小于四合木根、茎部导管分子的直径。已有学者从四合木的花结构、花粉形态、果实构造及木质部解剖构造多方面进行研究,认为四合木属是蒺藜科较为进化的属[1],因而推测四合木的次生木质部比其他旱生植物更为进化。
3.2 根、茎导管分子差异分析
根部导管分子的平均长度及50个最长导管分子的平均长度均显著大于茎部(P<0.05)。根部导管分子长度主要分布在90~130 μm;茎部导管分子长度主要分布在70~110 μm,说明根部较茎部具长导管性。根部导管分子的平均直径及50个最大导管分子的平均直径均显著大于茎部(P<0.05)。四合木根部导管分子直径主要分布在20~60 μm;茎部导管分子直径主要分布在20~40 μm,说明根部较茎部还具有宽导管性。四合木根部和茎部导管分子端壁角度无显著性差异。四合木根部导管分子的管壁厚度较茎部的有显著性差异(P<0.05),根导管分子管壁厚度主要分布在1.5~3 μm,而茎部导管分子管壁厚度主要分布在1.0~2.0 μm。
对四合木根部、茎部导管分子分别研究表明,导管分子长度与直径间呈非等比例地增长,导管分子长度的增长速率大于直径。对根部和茎部导管的长度、直径综合研究表明,四合木根部导管分子的长、宽增长速度要大于茎部,表现为异速生长关系。已有研究表明,导管分子的某些特征在种间、属间乃至亚科间其进化速度都存在着差异[13-14]。
3.3 导管分子对草原化荒漠地区的适应
四合木根部与茎部导管分子的平均长度均为短级水平,这对于生长在多风的草原化荒漠地区的植物提高抗风沙能力是非常重要的,因为短导管的机械支持力更强。四合木的根系为雨养根系,只依靠降水后下行水的重力生长,其主根系分布在离地面0~30 cm的范围内,侧根系分布在地下0~60 cm范围内[15]。四合木根部和茎部导管分子的平均直径均远小于各自的50个最大导管分子的平均直径,均表现出宽、窄导管并存的特征,这个结果与江莎[6]的一致。导管分子长度越长,直径越大,输水阻力越小,输水效率越高[16]。本研究得出根部导管分子较茎部的直径更大、长度长,说明根部导管分子可以适应干旱区短暂的脉冲式降雨,发挥着较强的输水性,利于旱生植物的生长发育。而茎是植物地上的重要部分,经受干旱的影响,远比根部显著。当其遭遇干旱胁迫时,茎部导管直径相对较小,可以有效地增强水分的内聚力和抗负压能力,进一步保证水分输导的有效性和安全性,提高植物的抗旱能力,维持恶劣环境下植株的正常生长。
分析得出,四合木与同科植物及其他旱生植物相比是较为进化的类群,但是也具有一些原始性。根部与茎部导管的共同特征是:以孔纹,网纹和两端无尾类型为主,具单穿孔板。长度上均为短级水平,直径上均为小级水平,且宽窄导管并存。四合木茎部导管分子较根部导管分子更加“小型化”,具体表现在:导管分子长度、50个最长导管长度、直径、50个最宽导管直径及管壁厚度均显著小于根部,这与在草原化荒漠环境中,茎部需要更强的机械支持力相一致。通过SMA法分析,四合木根部导管分子的长度、直径与其茎部导管分子的长度、直径之间表现出异速生长的关系。对四合木根部、茎部导管分子的分别研究表明,导管分子长度的增长速率大于直径的增长速率。这些特征是荒漠植物生长发育以及进化的结果,保证体内水分运输的安全性和有效性,表现了其对草原化荒漠地区的适应性。
[1] 吴树彪,屠骊珠.从胚胎学特征探讨四合木的系统位置[J].西北植物学报,1990,10(1):23-29.
WU S B, TU L Z. A discussion of systematic position ofTetraenaby embryological feature [J].ActaBot.Boreal.-Occident.Sin.,1990,10(1):23-29.
[2] 国家环境保护局,中国科学院植物研究所.中国珍稀濒危保护植物名录:第一册[M].北京:科学出版社,1987:68.
[3] 内蒙古自治区人民政府.内蒙古珍稀濒危保护植物名录[J].内蒙古林业,1989,8:11-12.
The people’s Government of Inner Mongolia Autonomous Region. Rare and endangered protection plants of Inner Mongolia [J].JournalofInnerMongoliaForestry,1989,8:11-12.
[4] 王 乐,赵利清,陈 育,等.西鄂尔多斯草原化荒漠植物群落多样性[J].干旱区研究,2015,32(2):258-265.
WANG L, ZHAO L Q, CHEN Y,etal. Plant community diversity of steppification desert in West ERDOS [J].AridZoneResearch,2015,32(2):258-265.
[5] 杨淑敏,江泽慧,任海青.8种木本旱生植物的木质部解剖学特性研究[J].西北植物学报,2007,27(8):1 507-1 516.
YANG S M, JIANG Z H, REN H Q. Study on anatomical property of xylem cells of the eight woody xerophytes grown in China [J].ActaBot.Boreal.-Occident.Sin.,2007,27(8):1 507-1 516.
[6] 江 莎,任小华,古 松,等.四合木属四合木(蒺藜科)的木材解剖学研究[J].热带亚热带植物学报,2008,16(5):466-471.
JIANG S, REN X H, GU S,etal. Wood anatomy ofTetraenamongolicaMaxim. (Zygophyllacea) [J].JournalofTropicalandSubtropicalBotany,2008, 16(5):466-471.
[7] YANG Shu-min, FURUKAWA Ikuo. Anatomical adaptations of three species of Chinese xerophytes (Zygophyllaceae) [J].JournalofForestryResearch,2006,17(3):247-251.
[8] 杨淑敏,江泽慧,任海青,等.13种沙生植物纤维和导管分子长度变异的研究及木质部特征的定量研究[J].植物研究,2007,27(5):601-606.
YANG S M, JIANG Z H, REN H Q,etal. Quantitative characteristics of xylem cells and variation in vessel element length and fiber length for 13 psammophytes [J].BulletinofBotanicalResearch,2007,27(5):601-606.
[9] WARTON D I, WRIGHT I J, FALSTER D S,etal. Bivariate line-fitting methods for allornetry[J].BiologicalReviews,2006,81(2):259-291.
[10] 成俊卿.木材学[M].北京:中国林业出版社,1985:102-104.
[11] TAKHTAJAN A. Morphological Evolution of the Angiosperms [M]. Moscow: Agricultural Press,1984.
[12] 楚光明,刘 娜,牛攀新,等.准噶尔盆地3种荒漠植物木质部导管解剖特征[J].干旱区资源与环境,2016,30(2):104-110.
CHU G M, LIU N, NIU P X,etal. The anatomy characteristics of xylem vessel of three typical desert plants in Junggar basin[J].JournalofAridLandResourcesandEnvironment,2016,30(2):104-110.
[13] 李玉斌,宋治江,徐 伦,等.12种豆科植物次生木质部导管分子比较观察[J].黄冈师专学报1992,12(1):38-41.
LI Y B, SONG Z J, XU L,etal. Comparative studies on the vessel elements of 12 species in Leguminosae[J]. JournalofHuanggangTeachersCollege, 1992,12(1):38-41.
[14] 刘冠志,刘果厚,等.黄柳与小红柳导管分子形态特征及其生态适应性比较研究[J].西北植物学报,2016,36(2):316-322.
LIU G Z, LIU G H,etal. Comparative study on morphological characteristics and ecological adaptability of vessel elements ofSalixgordejeviiandS.microstachyavar.bordensis[J].ActaBot.Boreal.-Occident.Sin.,2016,36(2):316-322.
[15] 张永明,高润宏,金 洪.西鄂尔多斯荒漠4种灌木根系生态特性研究[J].内蒙古农业大学学报,2005,26(3):39-43.
ZHANG Y M, GAO R H, JIN H. Studies on the ecological characteristics of four bushes roots in desert of West ERDOS [J].JournalofInnerMongoliaAgriculturalUniversity,2005,26(3):39-43.
[16] 李国秀,郑宝江.10种茶藨子属植物导管分子形态特征及其生态适应性比较研究[J].植物研究,2014,34(1):25-31.
LI G X, ZHENG B J. Comparative study on morphological characteristics and ecological adaptability of vessel elements of tenRibesL. varieties[J].BulletinofBotanicalResearch, 2014,34(1):25-31.

图版Ⅰ 光学显微镜下四合木导管分子形态1~4.根部导管分子,×400 5~9.茎部导管分子,×400PlateⅠ The characteristics of root and stem vessel elements of T. mongolica1-4. Root vessel element, ×400 5-9. Stem vessel element, ×400
(编辑:潘新社)
Comparative Anatomy of Root and Stem Vessel Elements ofTetraenamongolica
LAN Qing1,LIU Guohou1*,LIU Guanzhi1,LI Hongying1
(College of Grassland, Resources and Environment, Inner Mongolia Agricultural University, Hohhot, 010011 China)
The study used photomicrographic technique to compare the morphological characteristics and quantity characteristics between the root and stem vessel elements ofTetraenamongolica, and used the SMA to analyze the allometric relation between the length and diameter of vessel elements, in order to discuss the adaptability of secondary xylem elements ofT.mongolicato the grassland desertification area. The results showed that: (1) the root and stem vessel elements ofT.mongolicaare mainly pitted and reticulate catheter types, and both have no tail, and both of them have simple perforation plate; (2) the root and stem vessel elements ofT.mongolicawhich catheter length are short grade level, and their catheter diameter are small short level and both have wide and narrow vessel elements, and stem vessel elements are more “miniaturization” than that of roots; (3) regardless of the roots or stems, their length and diameter of the catheter molecules exhibit allometric relationship, and the growth rate of vessel length is greater than that of diameter. These results are helpful to understand the adaptation strategies from catheter scale in desert area of steppe desert region.
Tetraenamongolica; vessel element; allometry; desert plants
1000-4025(2016)11-2220-06
10.7606/j.issn.1000-4025.2016.11.2220
2016-08-05;修改稿收到日期:2016-11-01
内蒙古自治区应用技术研究与开发资金计划(20131902)
兰 庆(1992-),女,在读硕士研究生,主要从事植物多样性保护与利用研究。E-mail:1246055889@qq.com
*通信作者:刘果厚,教授,博士生导师,主要从事植物多样性保护与利用研究。E-mail:guohouliu@163.com
Q944.5
A
