普通小麦/ 山羊草属间杂交F1的形态学及细胞遗传学研究
2016-12-30彭娜娜王亚娟王长有吉万全
彭娜娜,王亚娟,王长有,权 威,吉万全
(西北农林科技大学农学院,陕西杨凌 712100)
普通小麦/ 山羊草属间杂交F1的形态学及细胞遗传学研究
彭娜娜,王亚娟,王长有,权 威,吉万全
(西北农林科技大学农学院,陕西杨凌 712100)
为了解普通小麦与山羊草属的杂交F1代的育性、细胞遗传学表现及育种利用潜力,以普通小麦7182和丰优1718为母本,分别以卵穗山羊草SY163和离果山羊草SY119为父本进行杂交,对杂交F1代的生育特性、染色体构型、田间条锈病抗性、F1自交和回交结实性等进行了分析。结果表明,F1代植株生长势较强,株型倾向母本,成熟期穗子易断,壳硬;杂交F1代高度不育,花粉可育率1.26%~3.54%,回交结实率2.08%~5.26%,仅普通小麦/卵穗山羊草自交结实,但结实率最高仅为0.25%。7182/SY163、7182/SY119、丰优1718/SY163、丰优1718/SY119杂交F1代花粉母细胞减数分裂中期I的染色体构型平均分别为31.11I + 1.90II + 0.03III、20.04I + 7.45II + 0.02III、30.08I + 2.40II + 0.04III、22.37I +6.30II + 0.01III,说明普通小麦与2种山羊草杂交亲和性差,且可交配性主要由山羊草基因决定。田间条锈病抗性鉴定结果显示,F1代均对条锈菌混合小种高抗或免疫,这对于丰富小麦抗病基因资源有重要意义。
普通小麦;卵穗山羊草;离果山羊草;生育特性;染色体构型;条锈病
山羊草属(Aegilops)植物与小麦亲缘关系较近,全属有24个或更多个种,包括C、D、M和U等l7个染色体组,蕴藏着许多抗病、抗虫、抗逆等有益基因,将山羊草属植物与小麦远缘杂交,是获得新种质并从其中转移有益基因的重要手段[1-6]。卵穗山羊草(Ae.geniculataRoth.,2n=2x=28,UUMM)具有抗白粉病、叶锈病、条锈病、秆锈病基因,而且具有高蛋白特性[1];离果山羊草(Ae.triuncialisL.,2n=2x=28,UUCC)含有抗白粉病、叶锈病、条锈病基因,并且3C染色体上含有控制不育性状的基因[7-15],均是麦类作物品种改良的丰富和宝贵种质基因库。国内目前关于山羊草与小麦杂交的研究并不多,且大多是利用一种山羊草种与小麦进行杂交,预测其杂交亲和性,两种山羊草进行比较的报道鲜见。本研究以普通小麦7182和丰优1718为母本,分别以卵穗山羊草SY163和离果山羊草SY119为父本进行杂交,对其杂交F1代的形态学、细胞学和抗病性进行研究,旨在为小麦与卵穗山羊草和离果山羊草杂交的育种潜力进行评价及作为种质资源的利用提供依据。
1 材料与方法
1.1 材 料
供试母本为普通小麦(Triticumaestivum,2n=42,染色体组AABBDD)品种7182、丰优1718,由本实验室保存;父本为卵穗山羊草SY163(Ae.geniculata,2n=28,染色体组UUMM)、离果山羊草SY119(Ae.triuncialis,2n=28,染色体组UUCC),由中国农科院作物科学研究所李立会博士和杨欣明老师惠赠。7182/ SY163、丰优1718/SY163、7182/SY119、丰优1718/SY119杂交种子由本实验室获得并保存。
1.2 方 法
1.2.1 形态学和田间抗条锈病鉴定[16]
2014年10月8日将杂交亲本和F1种子种植在西北农林科技大学农学院试验地,肥力中等,每份种植2行,行长1 m,行间距0.25 m,在生育期考察分蘖、芒、壳、粒色、株高、穗长、穗粒数等农艺性状。2015年3月25日傍晚接条中31、32和水源14混合小种,15 d 后进行条锈病分级调查,7 d后复查,划分标准0~4级。
1.2.2 花粉育性及结实性观察
(1)花粉育性:采用醋酸洋红染色法,每种材料观察300个视野。形状规则,大小整齐,染色深为可育;形状不规则,大小参差不齐,染色浅为不育。花粉可育率=可育花粉粒总数/观察花粉粒总数×100%。
(2)结实率:对F1自交套袋和回交后代统计结实情况。在种子成熟期,收获所有植株上的穗子,进行室内考种,统计结实数和小花数,结实率=结实总数/观察小花总数×100%。
1.2.3 细胞学鉴定
(1)根尖细胞染色体观察:2015年3月下旬,田间挖取F1代植株的根尖,冰水中处理24 h;放入卡诺氏固定液I(无水乙醇∶冰乙酸=3∶1)中固定24 h。将固定好的根尖放入1 mol·L-1冰乙酸中,煮沸解离3~4 min;1%醋酸洋红染色,压片后显微镜下观察。
(2)花粉母细胞减数分裂中期I(PMC MI)染色体观察:2015年4月中旬,田间剪取F1代植株孕穗期的幼穗,用卡诺氏固定液II(无水乙醇∶氯仿∶冰乙酸=6∶3∶1)固定24 h,4 ℃冰箱保存备用。花粉母细胞用1%醋酸洋红染色制片。使用Olympus BX 51显微镜观察统计100个细胞,并记录PMC MI的染色体配对构型。
2 结果与分析
2.1 农艺性状调查结果
田间调查发现,母本(普通小麦品种7182和丰优1718)的穗粒数较父本(卵穗山羊SY163和离果山羊草SY119)多,籽粒饱满,植株较低,但田间高感条锈病。父本的分蘖数较高,均超过50个,对条锈病免疫,但在成熟期容易发生穗轴断裂,壳硬,不易脱粒。杂交F1单株分蘖较多,介于双亲之间,其穗在成熟期容易在穗轴处断裂(表1)。由图1可知,普通小麦7182穗形为长方形,丰优1718为纺锤形,两种山羊草穗形都为纺锤形;在杂交F1代中,除7182/SY163为长方形外,其他都为纺锤形,其穗长介于两亲本之间。经过田间条锈病抗性鉴定,杂交F1代对供试条锈菌混合小种均表现高抗至免疫(图1e)。
2.2 结实性和花粉育性
由表2可以看出,7182/SY119、丰优1718/ SY119的F1自交结实率均为0,7182/SY163、丰优1718/SY163 F1的自交结实率分别为0.15%、0.25%。表明远缘杂交F1的自交系高度不育,且其育性因山羊草种的不同而异。由表3和表2可以看出,杂交F1代与普通小麦回交结实率为2.08%~5.26%,杂交F1代的花粉可育率为1.26%~3.54%,可育率极低。田间试验观察发现,杂交F1代花药瘦小不开裂,花粉干瘪,结实率较低。
表1 亲本及杂交F1代的形态特性及抗条锈性
Table 1 Morphological characters and stripe rust resistance of parents and hybrid F1

材料Material芒长Awnlength/cm株高Height/cm穗长Spikelength/cm穗形Typeofspike分蘖数Numberoftiller条锈病抗性Resistancetostriperust71826.185.110.2长方形Rectangle154丰优1718 Fengyou17185.280.38.5纺锤形Spindle124SY1634.350.14.5纺锤形Spindle600SY1194.670.56.3纺锤形Spindle5407182/SY1633.240.25.3长方形Rectangle450⁃17182/SY1195.564.29.1纺锤形Spindle350⁃1丰优1718/SY163 Fengyou1718/SY1636.166.17.5纺锤形Spindle300⁃1丰优1718/SY119 Fengyou1718/SY1194.180.18.2纺锤形Spindle250⁃1

a:7182、SY163及其F1;b:7182、SY119及其F1; c:Fengyou 1718、SY163及其F1; d:Fengyou 1718、SY119及其F1; e:7182、SY119及其F1的抗条锈性;1:7182;2:SY119;3:7182/SY119的F1
a:7182,SY163,(7182/SY163)F1;b:7182,SY119,(7182/SY119)F1; c:Fengyou 1718,SY163,(Fengyou 1718/SY163)F1; d:Fengyou 1718,SY119,(Fengyou 1718/SY119)F1; e:The resistance to stripe rust of 7182,SY119 and their F1;1:7182;2:SY119;3:(7182/SY119)F1
图1 亲本及杂交F1代的农艺性状比较
Fig.1 Comparisons of agronomic characters of parents and hybrid F1
表2 杂交F1代的花粉育性和结实率
Table 2 Pollen fertility and seed setting rate of hybrid F1

组合Combination统计总数StatisticalNo.可育数FertilityNo.可育率Fertilityrate/%结实率Seedsettingrate/%7182/SY163360123.330.257182/SY119392102.550丰优1718/SY163 Fengyou1718/SY16331841.260.15丰优1718/SY119 Fengyou1718/SY119395143.540
2.3 花粉母细胞染色体特征
普通小麦为六倍体(2n=6x=42),卵穗山羊草与离果山羊草都为四倍体(2n=4x=28),7182/SY163、7182/SY119、丰优1718/SY163、丰优1718/SY119的杂交F1代根尖有丝分裂中期染色体计数表明,杂交F1代具有预期的染色体数,2n=5x=35(图3),即属间杂交F1染色体数目是双亲的配子染色体数目总和,为五倍体。
表3 杂交F1代回交结果
Table 3 Backcross of hybrid F1
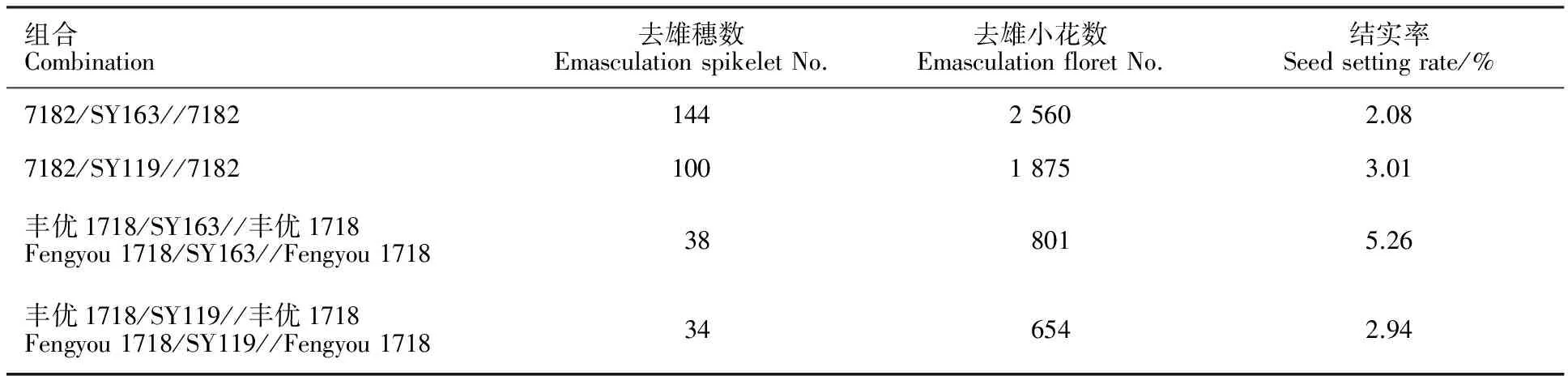
组合Combination去雄穗数EmasculationspikeletNo.去雄小花数EmasculationfloretNo.结实率Seedsettingrate/%7182/SY163//718214425602.087182/SY119//718210018753.01丰优1718/SY163//丰优1718Fengyou1718/SY163//Fengyou1718388015.26丰优1718/SY119//丰优1718Fengyou1718/SY119//Fengyou1718346542.94
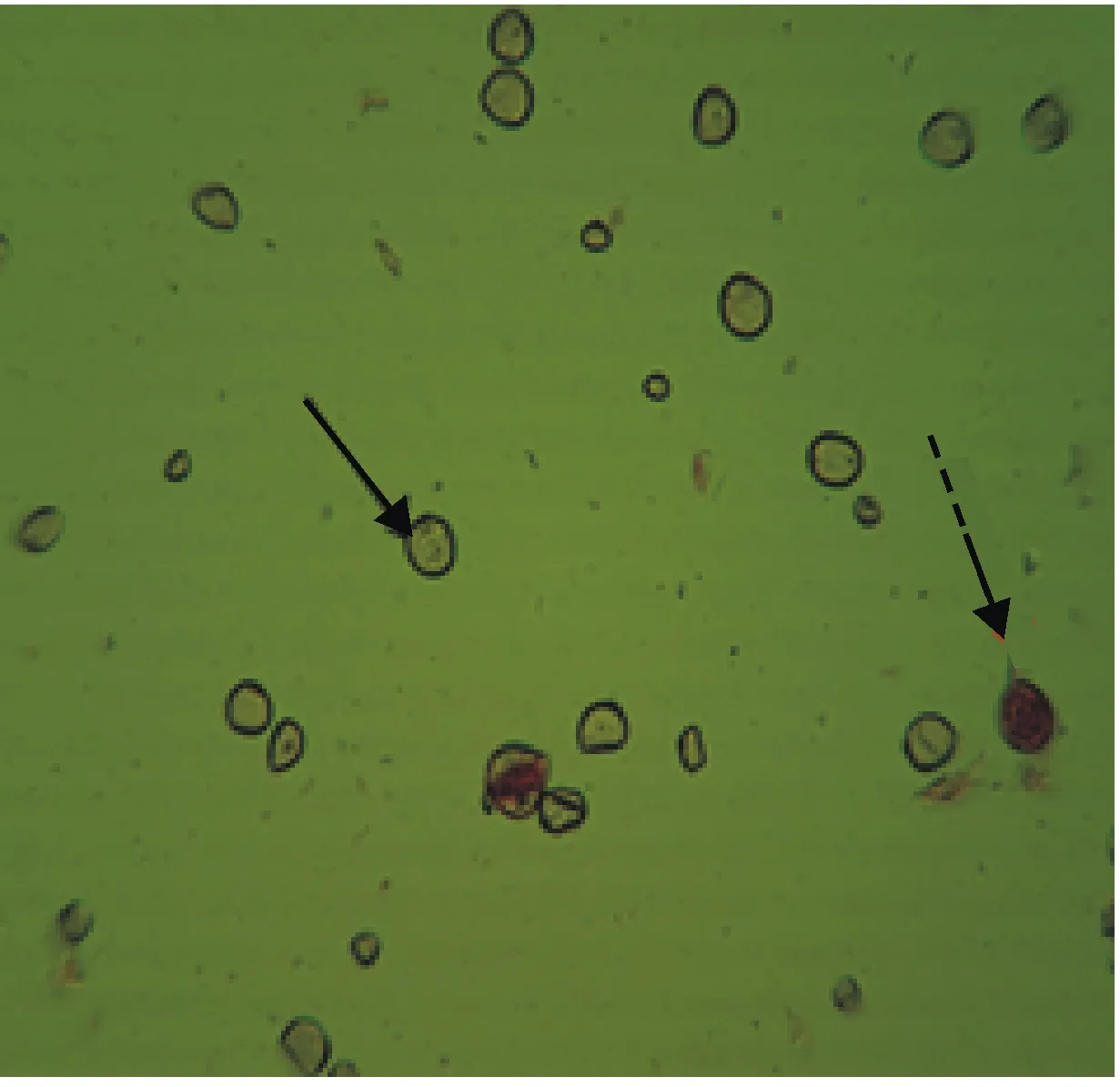
→箭头所指为败育花粉;---→箭头所指为可育花粉。
→ arrow represents sterile pollen,---→arrow represents fertile pollen.
图2 7182/SY163杂交F1代的花粉
Fig.2 Pollens of 7182/SY163 F1
对4个组合的杂交F1代PMC MI染色体配对情况进行观察统计(表4)发现,杂交F1代(2n=35)绝大部分染色体以单价体存在,频率为20.04~31.11,二价体频率较低,为1.90~7.45,多价体频率极低,为0.01~0.04,表明双亲的染色体同源性很低。且父本为卵穗山羊草的杂交F1代比父本为离果山羊草的杂交F1代单价体多,分别为31.11、30.08和20.04、22.37。由图4可看出,在PMC MI染色体二价体多为棒状且联会松弛,表明五倍体F1代的PMC MI染色体行为很不规律。这为杂交F1代高度不育提供了细胞遗传学依据。

a:7182/SY163杂交F1代;b:7182/SY119杂交F1代;c:丰优1718/SY163杂交F1代;d:丰优1718/SY119杂交F1代
a:7182/SY163 F1; b:7182/SY119 F1; c:Fengyou 1718/SY163 F1; d:Fengyou 1718/SY119 F1
图3 杂交F1代体细胞染色体组型
Fig.3 Somatic cell chromosome number of hybrid F1
表4 杂交F1代的PMC MI染色体构型
Table 4 Chromosome pairing configuration of F1at PMC MI

组合Combination2n细胞数CellNo.IIIIII7182/SY1633510031.11(27~35)1.90(0~4)0.03(0~1)7182/SY1193512820.04(15~29)7.45(3~8)0.02(0~1)丰优1718/SY163 Fengyou1718/SY1633514030.08(25~35)2.40(0~5)0.04(0~1)丰优1718/SY119 Fengyou1718/SY1193516322.37(19~31)6.30(2~12)0.01(0~1)
I:单价体; II:二价体; III:三价体。
I:Univalent; II:Bivalent; III:Trivalent.

a:7182/SY163杂交F1代;b:7182/SY119杂交F1代;c:丰优1718/SY163杂交F1代;d:丰优1718/SY119杂交F1代
a:7182/SY163 F1; b:7182/SY119 F1; c:Fengyou 1718/SY163 F1; d:Fengyou 1718/SY119 F1
图4 杂交F1代的PMC MI构型
Fig.4 Chromosome configuration of chromosome PMC MI in hybrid F1
3 讨 论
3.1 山羊草属农艺性状的多样性研究及其利用
长期人为定向选择严重限制了小麦遗传基础,增加了小麦育种工作取得突破性进展的难度。小麦近缘属有许多普通小麦所不具备的优良性状,可以作为优异基因的来源。山羊草属是普通小麦的一个近缘属,其中蕴藏着许多抗病、抗虫、抗逆等有益基因,属于普通小麦改良的外源基因导入的三级基因库,目前主要是通过远缘杂交将一些有益基因定向转移到小麦中。山羊草的穗轴在成熟期容易断裂,开花期持续长[17],在杂交过程中,应选择生育期相近的普通小麦与山羊草做杂交,才更有利于授粉,使有益基因更易转移到小麦中,达到改良小麦的目标。
3.2 山羊草属在小麦育种中育性机理的分析
本试验发现,在杂交F1植株的PMC MI染色体构型中,不仅有二价体,也有三价体,这表明小麦与两种山羊草染色体之间存在部分同源性。染色体之间的联会配对,使山羊草与普通小麦之间进行基因交流,将山羊草的优良基因转移到普通小麦中存在可能性。
普通小麦与山羊草染色体组类型和数目差异较大,在F1中,来自双亲的染色体在后代分离中出现染色体的重组,但同源染色体配对频率低,相互配对的染色体很少,产生可育配子的几率很低,两种可育配子结合的几率更低,并且普通小麦的5B、3A、4D等染色体上的ph基因对染色体配对也有影响[18],使本试验中两种山羊草与小麦杂交时,结实性较差,杂交F1代高度不育。Riley等[19]报道,尾状山羊草(2n=2x=14,CC)与小伞山羊草(2n=2x=14,UU)染色体中有ph基因作用的抑制因子,并且C和U基因组中ph基因抑制因子联合作用高于单个基因组中抑制基因的作用效果。在本试验中,丰优1718/SY119杂交F1代细胞,最多可观察到12个二价体,而在丰优1718/SY163的杂交F1代细胞中,最多只可观察到5个二价体,SY119染色体组构成为UUCC,SY163为UUMM,证实了C、U基因组中ph基因抑制因子的作用,并且由于染色体的不平衡,有功能性的配子与无功能性的配子之间相互作用,使外源有利基因的重组个体比例变少,这也能部分解释杂种中,同源配对较多,获得有用后代个体频率较低的现象[20]。
叶兴国等[18]认为,小麦与山羊草的可交配性主要由山羊草的基因型决定,普通小麦的基因型对杂交结实率无明显影响。Endo等[21]发现,在小麦背景中,离果山羊草的3C染色体具有优先传递的作用,当离果山羊草3C染色体处于单体状态时,会导致后代不含有杀配子染色体的配子中产生缺失和易位等染色体结构,从而使配子不育[22-23]。从本试验的结果看,在杂交后代中,单价体频率以卵穗山羊草为亲本的后代较多,以离果山羊草为亲本的后代较少,自交结实率却以卵穗山羊草为亲本的后代较高,这与叶兴国及Endo等[18,21]研究结果一致。
3.3 卵穗山羊草在小麦育种改良中的贡献
卵穗山羊草在小麦育种利用中已有较多报道,例如Friebe等[24]用C分带的方法区分出了卵穗山羊草的染色体组,并创造了9个双端体附加系,2个单端体附加系和一整套二体异附加系;Ervin等[25]利用Friebe的整套二体异附加系开发出了一批卵穗山羊草特异的S-SAP标记;谢 纳等[26]证明普通小麦-卵穗山羊草异附加系对麦长管蚜高抗,其广泛的分布使其对各种逆境有极强的适应性;Zeller等[27]发现并证明了7D染色体上含有一个源于卵穗山羊草的抗白粉病基因 Pm29; Kuraparthy等[28]在卵穗山羊草的5Mg发现了两个新的抗病基因,一个是条锈病抗性基因 Yr40,另一个是叶锈病抗性基因 Lr57,这为其用于普通小麦的遗传改良提供了广泛的前景。本试验中,杂交F1代植株生长势强,且均对条锈菌混合小种高抗至免疫,在小麦育种中,可以通过远缘杂交将这些优良基因转到普通小麦中,以丰富小麦的遗传背景。
[1] 孔令让,董玉琛.粗山羊草(Aegilopstauschii)有效利用研究进展[J].山东农业大学学报,1998,29(4):543.
KONG L R,DONG Y C.Progress of effective utilization ofAegilopstauschii(Coss.)schmal[J].JournalofShandongAgriculturalUniversity,1998,29(4):543.
[2] 王洪刚,刘树兵,高居荣,等.小麦与钩刺山羊草杂种的育性,抗病性和细胞遗传学研究[J].麦类作物学报,2000,20(3):1.
WANG H G,LIU S B,GAO J R,etal.Study on fertility,disease resistance and cytogenetics of hybrid betweenTriticumaestivumandAegilopstriuncialis[J].JournalofTriticeaeCrops,2000,20(3):1.
[3] 樊 路,韩敬花,潘淑婷,等.(普通小麦×山羊草)F1用普通小麦回交的初步研究[J].遗传,1993,15(3):25.
FAN L,HAN J H,PAN S T,etal.Preliminary study of backcross of(common wheat×Aegilops)F1with common wheat [J].Hereditas,1993,15(3):25.
[4] CENCI A,DOVIDIO R,TANZARELLA O A,etal.Identification of moleculer markers linked to Pm13,anAegilopslongissimagene conferring rrsistance to powdery mildew in wheat [J].Theoretical&AppliedGenetics,1999,98(3-4):449.
[5] JIA J Z,DEVOS K M,CHAO S,etal.RFLP-based maps of the homoeologous group-6 chromosomes of wheat and their application in the tagging of Pm12,a powdery mildew resistance gene transferred fromAegilopsspeltoidesto wheat [J].Theoretical&AppliedGenetics,1996,92:560.
[6] GILL B S,RAUPP W J.Evaluation ofAegilopsspecies for resistance to wheat powdery mildew,wheat leaf rust,hessianfiy,and green bug [J].PlantDisease,1985,69:314.
[7] 叶兴国,樊 路. ph1b,ph2a,ph2b基因在小麦与卵穗山羊草,小伞山羊草,离果山羊草F1杂种中的作用[J].作物杂志,1993,9(1):16.
YE X G,FAN L.The role of Ph1b,Ph2a,Ph2b in wheat andAe.ovata,Ae.umbellulata,Ae.triuncialishybrid F1[J].Crops,1993,9(1):16.
[8] 金善宝.中国小麦学[M].北京:中国农业出版社,1996:260.
JIN S B.Chinese Wheat [M].Beijing:China Agriculture Press,1996:260.
[9] 薛秀庄,王祥正,吉万全,等.小麦染色体工程与育种[M].石家庄:河北科学技术出版社,1993:78.
XUE X Z,WANG X Z,JI W Q,etal.Chromosome Engineering and Breeding of Wheat [M].Shijiazhuang:Heibei Science and Technology Press,1993:78.
[10] 叶兴国,徐惠君,杜丽璞.东方山羊草,尾状山羊草中染色体配对控制机制的存在和作用[J].遗传学报,1997,24(2):149.
YE X G,XU H J,DU L P.Discovery of mechanisms on homoeologous chomosome pairing inAegilopsaucheriandAe.caudata[J].ActaGeneticaSinica,1997,24(2):149.
[11] 王洪刚,亓增军,孔凡晶,等.(普通小麦/长穗偃麦草)F1小孢子发生和雄配子发育的细胞学特点[J].西北植物学报,1998,18(1):15.
WANG H G,QI Z J,KONG F J,etal.Gytological characteristic on microsporogenesis and male gametophyte development of(T.aestivum/E.elongata)F1[J].ActaBotanicaBorealOccidentSinoca,1997,24(2):15.
[12]FRIEBE B,HEUN M,TULEEN N,etal.Cytogenetically monitored transfer of powdery mildew resistance from rye into wheat [J].CropScience,1994,34(3):621.
[13] 孔秀英,周荣华,董玉琛,等.尾状山羊草与硬粒小麦、普通小麦的杂交及外源染色体检测[J].植物学报,1999,41(11):1164.
KONG X Y,ZHOU R H,DONG Y C,etal.Hybridization ofAegilopscaudatawithTriticumdurum,Triticumaestivumand the detection of alien chromatins [J].ActaBotanicaSinica,1999,41(11):1164.
[14] 孔令让,董玉琛,张红军,等.普通小麦与粗山羊草正,反交的育性研究[J].西北植物学报,2000,20(4):519.
KONG L R,DONG Y C,ZHANG H J ,etal.Studies on the fertility of original cross and reciprocal cross betweenT.aestivumandAe.tauschii[J].ActaBotanicaBorealOccidentSinica,2000,20(4):519.
[15]林 娜,颜泽洪,魏育明,等.钩刺山羊草(Aegilopstriuncialis)低分子量谷蛋白基因序列分析[J].农业生物技术学报,2006,14(4):569.
LIN N,YAN Z H,WEI Y M,etal.Sequence analysis of LMW-GS genes fromAegilopstriuncialis[J].JournalofAgriculturalBiotechnology,2006,14(4):569.
[16] 李立会,杨欣明,李秀全,等.小麦种质资源描述规范和数据标准[M].北京:中国农业出版社,2006:36.
LI L H,YANG X M,LI X Q,etal.Descriptors and Data Standard for Wheat(TririeunaestivumL.) [M].Beijing:China Agriculture Press,2006:36.
[17] 王亚娟,王耀勇,张德华,等.节节麦农艺性状及高分子量谷蛋白亚基遗传多样性研究[J].西北植物学报,2007,27(10):1971.
WANG Y J,WANG Y Y,ZHANG D H,etal.Genetic diversity of agronomic traits and high molecular weight glutenin subunits inAegilopssquarrosa[J].ActaBotanicaBorealOccidentSinina,2007,27(10):1971.
[18] 叶兴国,董建力.宁夏小麦品种与黑麦,离果山羊草杂交研究初报[J].宁夏农林科技,1992(4):15.
YE X G,DONG J L.The study of Ningxia wheat and rye,Aegilopstriuncialishybrization [J].NingxiaJournalofAgricultureandForestryScienceandTechnology,1992(4):15.
[19] RILEY R,KIMBER G,CHAPMAN V J.Origin of genetic control of diploid like behavior of polyploid wheat [J].JouralofHeredity,1961,52:22.
[20] 孔秀英,董玉琛,周荣华,等.普通小麦与钩刺山羊草杂交F1的农艺性状及细胞遗传学的观察[J].植物学报,2000,42(8):845.
KONG X Y,DONG Y S,ZHOU R H,etal.Agronomic characteristics and cytogenetic observation of the hybrid F1fromTriticumaestirumandAegiliopstriuncialis[J].ActaBotanicaSinica,2000,42(8):845.
[21] ENDO T R.Induction of chromosomal structure changes by a chromosome ofAegilopscylindricalL. in common wheat [J].JournalofHeredity,1998,79:366.
[22] 陈全战,亓增军,冯祎高,等.利用离果山羊草3C染色体诱导簇毛麦4V染色体结构变异[J].遗传学报,2002,29(4):356.
CHEN Q Z,QI Z J,FENG Y G,etal.Structural changes of 4V chromosome ofHaynaldiavillosainduced by gametocidal chromosome 3C ofAegilopstriuncialis[J].ActaGencticaSinica,2002,29(4):356.
[23] 陈全战,曹爱忠,亓增军,等.利用离果山羊草3C染色体诱导簇毛麦2V染色体染色体结构变异[J].中国农业科学,2008,(2):363.
CHEN Q Z,CAO A Z,QI Z J,etal.Structural change of 2V chromosome ofHaynaldiavillosainduced by gametocidal chromosome 3C ofAegilopstriuncialis[J].ScientiaAgriculturaSinica,2008(2):363.
[24] FRIEBD B R,TULEEN N A,GILL B S.Development and identification of a complete set ofTriticumaestivum-Aegilopsgeniculatachromosome addition lines [J].Genome,1999,42(3):375.
[25] NAGY E D,MOLNAR I,SCHNEIDER A,etal.Characterization of chromosome-specific S-SAP markers and their use in studying genetic diversity inAegilopsspecies [J].Genome,2006,49(4):289.
[26] 谢 纳,王 雷,齐珊珊,等.小麦近缘种属对麦长管蚜抗性分析[J].河南科学,2014,32(6):1020.
XIE N,WANG L,QI S S,etal.Resistance analysis of wheat relatives to sitobion avenve [J].Henanscience,2014,32(6):1020.
[27] ZELLE F K,HARTL L,MOHLER L,etal.Chromosomal location of genes for resistance to powdery mildew in common wheat(TriticumaestivumL.emThell.) 7.Gene Pm29 in line Pova [J].Euphytica,2002,123(2):187.
[28] KURAPARTHY V C,PARVEEN D,HARCHARAN S K,etal.Characterization and mapping of cryptic alien introgression fromAegilopsgeniculatawith new leaf rust and stripe rust resistance genes Lr57 and Yr40 in wheat [J].Theoretical&AppliedGenetics,2007,114(8):1379.
Study on Morphological and Cytogenetics of F1Hybrids betweenTriticumaestivumandAegilopsspecies
PENG Nana,WANG Yajuan,WANG Changyou,QUAN Wei,JI Wanquan
(College of Agronomy,Northwest Agriculture and Forestry University,Yangling,Shaanxi 712100,China)
In order to investigate the compatibility,cytogenetic basis and breeding potential for selecting and utilizing intergeneric hybrid between wheat andAegilopsspecices,two common wheat varieties 7182 and Fengyou 1718 were used as female parent crossed withAe.geniculataandAe.triuncialis. F1was selfing and backcrossed with its female parent. The growth,fertility,cytogenetic characters,resistance to stripe rust,and the ripening rate of hybrid F1selfing and backcrossed lines were analyzed with the methods of measuring fixed plants,checking chromosome slide and staining pollen. The results showed that hybrid F1had stronger growth potentiality,maternal parent-like plant type,easy-broken spike at maturity,hard glumes,low pollen fertility with fertility rate of 1.26%-3.54% and the backcross seed setting rate of 2.08%-5.26%. Only common wheat/Ae.geniculatahas cross-compatibility,with a maximun setting percentage of 0.25%. The chromosome configurations in F1PMC MI were 31.11I + 1.90II + 0.03III,20.04I + 7.45II + 0.02III,30.08I + 2.40II + 0.04III and 22.37I +6.30II + 0.01III. It was suggested that the cross-compatibility are poor between the common wheat and twoAegilopsspecies,and the compatibility was controlled by theAegilopsgene mainly. All F1plants showed high resistance to mixture stripe rust race,which is significant for disease resistance gene resources of wheat.
Triticumaestivum;Ae.geniculata;Ae.triuncialis; Characters of growth and development; Chromosome configurations; Stripe rust
2016-04-08
2016-05-24
国家自然科学基金项目(31471481);陕西省自然科学基金项目(2015JQ3078);西北农林科技大学基础科研业务经费项目(2014YB080,2452015268);西北农林科技大学博士启动经费(2013BSJJ056)。
E-mail:pengnn0910@163.com
吉万全(E-mail:jiwanquan2003@126.com)
时间:2016-12-07
S512.1;S330
A
1009-1041(2016)12-1563-07
网络出版地址:http://www.cnki.net/kcms/detail/61.1359.S.20161207.1748.002.html
