长江营养盐浓度的增加对邻近海区海-气CO2通量的影响
2016-12-28罗晓凡
刘 哲,罗晓凡,魏 皓,赵 亮
(1. 天津科技大学海洋与环境学院,天津 300457;2. 天津大学海洋科学与技术学院,天津 300072)
长江营养盐浓度的增加对邻近海区海-气CO2通量的影响
刘 哲1,罗晓凡1,魏 皓2,赵 亮1
(1. 天津科技大学海洋与环境学院,天津 300457;2. 天津大学海洋科学与技术学院,天津 300072)
近50年来,长江溶解无机氮和溶解无机磷营养盐浓度不断增加,对长江口邻近海区生态系统产生影响的同时,也对海-气CO2通量带来扰动.基于三维的东中国海生态动力-无机碳循环耦合模型,设计敏感性实验,在不考虑营养盐结构变化的前提下,假设长江营养盐浓度各增加50%,,模拟长江营养盐输入增加情景,探究其对长江口邻近海区海-气CO2通量的影响.结果表明:与参考实验相比,长江营养盐浓度的增加使得夏季长江浅滩大气CO2向海通量增加,海表可多吸收约68%,的大气CO2,而冬季长江浅滩和苏北浅滩大气CO2向海通量减小,即冬季碳汇作用减弱,全年尺度上长江浅滩吸收CO2的能力增强约6%,.长江营养盐浓度增加所致的生物效应主控长江口邻近海区溶解无机碳的变化,进而对海-气CO2通量产生影响.
海-气CO2通量;营养盐;长江口;生态模型
河流向海洋输送了大量淡水、溶解无机营养盐、陆源有机碳及无机碳等,对河口毗邻海区生物地球化学过程有重要影响[1].Mckee等[2]提出河流主控型边缘海概念,指出这种类型的陆架边缘海具有所有海洋系统中最高的初级生产能力,其碳循环的基本特征表现为海表二氧化碳分压(partial pressure of carbon dioxide,pCO2)下降或是显著的大气CO2汇,海-气CO2通量取决于生物群落中自养和异养生物活动的强弱[3].Chen等[4]指出在大河羽流区,丰富营养盐被浮游植物用来进行光合作用,使得该区甚至是距离河口几百公里远的海区都可能成为CO2汇.
长江位于我国黄、东海交界之处,其季节性径流输入使得邻近海区水文结构、生物活动以及海-气CO2通量呈现明显的季节特征.Tseng等[5]指出CO2的吸收与初级生产有关,特别是在长江冲淡水区,在三峡大坝建成后,长江冲淡水流量减小,富含营养盐的冲淡水区域减小,高初级生产范围也随之减小,可能导致CO2吸收能力下降,甚至会使东海陆架由CO2汇转为源.而后,基于同样的前提,Tseng等[6]利用温度、长江径流量建立模型用以估算33°N以南的整个东海海表pCO2年际与季节变化,指出1998—2011年东海pCO2有上升趋势,这很可能与径流量的下降趋势相关.然而在年际变化上长江径流量并非长江冲淡水扩展的主控因子[7-8],径流量较大时,长江冲淡水扩展面积并非较大,且受台湾暖流的影响,冲淡水北分支在某些年份可能更为偏北,甚至达到35°N[7].此外,长江冲淡水扩展主要影响区域为黄、东海交界海区,东海中外陆架海区几乎不受长江冲淡水的影响,因此东海并不是严格意义上的河流主控型边缘海,用长江径流量估算整个东海碳通量存在一定的不确定性.
研究表明,长江径流量在近60年来整体上变化不大,基本上在892.2亿m3上下波动.而长江营养盐浓度则表现出较大幅度的变化,其中溶解无机氮和溶解无机磷浓度呈显著增加趋势[9].1963—2007年,长江溶解无机氮从22.9,µmol/L增加到122.9,µmol/L,累积增加436.7%,;溶解无机磷从0.12,µmol/L增加到1.58,µmol/L,累积增加1,217%,[10].Gypens等[11]的研究指出,比利时沿海地区在20世纪70年代至80年代随着氮、磷的增加,此海区由大气CO2的源转换成CO2的汇;20世纪90年代末,随着磷的减少,此海区又由CO2的汇变成CO2的源.Chou等[12]指出21世纪初相对于20世纪90年代东海内陆架海水夏季pCO2明显降低,秋冬季海水pCO2明显增加,认为这两个不同年代海水pCO2的变化可能与富营养化有关.长江营养盐浓度的增加对长江口邻近海区海-气CO2通量有什么影响?又是如何影响的?这是本文探讨的主要问题.基于此,本文应用三维的东中国海生态动力-无机碳循环耦合模型,设计敏感性实验,在不考虑营养盐结构变化的前提下,仅使长江营养盐浓度各增加50%,,并将营养盐增加的实验所得结果与参考实验进行比较.
1 模型配置及实验设计
1.1 模型配置
本模型是一个三维的东中国海生态动力-无机碳循环耦合模型,包括3个模块:水动力模块、生态模块和无机碳化学模块.其中,水动力模块为生态模块和无机碳化学模块提供海水的温度、盐度、流速、扩散系数等数据,3个模块同时运行.
水动力模块使用Guo等[13]基于普林斯顿海洋模型(Princeton Ocean Model,POM)[14-15]在东中国海模拟的结果.Guo等[13]采用3层嵌套技术得到了较高分辨率(水平方向1/18°×1/18°,垂向sigma坐标,共21层)的物理场,可以更准确地刻画黄东海的水动力环境.
生态模块是基于挪威生态模型(NORWegian ECO-logical Model,NORWECOM)[16-18]在东中国海进行了重建,并且与Guo等[13]改进的水动力模块耦合[19],即东中国海生态动力模型(East China Sea ECOlogical Model,ECSECOM).Luo等[20]在ECSECOM基础之上加入无机碳循环过程,初步完成了陆架海碳循环模型的建立.该模型包含了黄、东海碳循环的主要控制过程,如海-气界面CO2交换、河流溶解无机碳(dissolved inorganic carbon,DIC)输入、沉积物-水界面颗粒有机碳(particulate organic carbon,POC)的沉积与再悬浮过程以及开边界外海营养盐和DIC的输入.
在无机碳化学模块中,DIC和总碱度(total alkalinity,TA)为预报变量,海-气CO2通量(air-sea CO2,flux,FDIC)和pCO2为诊断变量.海水中DIC和TA的控制方程为
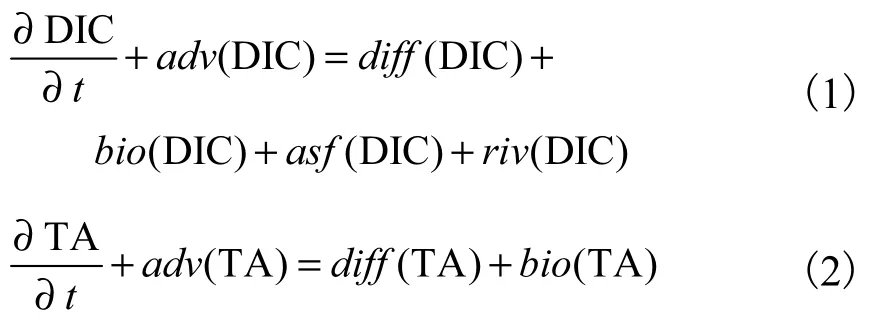
式中:adv、diff、bio、asf、riv分别表示对流项、扩散项、生物过程项、海-气CO2交换过程和河流输入项.
该模型大气强迫条件中,1/4°×1/4°分辨率的气候态月均风场和气压数据来自欧洲中期天气预报再分析数据集(European Centre for Medium-Range Weather Forecasts-Reanalysis DATA Archive,ECMRW-ERA:http://data-portal.ecmwf.int/data/d/ interim_full_monthly/).全球3°×2°月均干大气CO2物质的量比(2000—2010年)来自美国国家海洋与大气管理局(National Oceanic and Atmospheric Administration,NOAA:http://www.esrl.noaa.gov/gmd/ccgg/ carbontracker/download.html).DIC和TA的初始场以及DIC开边界数据来自二氧化碳信息处理中心(Carbon Dioxide Information Analysis Center,CDIAC:http://odv.awi.de/en/data/ocean/global_alkanity_tco2/),该数据是基于全球多航次构建的空间分辨率为1°×1°,时间分为4个季节,垂向标准33层的数据集.长江入海月均DIC通量根据长江年入海DIC总量和长江径流量在季节内的配比计算[21-22].在开阔大洋TA浓度主要受盐度控制,因此开边界TA浓度设置为盐度的函数[23].模型计算时间步长为6,min,模式运行3年达到稳定状态,取第3年的输出结果进行分析.
本文研究范围为25°N ~37°N,118°E ~128°E,水深200,m以浅海区.
1.2 模型验证
Guo等[13]采用三重嵌套POM模型所模拟的海面高度与T/P(TOPEX/Poseidon)卫星反演的海面高度的均方根空间分布一致,并较好地模拟了东中国海黑潮路径;通过与实际调查数据的比较,POM亦可较好地模拟南黄海以及东海北部的底层温度场[24].此外,与POM耦合的东中国海生态动力学模型,能够抓住氮、磷、硅3种无机营养盐以及叶绿素a浓度季节分布的主要特征[19].Luo等[20]在东海选取一适当的子区域将模拟得到的海表DIC浓度与多航次调查数据经空间平均后进行了比较,结果表明实测数据和模拟结果呈现较为一致的季节变化趋势.不同比较表明,水动力环境、营养盐、叶绿素和DIC都得到比较可靠的模拟结果.
模型结果的海表pCO2四季分布(图1)特征表明:长江口外及羽流区pCO2在秋季达最大,约在350~430,µatm,而其他三季pCO2整体小于350,µatm,春季局部海区pCO2小于250,µatm;闽浙沿岸秋季pCO2最高,大于350,µatm,其他季节约在250~350,µatm;东海北部陆架pCO2在夏季最高,秋季次之,然而两季pCO2均大于350,µatm,而冬、春季pCO2则在250~300,µatm;东海中陆架和东海南部海区pCO2在夏季达最大,约在350~430,µatm,其他季节除春季东海最南端pCO2约达400,µatm之外,余下海区pCO2均小于350,µatm.Guo等[25]对东海陆架2006—2011年多航次海表pCO2的调查结果进行统计,结果表明东海冬季pCO2分布较为均匀,整体在330~360,µatm,长江口外春夏季pCO2小于100,µatm,秋季pCO2大于400,µatm,达最大值;闽浙沿岸pCO2亦是在秋季达最大,超过350,µatm,而其他三季则在300~350,µatm;东海北部陆架夏秋季(pCO2>340,µatm)要高于冬春季;东海中陆架和东海南部pCO2在夏季达最大,约在380~400,µatm,其他季节则小于350,µatm.可见,除夏季长江口外pCO2模拟明显偏高外,东海其他海区pCO2季节变化的模拟与Guo等[25]多年观测数据统计结果是比较一致的.

图1 模式数据海表pCO2的季节分布(单位:µatm)Fig. 1Seasonal distribution of surface pCO2of the modeled data(unit:µatm)
1.3 敏感性实验设计
研究表明,长江径流量在近60年来整体上变化不大[10],而长江营养盐的浓度,主要是溶解无机氮和磷酸盐呈显著增加趋势[9].因此,通过设计敏感性实验,模拟长江营养盐增加的情景,探究长江营养盐浓度的增加对邻近海区海-气CO2通量的影响.敏感性实验包括一个参考算例(参考实验)和一个营养盐增加的算例(NUTRINR实验).NUTRINR实验中,在不考虑营养盐结构变化的前提下,仅使长江营养盐(即无机氮、无机磷、硅酸盐)浓度各增加50%,,其他条件与参考实验相同.
2 结果与讨论
2.1 营养盐变化量的分布
将NUTRINR实验结果与参考实验结果作差,得到两组实验各变量的变化量.从营养盐浓度变化量分布来看,长江营养盐浓度的变化对东海中、外陆架大部分海区营养盐浓度分布几乎没有影响.
长江营养盐浓度增加50%,后,NUTRINR实验表层溶解无机氮(NIT)浓度相对于参考实验变化量的分布(图2)显示:1—5月NIT增加的海区主要集中在50,m以浅海区,长江口外、苏北浅滩等30,m以浅近岸海区NIT增加在5,mmol/m3以上,约是参考实验的30%,以上(图2(a)—(e)).6月至夏末,随着长江径流量的增加,营养盐向海通量增加,NIT随长江冲淡水向济州岛方向扩展,50,m以浅海区NIT继续保持着较高浓度;与此同时,受台湾暖流的顶托作用,长江冲淡水羽流南下分支较弱,长江口南部30,m以浅沿岸海区NIT增加不明显(图2(f)—(h)).秋季后,除50,m以浅海区保持高浓度外,NIT浓度明显发生变化的海区范围收缩(图2(i)—(l)).
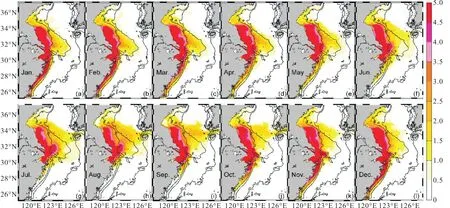
图2 NUTRINR实验表层NIT浓度相对于参考实验的变化量(单位:mmol/m3)Fig. 2 Variation of the surface dissolved inorganic nitrogen concentration between NUTRINR case and normal run (unit:mmol/m3)
NUTRINR实验表层溶解无机磷(PHO)浓度相对于参考实验变化量的分布(图3)显示:PHO增加的海区主要集中在30,m以浅海区,同NIT增加的海区相似,特别是在长江口外和苏北浅滩等海区.
6—9月,PHO增加量相较其他月份并不明显(图3(f)—(i)),结合叶绿素质量浓度变化量分布(图4),表明这种现象主要是由浮游植物的消耗所引起的.春季浮游植物水华首先发生在黄海中部,然而长江营养盐浓度的增加此时并不能影响到该区,因此,早春NUTRINR实验叶绿素质量浓度相对参考实验结果的差异并不明显.至春末及夏季,潮汐锋区以及长江口外出现浮游植物水华,这些海区的NIT、PHO增加,促进了植物光合作用,叶绿素质量浓度增加,由于NIT过量,浮游植物生长受到PHO限制[26],所以在此期间河流输入增加的PHO被浮游植物消耗尽,PHO相对参考实验无明显变化.而在50,m以深海区,尽管NIT增加明显,但受PHO影响范围的限制,叶绿素质量浓度几乎没有变化(图4).总体而言,夏季长江输入营养盐通量随着长江径流量的增加而增加,由于长江冲淡水的输运作用及浮游植物的消耗作用,使得NIT在长江口邻近海区表现为向济州岛方向扩展,而PHO增加却不明显;秋季长江输入营养盐通量逐渐减少,冬季为长江输入营养盐通量最低值,此时NIT扩展范围收缩,集中在50,m以浅海区,浮游植物消耗的降低使PHO能够扩展到30,m以浅海区;从春季开始,长江输入营养盐通量又开始增加,营养盐分布逐渐过渡到夏季的状态.

图3 NUTRINR实验表层溶解PHO浓度相对于参考实验的变化量(单位:mmol/m3)Fig. 3 Variation of the surface dissolved inorganic phosphate concentration between NUTRINR case and normal run (unit:mmol/m3)
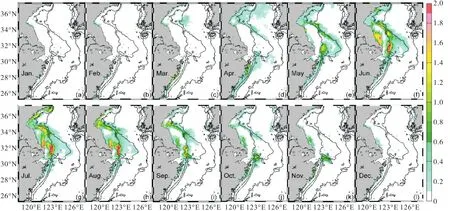
图4 NUTRINR实验表层叶绿素质量浓度相对于参考实验的变化量(单位:mg/m3)Fig. 4 Variation of the surface chlorophyll concentration between NUTRINR case and normal run(unit:mg/m3)
2.2 海-气CO2通量变化量的分布及影响因素
NUTRINR实验相对于参考实验的海-气CO2通量的变化量分布(图5)显示:夏季至中秋,长江口邻近海区FDIC出现负变化,大气向海洋的碳通量增加,并且FDIC负变化范围与冲淡水范围一致(图5(f)—(j));长江口外变化幅度最大,FDIC变化量减少了0.8,mmol/(m2·d),约使FDIC下降了40%,,即碳汇作用增加了40%,.秋季末,苏北浅滩开始出现FDIC正变化海区,表明海洋向大气的碳通量逐渐增加,随时间推移,1月FDIC正变化达最大(图5(a)),此时海洋碳汇作用减弱最多,1月NUTRINR实验FDIC相对参考实验最大增加了0.4,mmol/(m2·d),约使FDIC增加了10%,,即碳汇作用下降了10%,.夏秋季长江输入营养盐通量增加对长江口邻近海区的碳汇变化有正作用,而冬春季长江输入营养盐通量的减少对其邻近海区的碳汇变化有负作用.
海-气交换过程即FDIC依赖于海-气CO2分压差,CO2的溶解度以及海-气界面CO2气体传输速率.

式中:k为CO2气体传输速率,本文采用Wanninkhof[27]提出的与海面10,m风速和水温相关的经验公式来确定k;α 为CO2在海水中的溶解度,本文采用Weiss[28]提出的与海水温盐相关的经验公式确定α ;pCO2a为大气CO2分压;pCO2w为海水的CO2分压.
在NUTRINR实验中,由于物理场相对于参考实验没有变化,所以k、α 以及pCO2a与参考实验是一致的,仅有pCO2w在调控FDIC的变化.

图5 NUTRINR实验海-气CO2通量相对于参考实验的变化量(单位:mmol/(m2·d))Fig. 5 Variation of the air-sea CO2flux between NUTRINR case and normal run(unit:mmol/(m2·d))
夏季,海表pCO2变化量的分布(图6)表明:随着长江营养盐输运的增加,即长江输入营养盐通量增加,长江冲淡水扩展区域内pCO2降低,苏北浅滩和长江口外下降幅度最大,海表pCO2下降量在20,µatm以上;秋季至春季,苏北浅滩以及长江浅滩海表pCO2逐渐增加到10,µatm以上.而pCO2的变化主要受DIC和TA浓度变化的影响.研究表明,全球表层海水DIC平均增加1%,,pCO2将增加10%,, TA增加1%,,pCO2将减少9.4%,[29],这两者对pCO2的贡献是反相的.浮游植物的生长与呼吸在改变DIC浓度的同时,NIT浓度的变化也会影响TA的变化.长江营养盐浓度的增加所刺激的生物活动的变化通过改变DIC和TA对pCO2产生影响.在长江口及其邻近海区,DIC的变化幅度要明显大于TA的变化幅度(图7),表明海表pCO2的变化主要由DIC的变化主导.
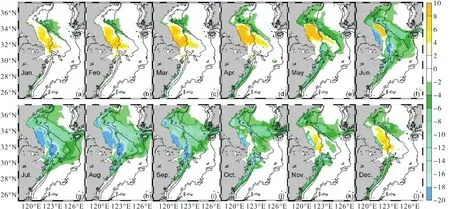
图6 NUTRINR实验表层海水pCO2相对于参考实验的变化量(单位:µatm)Fig. 6 Variation of sea surface pCO2between NUTRINR case and normal run(unit:µatm)
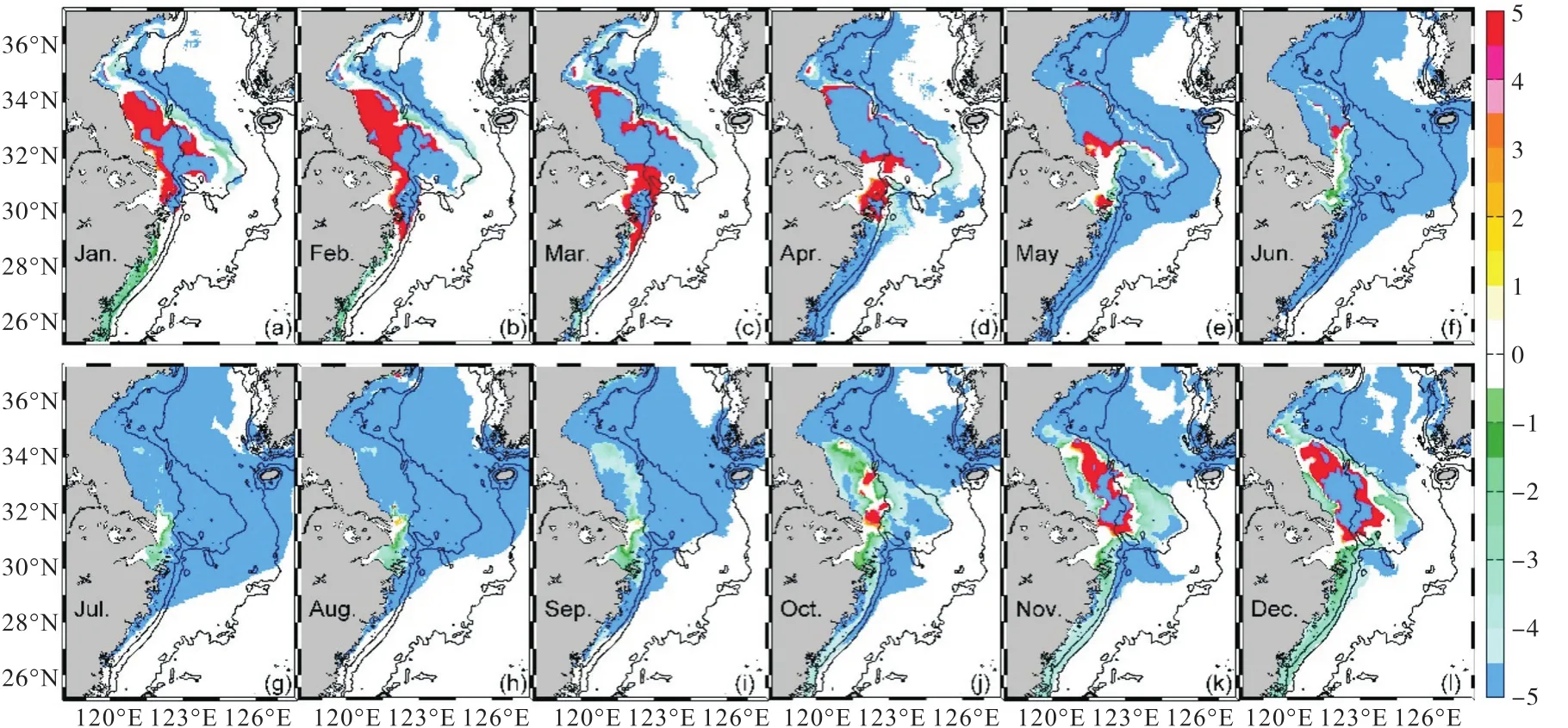
图7 NUTRINR实验表层DIC浓度与TA浓度相对于参考实验变化百分比的比值Fig. 7 Ratio of the percentage variations of the sea surface DIC and TA between NUTRINR case and normal run
NUTRINR实验表层DIC浓度相对于参考实验的变化量分布(图8)显示:夏季,随着长江输入营养盐通量的增加,河口邻近海区浮游植物生长消耗DIC增加,长江口邻近海区的DIC的负变化范围不断扩大,长江冲淡水的对流作用将低DIC水团向济州岛方向输运(图8(f)—(i)),此时受DIC变化的影响,pCO2负变化范围也在向济州岛方向扩展且与DIC变化范围相似(图6(f)—(i)),同时生物作用使得水体中有机碎屑增多(图9(f)—(i)).从秋末开始,长江输入营养盐通量减少,光合作用减弱,有机碎屑的生物降解开始占据生物活动的主体,水体中的有机碎屑含量逐渐减少并转变为DIC(图9(j)—(d)),从而使DIC的浓度不断增加(图8),DIC浓度的增加也使得pCO2出现正变化并逐渐增强的趋势(图6(j)—(d)).秋末至春初,长江口及苏北浅滩海区DIC有所增加,浮游植物光合作用很弱,此时海洋又处于吸收大气CO2的状态,DIC并没有消耗,所以DIC持续保持这一正变化量的状态,直至春末开始出现浮游植物水华,DIC正变化区域开始收缩,至夏季呈现为DIC负变化.整体而言,长江营养盐浓度增加,对DIC的影响主要表现为夏、秋季DIC的生物消耗增加,而冬季至春初有机碎屑的分解增加.
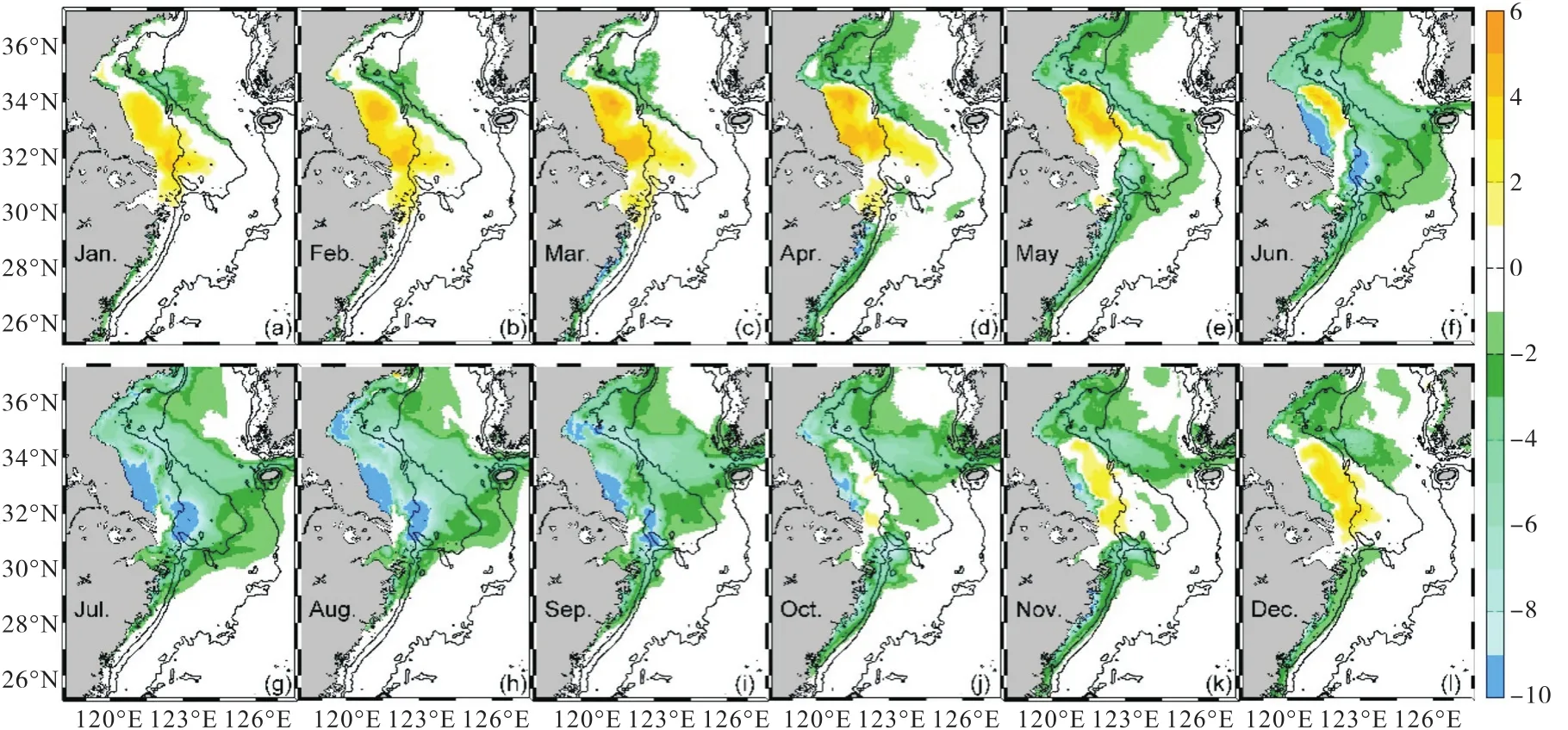
图8 NUTRINR实验表层DIC浓度相对于参考实验的变化量(单位:µmol/kg)Fig. 8 Variation of sea surface DIC concentration between NUTRINR case and normal run(unit:µmol/kg)
本文对长江浅滩(图9(a)所示方框,30.8°N~33°N,122.5°E ~125°E)在不同季节吸收CO2的能力做一个估算,并与参考实验进行比较(表1).长江浅滩在受长江冲淡水影响较明显的海区范围之内,因此可以选取此海区用以评估海表吸收CO2能力的强弱及其变化趋势.对长江浅滩而言,长江营养盐浓度的增加对夏季海-气碳通量的影响最为明显,夏季大气CO2向海通量增加,海表可多吸收约68%,的大气CO2;对秋季影响次之,长江营养盐的增加使长江浅滩秋季碳源作用减弱38%,;冬季和春季,大气CO2向海通量减小,即碳汇作用有所减弱;从全年看,长江营养盐增加50%,后,长江浅滩约多吸收6%,的大气CO2.
Chou等[30]对冬季观测数据总结指出20世纪90年代至21世纪初,东海碳汇作用在减弱,并认为这主要由夏季不断增加的有机碳分解所致.陈鑫等[31]对近年来夏季已有观测数据总结指出,长江羽流区夏季碳汇作用呈增加趋势,认为是夏季长江平均月径流量的增加造成冲淡水面积不断扩大所致,但是Wei等[7]根据实测数据发现在年际变化上长江径流量并非长江冲淡水扩展的主控因子,周锋等[8]通过模拟长江径流量极枯年2006年与极丰年1999年长江冲淡水扩展的差异指出径流量并非冲淡水扩展的主导因素.所以这种碳汇作用变化趋势可能就是对长江营养浓度不断增加的这种长期变化的响应,而非长江径流量的变化所导致的.
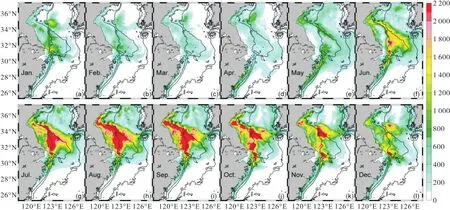
图9 NUTRINR实验有机碎屑浓度(以碳计)的水柱积分相对于参考实验的变化量(单位:mg/m2)Fig. 9 Variation of the column burden of detritus concentration(count by carbon)between NUTRINR case and normal run (unit:mg/m2)

表1 参考实验与NUTRINR实验中长江浅滩海-气CO2通量季节分布Tab. 1 Seasonal distribution of the air-sea CO2flux between the normal case and NUTRINR run about Changjiang Bank
3 结 语
本文基于三维的东中国海生态动力-无机碳循环耦合模型,根据长江营养盐浓度的变化,设计敏感性实验,在不考虑营养盐结构变化的前提下,仅使长江营养盐浓度各增加50%,,研究其引起的长江口及其邻近海区海-气CO2通量的变化.与参考实验相比,各营养盐的变化量分布如下:NIT增加主要分布在50,m以浅海区,特别是长江口外、苏北浅滩等地.PHO增加主要分布在30,m以浅海区,6—9月由于浮游植物的消耗表现出PHO增加不明显的特征.
长江营养盐的增加所致的生物效应主控长江口邻近海区DIC的变化,进而对海-气CO2通量产生影响.夏季浮游植物光合作用消耗DIC,使DIC浓度减小,此时水体中累积的生物有机碎屑在秋、冬季分解又使得水体DIC浓度增加.DIC比TA的变化幅度大很多,因此DIC的变化主导了pCO2的变化,而在NUTRINR实验中,物理场没有改变,海表pCO2的变化决定了FDIC的变化.夏季至中秋,长江口邻近海区海-气CO2通量为负变化,大气向海洋的碳通量增加;秋末到春初,苏北浅滩海-气CO2通量为正变化,海洋向大气的碳通量增加.长江浅滩在夏季碳汇作用增强,约多吸收68%,的大气CO2,而在冬春季则碳汇作用减弱;全年尺度上,长江浅滩海区海水吸收CO2的能力增强约6%,.在观测数据中总结出的长江口邻近海域冬季碳汇作用减弱,夏季碳汇增强的这一变化趋势很可能就是对长江营养盐浓度增加的响应.此外,初级生产的增强将导致底层有机碳的累积增多,推测其与黄东海颗粒有机碳的沉积埋藏及海洋酸化现象存在一定关系,这需要结合沉积物-水界面碳交换过程的研究作进一步的分析讨论.
[1] Chen C T A,Zhai W,Dai M. Riverine input and air-sea CO2exchanges near the Changjiang(Yangtze River) Estuary:Status quo and implication on possible future changes in metabolic status[J]. Continental Shelf Research,2008,28(12):1476-1482.
[2] McKee B A,Aller R C,Allison M A,et al. Transport and transformation of dissolved and particulate materials on continental margins influenced by major rivers:Benthic boundary layer and seabed processes[J]. Continental Shelf Research,2004,24(7):899-926.
[3] Dai M,Cao Z,Guo X,et al. Why are some marginal seas sources of atmospheric CO2?[J]. Geophysical Research Letters,2013,40(10):2154-2158.
[4] Chen C T A,Huang T H,Fu Y H,et al. Strong sources of CO2in upper estuaries become sinks of CO2in large river plumes[J]. Current Opinion in Environmental Sustainability,2012,4(2):179-185.
[5] Tseng C M,Liu K K,Gong G C,et al. CO2uptake in the East China Sea relying on Changjiang runoff is prone to change[J]. Geophysical Research Letters,2011,38(24):L24609. 1- L24609. 6.
[6] Tseng C M,Shen P Y,Liu K K. Synthesis of observed air-sea CO2exchange fluxes in the river-dominated East China Sea and improved estimates of annual and seasonal net mean fluxes[J]. Biogeosciences,2014,11(14):3855-3870.
[7] Wei H,Luo X F,Zhao Y,et al. Intraseasonal variation in the salinity of the Yellow and East China Seas in the summers of 2011,2012,and 2013[J]. Hydrobiologia,2015,754(1):13-28.
[8] 周锋,宣基亮,倪晓波,等. 1999年与2006年间夏季长江冲淡水变化动力因素的初步分析[J]. 海洋学报,2009,31(4):1-12.
[9] Wang B. Cultural eutrophication in the Changjiang (Yangtze River)plume:History and perspective[J]. Estuarine,Coastal and Shelf Science,2006,69(3):471-477.
[10] 辛明. 长江口海域关键环境因子的长期变化及其生态效应[D]. 青岛:中国海洋大学,2014.
[11] Gypens N,Borges A V,Lancelot C. Effect of eutrophication on air-sea CO2fluxes in the coastal Southern North Sea:A model study of the past 50 years [J]. Global Change Biology,2009,15(4):1040-1056.
[12] Chou W C,Gong G C,Cai W J,et al. Seasonality of CO2in coastal oceans altered by increasing anthropogenic nutrient delivery from large rivers:Evidence from the Changjiang-East China Sea system[J]. Biogeosciences,2013,10(6):3889-3899.
[13] Guo X,Hukuda H,Miyazawa Y,et al. A triply nested ocean model for simulating the Kuroshio-Roles of horizontal resolution on JEBAR[J]. Journal of Physical Oceanography,2003,33(1):146-169.
[14] Mellor G L. Users Guide for a Three Dimensional,Primitive Equation,Numerical Ocean Model[M]. Princeton:Princeton University,1998.
[15] Blumberg A F,Mellor G L. A description of a threedimensional coastal ocean circulation model[M]//Heaps N S. Three-dimensional Coastal Ocean Models. Washington:American Geophysical Union,1987:1-16.
[16] Aksnes D L,Ulvestad K B,Baliño B M,et al. Ecological modelling in coastal waters:Towards predictive physicalchemical-biological simulation models[J]. Ophelia,1995,41(1):5-36.
[17] Skogen M D,Svendsen E,Berntsen J,et al. Modelling the primary production in the North Sea using a coupled three-dimensional physical-chemical-biological ocean model[J]. Estuarine,Coastal and Shelf Science,1995,41(5):545-565.
[18] Skogen M D,Søiland H. A User's Guide to NORVECOM V2.0. The Norwegian Ecological Model System[M]. Bergen:Institute of Marine Research,1998.
[19] Zhao L,Guo X. Influence of cross-shelf water transport on nutrients and phytoplankton in the East China Sea:A model study[J]. Ocean Science,2011,7(1):27-43.
[20] Luo X F,Wei H,Liu Z,et al. Seasonal variability of airsea CO2fluxes in the Yellow and East China Seas:A case study of continental shelf sea carbon cycle model[J]. Continental Shelf Research,2015,107:69-78.
[21] 方娟娟,李义天,孙昭华,等. 长江大通站径流量变化特征分析[J]. 水电能源科学,2011,29(5):9-12.
[22] 李丹. 中国东部若干入海河流水化学特征与入海通量研究[D]. 上海:华东师范大学,2009.
[23] Christian J R,Feely R A,Ishii M,et al. Testing an ocean carbon model with observed sea surface pCO2and dissolved inorganic carbon in the tropical Pacific Ocean [J]. Journal of Geophysical Research:Oceans,2008,113(C7):C07047.1-C07041.17.
[24] Wei H,Deng L,Wang Y,et al. Giant jellyfish Nemopilema nomurai gathering in the Yellow Sea:A numerical study[J]. Journal of Marine Systems,2015,144:107-116.
[25] Guo X H,Zhai W D,Dai M H,et al. Air-sea CO2fluxes in the East China Sea based on multiple-year underway observations[J]. Biogeosciences,2015,12(18):5495-5514.
[26] Wang B D,Wang X L,Zhan R. Nutrient conditions in the Yellow Sea and the East China Sea[J]. Estuarine,Coastal and Shelf Science,2003,58(1):127-136.
[27] Wanninkhof R. Relationship between wind speed and gas exchange over the ocean[J]. Journal of Geophysical Research:Oceans,1992,97(C5):7373-7382.
[28] Weiss R F. Carbon dioxide in water and seawater:The solubility of a non-ideal gas[J]. Marine Chemistry,1974,2(3):203-215.
[29] 乔然,王彰贵,张滨,等. 海洋中的CO2观测与研究[J]. 海洋预报,2005,22(增刊):106-114.
[30] Chou W C,Gong G C,Tseng C M,et al. The carbonate system in the East China Sea in winter[J]. Marine Chemistry,2011,123(1):44-55.
[31] 陈鑫,宋金明,袁华茂,等. 东海2012年夏季海-气界面碳交换及其区域碳汇强度变化趋势初探[J]. 海洋学报,2014,36(12):18-31.
责任编辑:周建军
Influence of Increasing Nutrients of the Changjiang River on Air-sea CO2Flux in Adjacent Seas
LIU Zhe1,LUO Xiaofan1,WEI Hao2,ZHAO Liang1
(1.College of Marine and Environmental Sciences,Tianjin University of Science & Technology,Tianjin 300457,China;2.School of Marine Science and Technology,Tianjin University,Tianjin 300072,China)
In recent 50 years,the nutrient concentration of the dissolved inorganic nitrogen phosphorus in Changjiang River continued to increase,which has already had certain impact on the ecosystem of Changjiang River estuary and the adjacent seas and also caused disturbances of the air-sea CO2flux.Based on the three-dimensional ecological dynamic-inorganic carbon cycle coupling model of the East China Sea,sensitivity experiments were designed without considering structural changes in nutrients and assuming that each nutrient concentration of Changjiang River increased by 50%,,which simulated the increase of the nutrients in Changjiang River,so as to explore the influence of increasing nutrients on air-sea CO2flux in adjacent seas of Changjiang estuary.The results show that compared with the reference case,the increased nutrient concentration of Changjiang River has brought about a positive change in the atmospheric CO2seaward flux about Changjiang Bank.In summer,with the carbon sink strengthened,the sea surface absorbed more atmospheric CO2,about 68%,.In winter,the seaward flux of the atmospheric CO2decreased above Changjiang Bank and Subei Shoal.That is to say,carbon sink was weakened in winter.The CO2absorption capacity has had an increase of about 6%,anmually above Changjiang Bank.The biological effect,due to the increased nutrient concentration of Changjiang River,is the major cause leading to the changes of dissolved inorganic carbon in adjacent seas,which further influenced the air-sea CO2flux.
air-sea CO2flux;nutrient;Changjiang estuary;ecological model
P734.4
A
1672-6510(2016)06-0055-09
10.13364/j.issn.1672-6510.20150270
2015-12-30;
2016-03-05
国家自然科学基金资助项目(41376112,41276016);中国科学院战略性先导科技专项(A类)资助项目(XDA11020305)
刘 哲(1989—),男,山东滨州人,硕士研究生;
赵 亮,教授,zhaoliang@tust.edu.cn.
数字出版日期:2016−07−11;数字出版网址:http://www.cnki.net/kcms/detail/12.1355.N.20160711.1608.002.html.
