孕穗开花期持续低温对不同熟期水稻气孔导度的影响*
2016-12-27马熙达任传友王艳华徐一丹赵东妮
马熙达,任传友**,王艳华**,徐一丹,赵东妮,陈 伟,2,杨 斌,田 平
(1.沈阳农业大学农学院大气科学系,沈阳 110866;2.辽宁省北票市气象局,北票 122100)
孕穗开花期持续低温对不同熟期水稻气孔导度的影响*
马熙达1,任传友1**,王艳华1**,徐一丹1,赵东妮1,陈 伟1,2,杨 斌1,田 平1
(1.沈阳农业大学农学院大气科学系,沈阳 110866;2.辽宁省北票市气象局,北票 122100)
分别以 9023、9035、9036为早熟、中熟、晚熟水稻的代表品种,应用人工气候箱在孕穗开花期进行低温处理,旨在分析沈阳地区孕穗开花期不同强度低温对不同熟期水稻叶片气孔导度的影响。低温设置以前一天相同时刻的温度值为基准,设比外界温度低3℃(D3)和低5℃(D5)两个处理水平,持续处理5d,以自然条件下生长的水稻为对照(CK),利用LI-6400光合测定仪测定水稻倒2叶的CO2响应曲线和光响应曲线。结果表明:(1)CO2响应过程中,与CK相比,D3和D5的持续低温处理皆可导致9035和9036的气孔导度显著降低,而9023(早熟)仅D5的持续低温导致其气孔导度降低;(2)光响应过程中,与CK相比,D3和D5的持续低温处理使9035(中熟)和9036(晚熟)气孔导度降低,且降低幅度随低温强度的增强而增大,但持续低温处理显著增加了 9023(早熟)的气孔导度(P<0.01);(3)气孔限制是引起 9035(中熟)遭遇低温后光合速率下降的主要原因,而 9023(早熟)和 9036(晚熟)光合速率的变化主要由非气孔限制导致。说明中熟(9035)和晚熟(9036)水稻品种对低温处理响应较敏感,而早熟(9023)品种对低温不敏感。
孕穗开花期;持续低温;气孔导度;气孔限制
水稻是中国种植面积最广、产量最高的粮食作物之一,同时,水稻亦为喜温性作物,其生长发育和产量形成对低温条件反应敏感[1],且越晚熟的品种对热量的要求越高,遭遇低温冷害导致产量减少的幅度越大。低温冷害对水稻生产的影响一直是国内外学者关注的重点[2-5]。
植物叶片的气孔是水汽和 CO2进出的门户,气孔在控制碳的吸收和水分损失的平衡中起着关键作用,影响着植物的光合、呼吸、蒸腾等生理机能[6-7],且它随着所处环境状况的不同而发生变化,了解气孔对环境变量的响应规律,可加深对植物光合作用限制因素的理解。气孔导度是衡量气孔开张程度的度量指标,其值的大小直接决定了水汽和 CO2进出气孔的量的多少,因此,研究气孔导度值大小是探讨植物叶片水分蒸散和 CO2同化速率变化的一个关键环节。国内外学者对水稻生理指标的变化规律及其对环境因子的响应关系等进行了大量研究,如高温[8-9]、灌溉[10-11]、盐胁迫[12-13]、低温[14-15]等对水稻生理过程的影响,但以往研究主要侧重于单一熟期品种生理指标的变化,且主要集中于环境因素对光合、蒸腾的影响分析,同一地区不同熟期水稻品种遭遇低温后气孔导度的对比研究则鲜见报道。
东北地区地处中国北方,热量资源不足,低温冷害是该区水稻生产中面临的主要农业气象灾害之一。近年来,东北地区作物种植结构和品种布局有了较大调整,偏晚熟水稻品种不断向北部和东部扩展[16]。由于受水稻耐冷性不同以及地域气候差异的影响,不同熟期水稻的气孔行为对低温的响应也并不相同。东北地区近年冷害的增加除了与气候异常事件频繁有关外,也与盲目栽种偏晚熟品种有关,对比研究不同熟期水稻遭遇低温后气孔导度的变化规律,有助于加强对遭遇低温后不同熟期水稻光合作用限制因素的了解,为合理布局不同熟期品种、降低冷害风险提供理论依据。
研究表明,水稻在孕穗开花期对低温最敏感[17-19]。本文选取9023、9035和9036水稻品种作为沈阳地区早、中、晚熟水稻的代表品种,对比研究其在孕穗开花期不同低温下气孔导度的变化规律及光合过程的气孔限制因素,以及加强对沈阳地区不同熟期水稻对低温的响应规律的理解,为气候变化情景下不同熟期水稻在特定地区的适应性及田间管理、高效生产提供理论依据。
1 材料与方法
1.1 试验设计
试验于2011年在沈阳农业大学实验田进行,选用不同熟期代表品种 9023(早熟)、9035(中熟)、9036(晚熟)为试验材料,采用半盆栽半大田的栽培方式。盆钵为PP长方体塑料桶,钵口长24cm,宽17cm,钵底长18cm,宽12cm,钵高26cm。钵内先套上纱网袋,再将干土8.5kg装入纱网袋中,每钵施2.0g尿素、2.0g磷酸二铵作为底肥,4月18日播种,5月22日移栽到盆钵中,每钵1穴,每穴2株。5月29日待秧苗成活后将沙网袋连同秧苗从盆钵中取出,移栽到大田统一管理。在进入水稻孕穗开花盛期时再将沙网袋连同植株从大田中取出直接放回盆钵中,将盆钵置于人工气候箱内进行低温处理,9023、9035、9036进入开花盛期的时间分别为7月28日、8月17日、8月25日。
人工气候箱设置24波段,每个波段1h,用以体现气象要素的日变化规律,临近日出和日落的两个波段的间距可能不为1h,采用天文学方法确定日出、日落时间进行适当调整。各波段相对湿度、光照与中国天气网发布的实时数据一致。温度设置两个低温水平,以前一天相同时刻的温度值为基准,分别比外界低3℃和5℃,并以D3和D5表示。每个品种、每个低温处理放入气候箱内3穴,持续处理5d,以自然条件下正常生长的水稻为对照CK。
1.2 观测项目
将进入开花盛期的水稻置于人工气候箱进行低温处理,低温处理结束后置于大田恢复 1d,各组均选3穴,每穴选一株使用LI-6400便携式光合测定仪测定倒2叶的CO2响应曲线和光响应曲线,3个重复,CO2响应曲线在光合有效辐射(PAR)为1200µmol·m-2·s-1下进行测定,共设置11个CO2浓度水平,分别为800、600、400、300、200、150、100、80、60、50、40µmol·mol-1;光响应曲线在CO2浓度为400µmol·mol-1下进行测定,共设置15个PAR水平,分别为 1800、1500、1200、1000、800、600、400、200、100、80、60、40、20、10、0µmol·m-2·s-1。
1.3 计算指标
气孔导度模型是评价植物叶片气孔调节的重要工具[20-22]。本文选用王建林等[23]提出的双曲线方程来表述不同处理不同熟期气孔导度对叶表面 CO2浓度的响应特征,表达式为

式中,gmax为潜在最大气孔导度(mol·m2·s-1),Cs为叶片表面CO2浓度(µmol·mol-1),Cso为试验常数(µmol·mol-1)。
气孔的开闭和光合作用的光反应具有一致性,本研究采用光合作用速率-光响应的双曲线形式来表述气孔导度对光强的响应特征[22]。表达式为
高铁对珲春市旅游业产生了巨大影响,长珲高铁于2015年9月20日正式开通运营,对比2014年至2017年黄金周珲春市重点景区旅游数据可以看到,高铁对珲春市旅游业的巨大推动作用。

式中,gmax为最大气孔导度(mol·m-2·s-1),PARc为最大气孔导度 1/2时的光合有效辐射(µmol·m-2·s-1);gct为角质层导度(mol·m-2·s-1)。
植物的光合作用过程受外界环境条件和植物自身状况的共同影响。气孔开度的大小决定了外界CO2是否能顺畅地进入气孔内部,从而也就决定了参与光合作用过程的 CO2含量及浓度的高低。因此,气孔行为是制约光合作用强度的最重要的因素之一。通常植物在受到胁迫条件下,气孔导度会降低,使得进入气孔内部的 CO2减少,不能满足光合作用的需求,导致光合速率下降,称为光合作用的气孔限制。判断气孔限制值的两个重要因素是Ci和Ls[24],其中气孔限制值Ls表示为[25-27]

式中,Ci为胞间CO2浓度(µmol·mol-1),Ca为大气CO2浓度(µmol·mol-1)。
1.4 数据处理
采用SPSS统计软件进行参数拟合和统计分析,并采用均方根误差RMSE验证由式(1)和式(2)拟合气孔导度值和实际测量气孔导度值的接近程度,即

式中,di和si分别为实测值和模拟值,n为比较数据的对数。
2 结果与分析
2.1 低温条件下各品种叶片气孔导度对CO2响应的比较
由图 1可见,饱和光强下(PAR为 1200 mol·m-2·s-1),各品种水稻在不同条件下(CK、D3、D5)所测叶片气孔导度均随着CO2浓度的增加而减小,表现出相同的变化趋势,但是,同等条件下其数值大小、变幅和速率有一定区别。具体从 3个品种的比较看,相同条件下 9023(早熟)品种的气孔导度数值最大,在0.222~0.731mol·m-2·s-1,9035(中熟)品种最小,在0~0.173mol·m-2·s-1,9036(晚熟)品种居中,在 0.198~0.452mol·m-2·s-1。可见,叶片气孔导度数值大小与品种早晚熟期之间无明显的对应关系,为品种间的个体差异。对于同一品种来说,低温处理组(D3、D5)与对照组(CK)叶片气孔导度CO2响应曲线有一定差别,总体上看,9035、9036即中、晚熟品种中低温处理后叶片气孔导度 CO2响应曲线的下降幅度和速率较大(比CK)(图1b、c),说明这两个中、晚熟品种对低温处理的敏感性相对较高,而 9023(早熟)品种对低温处理的敏感性不高,D3处理曲线甚至在CK之上(图1a)。

图1 饱和光强下(PAR=1200µmol·m-2·s-1)不同熟期水稻叶片气孔导度的CO2响应曲线Fig.1 Stomatal conductance of different maturities in CO2response curves under saturation intensity(PAR=1200µmol·m-2·s-1)
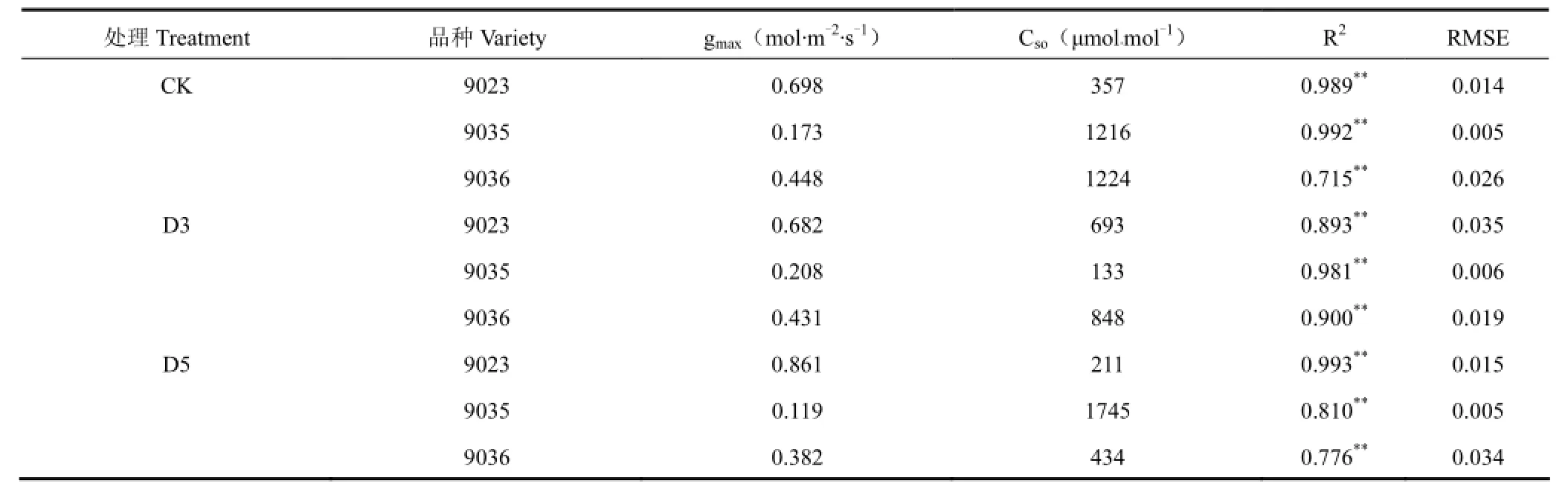
表1 不同熟期叶片气孔导度的CO2响应曲线特征参数Table 1 Parameters of stomatal conductance in CO2response curve with different maturities
2.2 低温条件下各品种叶片气孔导度对PAR响应的比较
由图2可见,CO2浓度为400µmol·mol-1时,不同熟期水稻在各处理下(CK、D3、D5)其叶片气孔导度随着PAR的变化呈相同趋势,即随着PAR的增加气孔导度逐渐增加。但是,不同熟期品种之间气孔导度数值、变幅和速率均存在差异。相同处理条件下,9023(早熟)气孔导度最大,为0.08~0.48mol·m-2·s-1;9035(中熟)最小,为0.02~0.20mol·m-2·s-1;9036为0.03~0.50mol·m-2·s-1。同熟期品种处理组和对照组叶片气孔导度光响应曲线存在一定差异,9035(中熟)和9036(晚熟)低温处理后,叶片的气孔导度均呈下降趋势,且随着低温强度的增加,气孔导度值下降幅度和速率增大(图2b、c)。说明9035(中熟)和9036(晚熟)对低温敏感性较高,而9023(早熟)品种D3和D5处理组气孔导度光响应曲线在对照组(CK)之上(图2a),可见适度的低温可增大9023的气孔导度。
以式(2)对不同持续低温处理下气孔导度对PAR的响应进行模拟,结果见表2。从表2可以看出,各处理拟合程度均达极显著水平。持续低温对不同熟期水稻光响应过程最大气孔导度的影响与对CO2过程的影响明显不同。相同处理条件下,9035(中熟)潜在最大气孔导度值最小,为0.067~0.234mol·m-2·s-1,D3处理组和CK对照组中9036(晚熟)的潜在最大气孔导度最大,为 0.534~0.933mol·m-2·s-1,而D5处理组中9023(早熟)的潜在最大气孔导度最大,为0.454mol·m-2·s-1。随着低温强度的增加,早熟品种9023和晚熟品种9036光响应过程的潜在最大气孔导度皆呈显著下降趋势,且晚熟品种 9036降幅更大,相对于CK,9036的D3和D5处理组的最大气孔导度分别下降了46.2%和61.2%。

图2 不同熟期水稻叶片气孔导度的光响应曲线Fig.2 Stomatal conductance of different maturities in light response curves

表2 不同熟期叶片气孔导度的光响应曲线特征参数Table 2 Parameters of stomatal conductance in light response curve with different maturities
2.3 低温条件下各品种叶片气孔限制值的比较
自然条件下,环境CO2浓度虽并非定值,但其变化幅度较 PAR小得多。因此,探讨光响应过程叶片碳同化的影响及制约因素更具实际意义。本文即从光响应过程来探讨不同熟期水稻在孕穗开花期遭遇持续低温后,气孔导度对光合作用的影响。植物的光合作用受气孔和非气孔两个因素的制约,气孔限制分析的方法已得到广泛应用。根据许大全[24]的研究,Ci和 Ls的变化方向是判断气孔因素和非气孔因素降低叶片光合速率的重要判据:Ci降低和Ls升高表明气孔导度降低是主要原因;而Ci增高和Ls降低则表明主要原因是非气孔因素。由图3可见,不同熟期品种随着低温处理强度的增加,Ls和Ci随着PAR的变化呈现不同的变化趋势。相同PAR水平下,9023(早熟)和 9036(晚熟)Ls值随着低温强度的增加逐渐降低(CK>D3>D5),Ci逐渐升高(CK

图3 不同低温处理下不同熟期Ci与Ls随PAR的变化Fig.3 Changes of Ci(intercellular CO2concentration)and Ls(stomata limitation)of different maturities with PAR (photosynthetically active radiation) after different low temperature treatments
3 结论与讨论
3.1 讨论
气孔对植物光合作用和蒸腾作用的平衡具有调节作用,在植物适应环境变化和环境压力中起着关键作用[28]。关于气孔导度对环境响应的生理过程已展开大量研究。唐凤德等[29]发现红叶松叶片气孔导度与温度呈正相关,而在本研究中不同熟期品种的气孔导度对温度的响应并非全部呈正相关变化,低温均降低了9035(中熟)和9036(晚熟)气孔导度,且随着低温处理强度增加气孔导度持续下降,中熟和晚熟品种气孔导度与温度呈正相关变化,而对于9023(早熟)CO2响应过程中的D3处理组和光响应过程中D3、D5处理组气孔导度值均在CK组之上,在一定低温范围内,早熟品种气孔导度与温度呈负相关,这可能与不同熟期水稻耐冷性不同有关。本研究中早熟品种9023和晚熟品种9036在孕穗开花期遭遇不同强度持续低温后,表现出气孔限制值 Ls随着低温强度的增加而下降,而胞间CO2浓度Ci值增加,表明非气孔限制是导致光合速率变化的主要原因,这一结论与李健陵等[14]对早晚兼种型水稻的研究结果一致。
在当前气候变暖背景下,中国东北地区水稻延迟性冷害减少[16],但障碍型冷害呈增多趋势[30]。障碍型冷害的增多除了与气候本身的波动增大有关外,不科学地引种偏晚熟品种也在一定程度上起了推波助澜的作用。明确不同熟期水稻气孔导度对持续低温的响应规律,将有助于加强对不同熟期水稻光合碳固定过程冷害机制的综合理解,这对于在气候变暖情景下科学布局不同熟期水稻品种、降低气候风险具有重要的指导作用。
本研究以9023、9035、9036为沈阳地区早、中和晚熟水稻的代表品种,评价了不同熟期水稻气孔导度对持续低温的响应规律。沈阳地区目前栽种的水稻品种较多,本研究每个熟期类型仅选取一个品种作为代表,具有一定的局限性。即使是熟期相同的水稻,由于其对低温的耐受性不同,气孔导度对低温的响应规律必然存在差异,能反应该区不同熟期水稻对持续低温响应的普适性规律的获得还需对各个熟期水稻更多品种的进一步验证。
水稻是喜温作物,东北地区热量资源不足是限制该区水稻生长发育及产量形成的关键环境限制因子之一。除孕穗开花期外,水稻在分蘖期、拔节期、灌浆期对低温也较敏感,分析评价不同熟期水稻在不同生育期对持续低温的响应规律,量化持续低温对不同熟期水稻产量的影响,对科学布局水稻生产具有重要的指导作用。加强不同熟期水稻在各生育期对持续低温响应规律的对比研究,更科学布局水稻生产,主动适应气候变化,已成为该区水稻生产中迫切的现实需求,也是摆在科学工作者面前的重要研究课题。
3.2 结论
(1)水稻孕穗开花期遭遇不同强度的持续低温,9035(中熟)和 9036(晚熟)品种气孔导度显著降低,随着低温处理强度的增加气孔导度值持续下降,低温对水稻叶片气孔导度影响越大。对于 9023(早熟),适当的低温反而增加CO2响应过程和光响应过程气孔导度。
(2)低温导致光合速率变化的主要原因也体现了品种之间的差异,气孔导度降低是引起9035(中熟)光合速率下降的主要原因,而 9023(早熟)和 9036(晚熟)光合速率的变化主要由非气孔限制导致。
(3)不同熟期水稻叶片气孔导度对低温的响应不同,表明 9023(早熟)对低温敏感性较小,9035(中熟)和 9036(晚熟)较 9023(早熟)耐低温能力弱。
References
[1]马树庆,王琪,王春乙,等.东北地区水稻冷害气候风险度和经济脆弱度及其分区研究[J].地理研究,2011,30(5):931-938.Ma S Q,Wang Q,Wang C Y,et al.Climate risk and economy vulnerability of rice chilling damage and division in Northeast China[J].Geographcal Research, 2011,30(5):931-938.(in Chinese)
[2]胡春丽,李辑,林蓉,等.东北水稻障碍型低温冷害变化特征及其与关键生育期温度的关系[J].中国农业气象,2014, 35(3):323-329.Hu C L,Li J,Lin R,et al.Characters of sterile-type chilling damage in rice and its relationship with temperature at key growth stage in Northeast China[J].Chinese Journal of Agrometeorology, 2014,35(3): 323-329.(in Chinese)
[3]王冬妮,郭春明,刘实,等.吉林省水稻延迟型低温冷害气候风险评价与区划[J].气象与环境学报,2013,29(1):103-107.Wang D N,Guo C M,Liu S,et al.Evaluation and division on climatic risk of rice delayed cool damage in Jilin province[J].Journal of Meteorology and Environment, 2013, 29(1): 103-107.(in Chinese)
[4]朱海霞,王秋京,闫平,等.孕穗抽穗期低温处理对黑龙江省主栽水稻品种结实率的影响[J].中国农业气象,2012, 33(2):304-309.Zhu H X,Wang Q J,Yan P,et al.Effect of low temperature at heading stage on seed setting rate of major rice varieties in Heilongjiang Province[J].Chinese Journal of Agrometeorology, 2012,33(2): 304-309.(inChinese)
[5]宋广树,孙忠富,孙蕾,等.东北中部地区水稻不同生育时期低温处理下生理变化及耐冷性比较[J].生态学报,2011, 31(13):3788-3795.Song G S,Sun Z F,Sun L,et al.Comparison between physiological properties and cold tolerance under low temperature treatment during different growing stages of rice in northeast central region of China[J].Acta Ecologica Sinica, 2011, 31(13): 3788- 3795.(in Chinese)
[6]Warren C R,Dreyer E.Temperature response of photosynthesis and internal conductance to CO2:results from two independent approaches[J].Journal of Experimental Botany,2006,57 (12): 3057-3067.
[7]高春娟,夏晓剑,师恺,等.植物气孔对全球环境变化的响应及其调控防御机制[J].植物生理学报,2012,48(1):19-28.Gao C J,Xia X J,Shi K,et al.Response of stomata to global climate changes and the underlying regulation mechanism of stress responses[J].Plant Physiology Journal,2012,48 (1): 19-28.(in Chinese)
[8]张玉屏,朱德峰,林贤青,等.高温对水稻剑叶生长和气孔导度影响[J].江西农业大学学报,2012,34(1):1-4.Zhang Y P,Zhu D F,Lin X Q,et al.Effect of high temperature stress on leaf growth and stomatal conductance in rice[J].Acta Agriculturae Universitatis Jiangxiensis,2012,34(1): 1-4.(in Chinese)
[9]谢晓金,李秉柏,李映雪,等.抽穗期高温胁迫对水稻产量构成要素和品质的影响[J].中国农业气象,2010,31(3):411-415.Xie X J,Li B B,Li Y X,et al.Effects of high temperature stress on yield components and grain quality during heading stage[J].Chinese Journal of Agrometeorology, 2010, 31(3): 411-415.(in Chinese)
[10]彭世彰,徐俊增,丁加丽.控制灌溉水稻气孔导度变化规律试验研究[J].农业工程学报,2005,21(3):1-5.Peng S Z,Xu J Z,Ding J L.Variations of stomatal conductance of rice under contolled irrigation[J].Transactions of the CSAE,2005,21(3):1-5.(in Chinese)
[11]赵黎明,李明,郑殿峰,等.灌溉方式与种植密度对寒地水稻产量及光合物质生产特性的影响[J].农业工程学报, 2015,31(6):159-169.Zhao L M,Li M,Zheng D F,et al.Effects of irrigation methods and rice planting densities on yield and photosynthetic characteristics of matter production in cold area[J].Transactions of the CSAE,2015, 31(6):159-169.(in Chinese)
[12]徐晨,凌风楼,徐克章,等.盐胁迫对不同水稻品种光合特性和生理生化特性的影响[J].中国水稻科学,2013, 27(3):280-286.Xu C,Ling F L,Xu K Z.et al.Effect of salt stress on photosynthetic characteristics and physiological and biochemical traits of different rice varieties[J].Chin J Rice Sci, 2013, 27(3):280- 286.(in Chinese)
[13]刘晓龙,徐晨,徐克章,等.盐胁迫对水稻叶片光合作用和叶绿素荧光特性的影响[J].作物杂志,2014,25(2):88-92.Liu X L,Xu C,Xu K Z,et al.Effects on characteristics of photosynthesis and chlorophyll fluorescence of rice under salt stress[J].Crops, 2014,25(2):88-92.(in Chinese)
[14]李健陵,霍治国,吴丽姬,等.孕穗期低温对水稻产量的影响及其生理机制[J].中国水稻科学,2014,28(3):277-288.Li J L,Huo Z G,Wu L J,et al.Effects of low temperature on grain yield of rice and its physiological mechanism at the booting stage[J].Chin J Rice Sci,2014, 28(3):277-288.(in Chinese)
[15]李霞,戴传超,程睿,等.不同生育期水稻耐冷性的鉴定及耐冷性差异的生理机制[J].作物学报,2006,32(1):76-83.Li X,Dai C C,Cheng R,et al.Identification for cold tolerance at different growth stages in rice (Oryza sativa L.)and physiological mechanism of differential cold tolerance[J].Acta Agronomica Sinica,2006,32(1): 76-83.(in Chinese)
[16]方修琦,王媛,朱晓禧.气候变暖的适应行为与黑龙江省夏季低温冷害的变化[J].地理研究,2005,24(5):664-672.Fang X Q,Wang Y,Zhu X X.Change of cool summer hazard under an adaptation behavior to the climate warming in Heilongjiang Province,Northeast China[J].Geographical Research, 2005, 24(5): 664-672.(in Chinese)
[17]曲辉辉,姜丽霞,朱海霞,等.孕穗期低温对黑龙江省主栽水稻品种空壳率的影响[J].生态学杂志,2011,30(3):489-493.Qu H H,Jiang L X,Zhu H X,et al.Effects of low temperature at booting stage on the percentage of unfilled grains of major rice varieties in Heilongjiang Province[J].Chinese Journal of Ecology,2011,30 (3): 489-493.(in Chinese)
[18]邓化冰,车芳璐,肖应辉,等.开花期低温胁迫对水稻花粉性状及剑叶理化特性的影响[J].应用生态学报,2011,22(1):66-72.Deng H B,Che F L,Xiao Y H,et al.Effects of low temperature stress during flowering period on pollen characters and flag leaf physiological and biochemical characteristics of rice[J].Chinese Journal of Applied Ecology, 2011,22(1):66-72.(in Chinese)
[19]姜丽霞,李帅,闫平,等.黑龙江水稻孕穗期障碍型冷害及其对产量的影响[J].中国农业气象,2009,30(3):463-468.Jiang L X,Li S,Yan P,et al.Sterile type cool injury of rice during booting stage and its impacts on rice yield in Heilongjiang province[J].Chinese Journal of Agrometeorology, 2009, 30(3): 463-468.(in Chinese)
[20]王海珍,韩路,徐雅丽,等.不同温度下灰胡杨叶片气孔导度对光强响应的模型分析[J].生态环境学报,2015,24(5):741-748.Wang H Z,Han L,Xu Y L,et al.Model analysis of the stomatal conductance response to light in populus pruinosa at different temperatures in the Taklimakan Desert[J].Ecology and Environmental Sciences,2015,24(5): 741-748.(in Chinese)
[21]李永秀,娄运生,张富存.冬小麦气孔导度模型的比较[J].中国农业气象,2011,32(1):106-110.Li Y X,Lou Y S,Zhang F C.Comparison of stomatal conductance models for winter wheat[J].Chinese Journal of Agrometeorology, 2011,32(1): 106-110.(in Chinese)
[22]王建林,于贵瑞,王伯伦,等.北方粳稻光合速率、气孔导度对光强和CO2浓度的响应[J].植物生态学报,2005,29(1):16-25.Wang J L,Yu G R,Wang B L,et al.Response of photosynthetic rate and stomatal conductance of rice to light intensity and CO2concentration in Northern China[J].Acta Phytoecologica Sinica, 2005, 29(1):16-25.(in Chinese)
[23]王建林,温学发.气孔导度对CO2浓度变化的模拟及其生理机制[J].生态学报,2010,30(17):4815-4820.Wang J L,Wen X F.Modeling the response of stomatal conductance to variable CO2concentration and its physiological mechanism[J].Acta Ecologica Sinica, 2010, 30(17): 4815-4820.(in Chinese)
[24]许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯,1997,33(4):241-244.Xu D Q.Some problems in stomatal limitation analysis of photosynthesis[J].Plant Physiology Communications, 1997,33 (4): 241-244.(in Chinese)
[25]丁雷,李英瑞,李勇,等.梯度干旱胁迫对水稻叶片光合和水分状况的影响[J].中国水稻科学,2014,28(1):65-70.Ding L,Li Y R,Li Y,et al.Effects of drought stress on photosynthesis and water status of rice leaves[J].Chin J Rice Sci,2014,28(1):65-70.(in Chinese)
[26]王春艳,庞艳梅,李茂松,等.干旱胁迫对大豆气孔特征和光合参数的影响[J].中国农业科技导报,2013,15(1):109-115.Wang C Y,Pang Y M,Li M S,et al.Effects of drought stress on soybean stomatal characteristics and photosynthetic parameter[J].Journal of Agricultural Science and Technology, 2013,15(1):109-115.(in Chinese)
[27]杨泽粟,张强,郝小翠.自然条件下半干旱雨养春小麦生育后期旗叶光合的气孔和非气孔限制[J].中国生态农业学报,2015,23(2):174-182.Yang Z S,Zhang Q,Hao X C.Stomatal or non-stomatal limitation of photosynthesis of spring wheat flag leaf at late growth stages under natural conditions in semiarid rainfed regions[J].Chinese Journal of Eco-Agriculture, 2015,23(2): 174-182.(in Chinese)
[28]鱼腾飞,冯起,司建华.极端干旱区多枝柽柳叶片气孔导度的环境响应模拟[J].植物生态学报,2012,36(6):483-490.Yu T F,Feng Q,Si J H.Simulating responses of leaf stomatal conductance to environmental factors for Tamarix ramosissma in an extreme arid region of China[J].Chinese Journal of Plant Ecology,2012, 36(6):483-490.(in Chinese)
[29]唐凤德,武耀祥,韩士杰,等.长白山阔叶红松林叶片气孔导度与环境因子的关系[J].生态学报,2008,28(11):5649-5655.Tang F D,Wu Y X,Han S J,et al.Relationship of stamatal conductance of leaf with environmental factors in broadleaved Korean pine forest at Changbai Mountain[J].Acta EcologicaSinica, 2008,28(11): 5649-5655.(inChinese)
[30]王绍武,马树庆,陈莉,等.低温冷害[M].北京:气象出版社,2009:23-40.Wang S W,Ma S Q,Chen L,et al.Low temperature and cold damage[M].Beijing: China MeteorologicalPress, 2009:23-40.(inChinese)
Effects of Consecutive Low Temperature on Stomatal Conductance of Rice with Different Maturity Periods at Booting and Blooming Stages in Shenyang Region
MA Xi-da1, REN Chuan-you1, WANG Yan-hua1, XU Yi-dan1, ZHAO Dong-ni1, CHEN Wei1,2, YANG Bin1, TIAN Ping1
(1.College of Agronomy, Shenyang Agriculture University, Shenyang 110866,China;2.Beipiao Meteorological Bureau, Beipiao 122100)
The experiment was conducted in climate chamber to explore the effects of different intensities of low temperature on stomatal conductance of rice with different maturity at booting and blooming stages in Shenyang region.9023, 9035, 9036 were chosen as representative early, middle and late maturity varieties, respectively.Two temperature treatments were stetted for 5 days, which were 3℃(D3) and 5℃(D5) lower than outside temperature of the last day, CO2and light response curves of second leaf from top were measured for treatment and control plants (grown under natural conditions as control, mark CK) by using LI-6400 photosynthesis analyzer.The results showed that:(1) during CO2response, the leaf stomatal conductance of D3 and D5 treatment plants showed a significant decrease compared with that of CK plants for 9035(middle) and 9036(late) maturity varieties, while decrease for 9023(early) maturity variety only in D5 treatment.(2) During light response, the leaf stomatal conductance of treatment plants showed the similar variation with the results of (1) compared with that of CK plants for 9035(middle) and 9036(late) varieties, but increase for 9023(early) maturity variety in D3 and D5 treatment(P<0.01).(3) Stomatal limitation mechanism is the main mechanism that leads to decrease of photosynthetic induced by low temperature for 9035(middle) variety, whereas, the decline in photosynthetic rate is mainly due to non-stomatal limitation for 9023(early) and 9036(late) maturity varieties.9035(middle) and 9036(late) varieties are more sensitive than 9023(early) maturity variety to low temperature.
Booting and blooming stage; Consecutive low temperature; Stomatal conductance; Stomatal limitation
2016-04-11**
国家自然科学基金(31201124);公益性行业(农业)科研专项经费项目(201503116;21303130)
马熙达(1990-)女,硕士生,主要从事农业气象灾害研究。E-mail:maxd90@126.com
10.3969/j.issn.1000-6362.2016.06.008
马熙达,任传友,王艳华,等.孕穗开花期持续低温对不同熟期水稻气孔导度的影响[J].中国农业气象,2016,37(6):682-690
