不同温度对钩栗光响应曲线拟合的影响
2016-12-21罗小浪
张 斌,罗小浪
(中南林业科技大学 林学院,湖南 长沙 410004)
不同温度对钩栗光响应曲线拟合的影响
张 斌,罗小浪
(中南林业科技大学 林学院,湖南 长沙 410004)
以珍贵用材树种钩栗2年生幼苗为研究对象,采用LI-6400光合速率测试仪对钩栗成熟叶片的光合能力进行测定分析,在对钩栗25 ℃和45 ℃条件下的光合特征曲线进行拟合后发现5个模型间存在不同程度的差异。高温胁迫提高了钩栗对弱光的利用能力,也显著降低了钩栗的最大净光合能力;从模型的拟合效果来看,直角双曲线修正模型在常温时的模拟值与实际值和其他模型相比高出12.99%~30.57%,而在高温条件下却高出12.44%~26.91%,拟合效果最差;常温条件下指数模型、指数修正模型和非直角双曲线模型具有较好的拟合效果,但在高温条件下,非直角双曲线模型和指数修正模型的效果最好,综合强光照条件下钩栗的光合规律,非直角双曲线模型综合表现最佳。
光响应曲线;光响应模型;钩栗;光合生理
光是调控植物生长的重要因子,植物通过改变叶片的解剖结构和生理生化特性来维持较强的光合能力[1-2],植物的光合生理对环境胁迫特别是温度的变化尤为敏感,研究树种生境中温度等逆境胁迫对光合作用的影响及其与植物的生态分布和演替状态之间联系多年来一直倍受关注[3-6]。因此开展珍贵用材树种钩栗的不同温度胁迫下光能利用参数研究,对于钩栗速丰性的前期鉴定和制定科学的栽培管理措施具有重要的意义[7-8]。植物的生长速度受净光合速率(Pn)的影响,影响净光合速率的因素有CO2浓度、光照强度、温湿度等指标,由于光照强度对净光合速率的影响最大,为了更准确的了解净光合速率与光照强度的关系,前人建立了许多光响应曲线的数学模型,并利用这些模型能准确而快速的模拟出所研究对象的光合生理特性和相关光合生理参数[9-13],如最大净光合速率Pnmax、光补偿点Lcp、光饱和点Lsp等,由于模型本身的局限性,针对不同的树种需要选择合适的模型并进行适当的参数修正,以达到更好的模拟树种真实光合特征的需求。
钩栗CastanopsistibetanaHance,又名钩栲,隶属壳斗科栲属,常绿大乔木,亚热带地区优良的用材树种,亦是具有开发价值的园林绿化树种[14]。综合分析钩栗的研究现状,多数研究停留在种群育苗实验和生命过程等方面,对钩栗生理特性如蒸腾特性、光合特性及环境因子对这些生理反应的影响等方面的研究尚未见很多报道[15-18]。本研究通过对钩栗光响应曲线的分析和拟合,旨在掌握钩栗在不同温度条件下对光照辐射的适应性和需光特征,研究结果可进一步丰富钩栗培育技术和理论。
1 试验材料与研究方法
1.1 试验材料
2015年5月中旬,选择生长基本一致,苗高65~70 cm,地径2.2~2.5 cm,叶片未受损伤且生长健壮的2年生钩栗幼苗60株,在中南林业科技大学森林培育实验室利用光照培养箱培养(培养温度为25 ℃、45 ℃,土壤含水量控制在田间持水量的80%左右,光照时间12 h,光照强度1 000 mol·m-2s-1左右)15 d后,进行光响应曲线测试。
1.2 研究方法
1.2.1 光源使用LI-6400
光合速率仪的02B红蓝光源,光照强度在0 ~ 2 000 μmol·m-2s-1范围按照 0、25、50、100、200、400、600、800、1 000、1 200、1 400、1 600、1 800、2 000 μmol·m-2s-1的梯度变化[14],间隔时间2 min。测定时叶片温度分别为(25±1) ℃和(45±1) ℃,空气相对湿度为(80±1)%,光合有效辐射为 (1000±5) μmol·m-2s-1。根据瞬时光照强度和对应的净光合速率,并结合不同的光响应曲线模型,绘制光合速率曲线并计算出光补偿点等钩栗光合生理参数。
1.2.2 光响应曲线模型
目前,光响应曲线很多[19],本研究选择5种常见的模型方程来拟合钩栗的光合特征曲线(表1)。
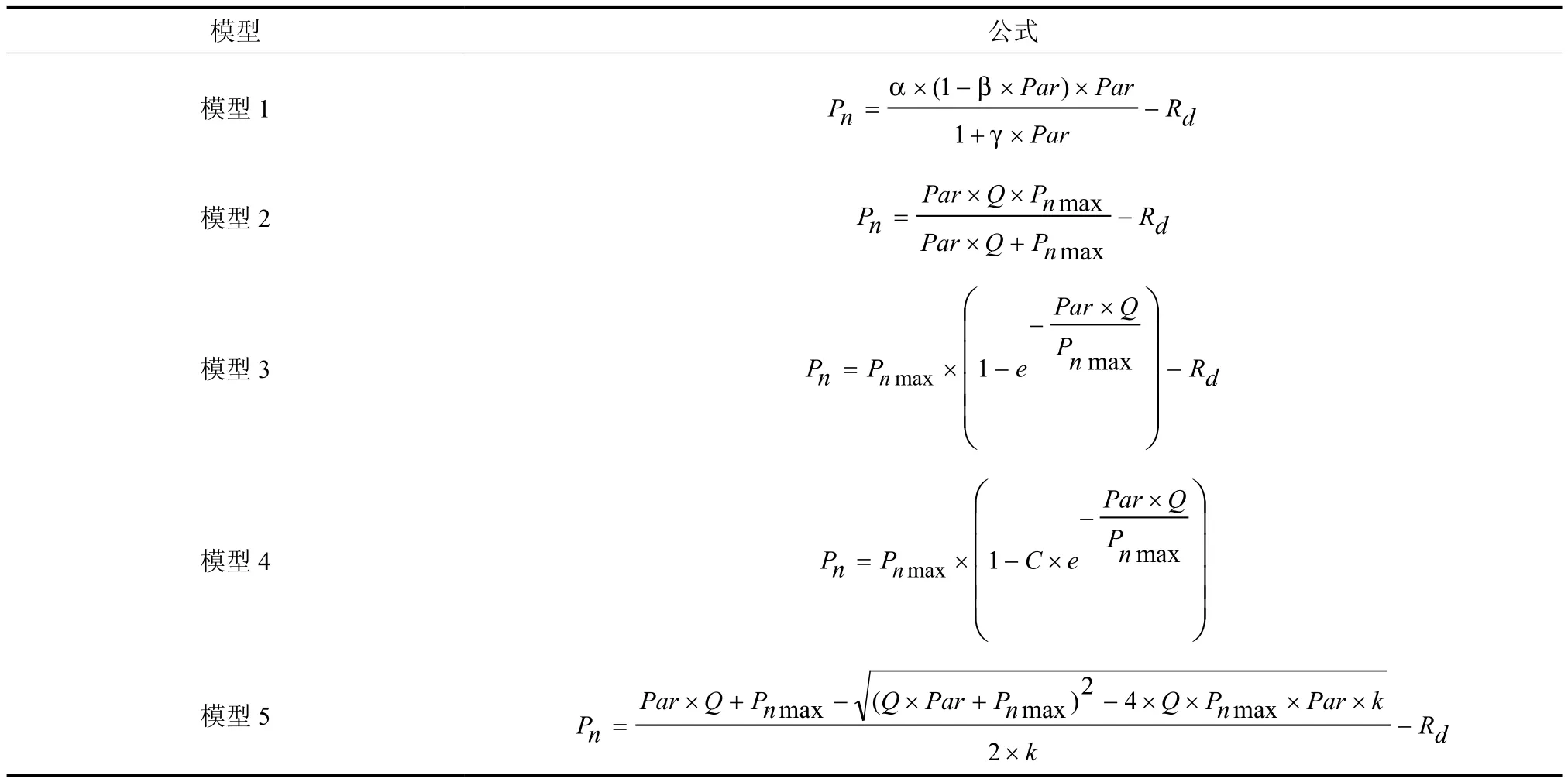
表1 光响应模型†Table 1 Light responsive curve models
1.2.3 数据分析
运用Excel进行数据统计和处理,采用SPSS软件的非线性回归功能模块对光响应曲线的数据进行拟合,并进行方差分析和相关性分析。在数据数据拟合过程中通过反复对相关参数的微调,来减少模型参数的误差。
2 结果与分析
2.1 不同温度下光响应测定结果
光响应曲线的测定在生长季节的5月中旬进行,以25 ℃和45 ℃两个温度梯度下生长7 d后的钩栗幼苗为研究对象,测定其在不同温度条件下的光响应曲线(图1),从图中可以得出:在高温条件下,钩栗的光合作用启动早,有光辐射时即可进行光合作用,而常温条件下当光照辐射高于25 μmol·m-2s-1时才启动光合作用,随着光照辐射强度的增加,净光合速率快速增加;当光照辐射大于100 μmol·m-2s-1时,光合作用增幅下降,但以高温条件时下降明显,当达到光饱和后,两种温度下钩栗的光合速率均趋于稳定,但常温条件下的光合速率高于高温条件,分别比45 ℃条件下高出9.48%~12.19%,但差异不显著,出现这种现象的原因可能是高温对植物的叶片组织造成了一定程度的伤害,以至于净光合速率下降。
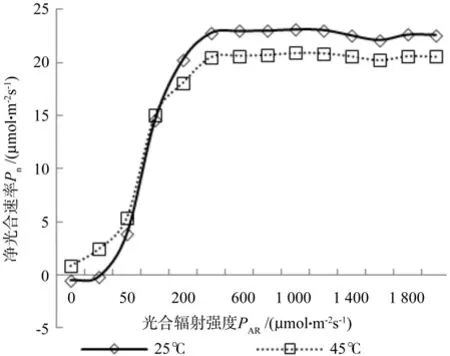
图1 25 ℃、45 ℃时钩栗的光响应曲线Fig.1 Light responsive curve under 25 ℃ and 45 ℃

图2 25 ℃时钩栗的光响应曲线模型Fig. 2 Light responsive curve models under 25 ℃
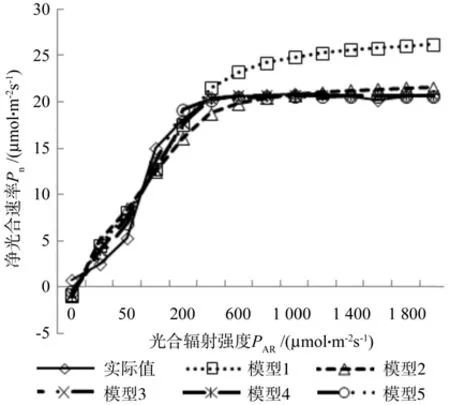
图3 45 ℃时钩栗的光响应曲线模型Fig. 3 Light responsive curve models under 45 ℃
植物接受光信号刺激后开始进行光合作用,随着光照强度的增强,光合产物也越来越多,当钩栗合成和消耗有机物质的速率相等时,此时的光照辐射强度即为钩栗在该条件下的光补偿点(Lcp),一般来说,光补偿点越低,说明钩栗对低强度光的利用能力越强;如果此时外界光照辐射进一步增强,钩栗的光合速率会进一步增大,当光照强度超过某一值时,钩栗的净光合速率达到饱和,此时的光照强度值称为光饱点(Lsp)。Lcp和Lsp代表了植物利用外界光照进行生理物质合成反应所要求的上限和下限,从另外一个方面也反应了该植物对强度光照辐射和弱度光照辐射的利用能力[20-21]。
2.2 不同温度下光响应曲线拟合
图2和图3分别是常温条件和高温条件下钩栗光响应曲线的实测值和5组不同模型值,从图中可以看出在光照辐射较低的情况下,不同模型与实测值接近,当光照辐射高于600 μmol·m-2s-1时,模型1的数值继续缓慢增加,而其他模型和实测值的数据基本稳定,模型1的数值明显高于其他4个模型,其中在25 ℃条件下,模型1值比模型2值高出18.94%~24.17%,比模型3值高出12.99%~29.67%,比模型4值高出12.98%~29.67%,比模型5值高出14.18%~30.57%;而在45 ℃条件下,模型1值比模型2值高出17.23%~21.51%,比模型3值高出12.44%~26.35%,比模型4值高出12.44%~26.35%,比模型5值高出13.71%~26.91%。
同实测值相比,在光照辐射为600~1 200 μmol·m-2s-1时,25℃和 45 ℃条件下均表现为模型3和模型4最优,其次为模型5,25 ℃条件下模型值3、4比实测值高出0.25%~0.74%,模型5比实测值高1.60%~1.75%,45 ℃条件下,模型3、4比实测值高-0.14%~ 0.70%,模型5比实测值高0.92%~1.61%。而当光照辐射大于1 200 μmol·m-2s-1时,均表现为模型5最优,其中在25 ℃条件下,模型5的值仅比实测值低0.69%~2.88%,在45 ℃条件下,模型5的值比实测值低0.15%~1.99%。
2.3 不同温度下光响应模型间差异性分析
通过分析测量实际值和模型拟合的数值之间的差异,采用LSD法进行多重比较,分别得到常温下(表2左下部数值)和高温下(表2右上部数值)的多种比较结果。
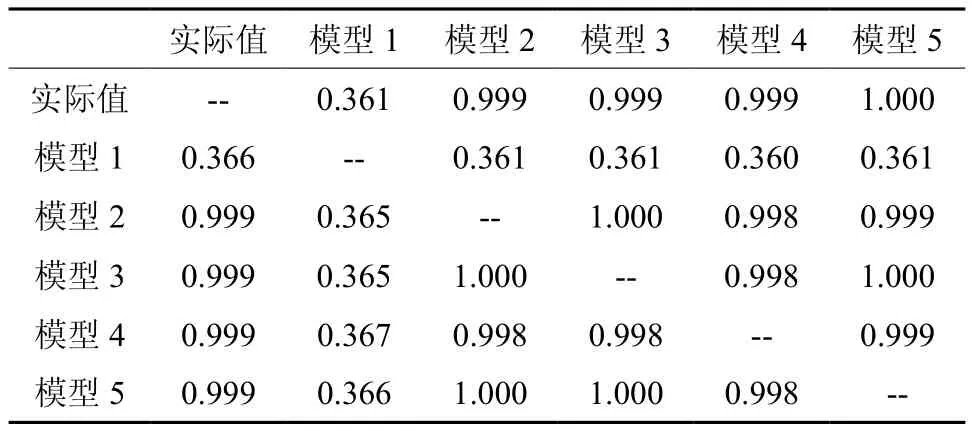
表2 25 ℃和45 ℃下不同模型间多重比较Table 2 LSD analysis of different models under 25 ℃ and 45 ℃
由表2可以看出:不管是在常温下还是在高温下,不同模型值与实测值之间均无显著性的差异,模型1的拟合效果差于其他4个模型,模型1与实测值存在比较大的差异,而其他4个模型相对于模型1来说要好很多;从模型3、4、5之间的多重比较结果来分析,其相关性在0.998~1.000之间,说明这3个模型之间几乎一致。
2.4 光响应曲线特征参数值的比较
根据叶片光响应曲线的5个数学模型,分别计算了钩栗光合作用时的光补偿点(Lcp)、光饱和点(Lsp)、最大净光合速率(Pnmax)、表观量子效率(Q)和呼吸速率(RD)及决定系数(R2),其中Lcp和Lsp是常用的光响应特征参数(表3)。由图2、3的光响应曲线可知,常温下(25 ℃)和高温(45 ℃)下随着光合有效辐射的增大,净光合速率不断增大,钩栗的光合能力不断提升,但增加幅度大小不一(图1~3),钩栗幼苗的光合作用也可能存在光合午休现象[22]。
由表3可以看出,常温(25 ℃)时钩栗的光补偿点(Lcp)、光饱和点(Lsp)均大于高温(45 ℃)条件时,这说明在常温时钩栗利用弱光的能力比在高温时弱,但能更有效地利用强光辐射;同时从最大净光合速率(Pnmax)来看,在常温条件下,钩栗的最大光合速率高于高温逆境下生长的幼苗,说明钩栗在未经过高温胁迫时,具有较快的生长速率[23]。
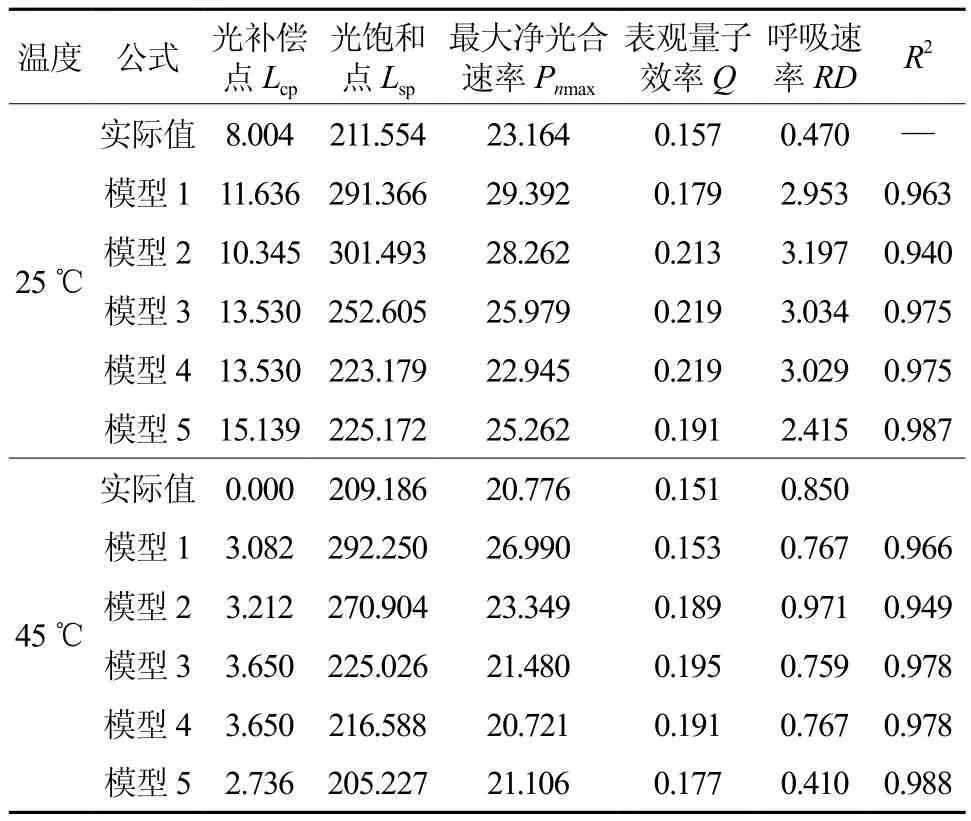
表3 实测值及5种模型光响应曲线特征参数值Table 3 Characters value of 5 different models and measured value
不同温度条件下的Lcp差异极显著(显著性水平<0.01),常温条件下各模型的Lcp分别比实际值高29.25%~89.14%,模型2、1的Lcp与实际值最接近,而高温条件下各模型的Lcp略高于实际值,模型5的Lcp与实际值最接近,其次为模型1、2。
不同温度条件下的Lsp差异不显著(显著性水平0.517),高温对钩栗的Lsp影响不大,常温条件下各模型的Lsp分别比实际值高5.50%~42.51%,高温条件下各模型的Lsp比实际值高-1.89%~ 39.7%,其中均以模型4、5的值与实际值最接近,表现最好,模型1、2的Lsp均表现最高,模型3表现居中。
不同温度条件下的Pnmax差异显著(显著性水平0.041),常温条件的Pnmax比高温条件下高出11.49%,说明高温胁迫降低了钩栗的光合能力,常温条件下各模型的Pnmax分别比实际值高-0.95%~26.89%,高温条件下各模型的Pnmax分别别实际值高-0.27%~29.91%,其中也均以模型4、5的值与实际值最接近,模型1、2均具有最高的Pnmax。
表观量子效率(Q)常温时整体均大于高温时的值,对于任何一种环境,如果植物具有较高的Lsp、Pnmax、Q和较低的Lcp,可预测该植物具有更大的适应性[24],这就表明了钩栗在常温时对光照的适应性广,具有较强的强光利用潜力和较高的光合速率。
5个不同光响应模型对钩栗的光响应曲线拟合精度较高,决定系数在0.940~0.988之间(表3),特别是模型5拟合的决定系数在常温下和高温下时都在0.98以上,说明模型5的拟合效果比较好。模型2在两个温度调节下的拟合精度都最低(R2<0.95);其他模型的拟合效果居中。由于决定系数是模型准确性的指标之一[25],进一步通过对模型的呼吸速率、最大光合速率、光饱和前的线性趋势的比较,来判断哪种模型更适合钩栗的光响应特性。
2.5 光饱和前的线性趋势
从表3看出在常温和高温时钩栗的光饱和点分别是 211.554 μmol·m-2s-1和 209.186 μmol·m-2s-1,都要小于400 μmol·m-2s-1,所以作出了实际值和5个模型拟合值在光合有效辐射为400 μmol·m-2s-1之前的线性趋势图。由图4和图5可以看出:模型2不管是在常温下还是高温下,其线性趋势图均与实际值图相隔距离最大,而模型5则刚好相反,它与实际值图最接近,模型1、3、4则比较均匀地分布在实际值图的两侧,但效果都不如模型5好。
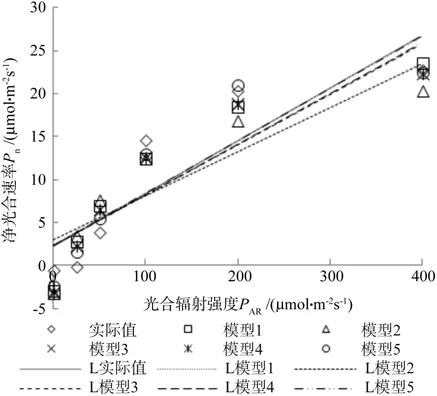
图4 25 ℃时光饱和前的线性趋势Fig. 4 Light curve of different models before light saturation point under 25 ℃
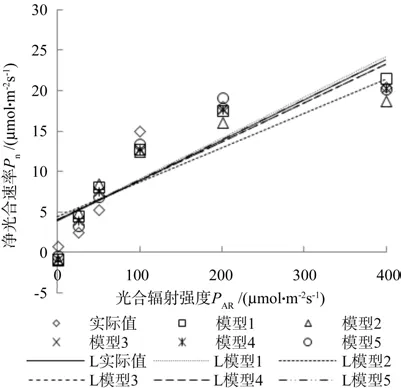
图5 45 ℃时光饱和前的线性趋势Fig.5 Light curve of different models before light saturation point under 45 ℃
3 结论与讨论
本研究采用不同的光响应曲线模型对钩栗在常温(25 ℃)和高温(45 ℃)时光响应进行拟合研究,结果表明温度对钩栗的光合特征参数如光饱和点、补偿点、最大净光合速率、表观量子效率等存在不同程度的影响。
(1)钩栗的光合过程除受叶片自身光合能力的影响,温度胁迫对钩栗的光合也产生了影响,由于光合作用器官对温度条件敏感,在高温和强光条件下,当光照辐射相同时,钩栗的光合能力较常温条件下显著降低,说明高温对钩栗的光合器官产生了较大的影响,进而影响到光合速率的大小,大苞景天在高温条件下也表现出类似的趋势[26]。
(2)植物光补偿点(Lcp)和光饱和点(Lsp)的大小是衡量其耐荫性的重要指标[27],从不同温度条件下的Lcp、Lsp及其差值来看,高温胁迫虽然降低了钩栗的光补偿点,提高了钩栗适应低光照的能力,同时也降低了钩栗光合作用对强光辐射的需求,促使钩栗在较窄的光照范围内就能达到最大的光合速率,这是植物对外界高温胁迫的适应性反应[27-29],但高温胁迫也显著降低了钩栗的最大净光合能力,其原因与高温导致银杏光系统II活性的下降相同[30]。
(3)温度胁迫对各模型的拟合精度影响不大,由于自然界中当阳光直射地面时的光照辐射常大于50 μmol·m-2s-1,因此光响应曲线模型的优劣可以从光饱和点和最大净光合速率的值来判断。在常温条件下指数模型、指数修正模型和非直角双曲线模型具有较好的拟合效果,其次为直角双曲线修正模型和直角双曲线模型;而在高温胁迫条件下,从模型的Lsp、Pnmax及其与实测值的差异来比较,直角双曲线修正模型明显高于实测值和其他模型值,综合表现最差,其次为直角双曲线模型,说明高温胁迫降低了这两个模型的拟合精度,而指数修正模型和非直角双曲线模型在高温胁迫条件下的拟合精度均较常温下增加。根据测定结果,每年5~10月份的钩栗生长旺盛季节,白天的平均辐射强度在1 000 μmol·m-2s-1以上,结合非直角双曲线模型在高辐射条件下具有较高的稳定性即不受温度影响的特点,综合分析表明非直角双曲线模型最优,其次为指数修正模型。
对比其他学者的相关研究发现,直角双曲线修正模型在小叶杨、黄枝油杉的拟合效果优于其他模型[31-32],在拟合栓皮栎和欧洲鹅耳枥的光响应规律中,非直角双曲线模型的效果仅次于直角双曲线模型[33-34]。本研究选择的5个光曲线模型与他人研究不尽相同,研究的结论也不同,其原因可能与钩栗自身的光合生理特性有关,由此可见,在使用方程来拟合不同植物光响应特征时存在差异,同种植物不同部位应该也是存在不同的,其根本原因还需要深入探讨。
[1]蒋高明. 植物生理生态学[M]. 北京: 高等教育出版社, 2004:46- 53.
[2]刘旭峰, 樊秀芳. 猕猴桃幼树光合特性的研究[J]. 园艺学报,1993, 20(4): 329-333.
[3]蔺 琛, 马钦彦, 韩海荣, 等. 山西太岳山辽东栎的光合特性[J]. 生态学报, 2002, 22(9): 1399-1406.
[4]芦站根, 赵昌琼, 韩 英, 等. 不同光照条件下生长的曼地亚红豆杉光合特性的比较研究[J]. 西南师范大学学报, 2003,28(1):117- 121.
[5]杜 虎, 曾馥平, 王克林, 等. 中国南方3种主要人工林生物量和生产力的动态变化[J]. 生态学报, 2014, 38(10):2712-2724.
[6]Xu Kui, Juneau, Philippe. Different physiological and photosynthetic responses of three cyanobacterial strains to light and zinc[J]. Aquatic Toxicology, 2016, 170: 251-258.
[7]王金名. 树木引种驯化概论[M]. 南京:江苏科学技术出版社,1990.
[8]丁希泉, 郑秀梅编著. 农业实用回归分析[M]. 吉林:吉林科学技术出版社, 1989.
[9]陈卫英, 陈真勇, 罗辅燕, 等. 光响应曲线的指数改进模型与常用模型比较[J]. 植物生态学报. 2012, 36(12):1277-1285.
[10]王海珍, 韩 路, 徐雅丽, 等. 不同温度下灰胡杨叶片气孔导度对光强响应的模型分析[J]. 生态环境学报, 2015, 24(5):741-748.
[11]王欢利, 曹福亮, 刘新亮. 高温胁迫下不同叶色银杏嫁接苗光响应曲线的拟合[J]. 南京林业大学学报(自然科学版), 2015,39(2):14-20.
[12]施 曼, 程龙霞, 祝遵凌. 欧洲鹅耳枥光响应曲线模型拟合与应用[J]. 福建林学院学报, 2014, 34(4):349-355.
[13]Higuchi Y, Sumitomo K, Oda A,et al. Day light quality affects the night-break response in the short-day plant chrysanthemum,suggesting differential phytochrome-mediated regulation of fl owering[J]. Journal of Plant Physiology, 2012, 169(18):1789-1796.
[14]祁承经, 汤庚国. 树木学(南方本)[M]. 北京:中国林业出版社, 2010:224-225.
[15]王佩兰, 许德琼, 张 斌, 等. 钩栗种子形态特征及主要生理指标测定分析[J]. 种子, 2013, 32(8):7-11.
[16]王佩兰, 许德琼, 张 斌, 等. 钩栗种源间幼苗生长状况和光合特性比较研究[J]. 北方园艺, 2014(5): 20-25.
[17]林 敏, 黄宗安. 钩栗种群生命表分析[J]. 福建林业科技,2003, 30(2):9-13.
[18]黄荣林, 王 勇, 刘晓蔚, 等. 钩栗不同基质和生根剂处理容器育苗对比试验研究[J]. 林业实用技术, 2014(7):41-44.
[19]AbdellahAkhkha. Modelling photosynthetic light-response curve in Calotropisprocera under salinity or water de fi cit stress using non-linear models[J]. Journal of Taibah University for Science,2010, (3): 49-57.
[20]王 佳, 莫惠芝, 蔡静茹, 等. 10中常见垂直绿化植物光响应曲线特性分析[J]. 安徽农业科学, 2015, 43(33): 23-26, 39.
[21]Ningning Wang, QinjunHuangc, Jingru Sun,et al.Shade tolerance plays an important role in biomass production of different poplar genotypes in a high-density plantation[J]. Forest Ecology and Management, 2014, 331:40-49.
[22]李 璟, 叶 充, 蔡仕珍, 等. 遮光对玉竹光合日变化和光合有效辐射-净光合速率响应曲线的影响[J]. 东北林业大学学报, 2015, 43(6):57-63.
[23]吴统贵, 曾广泉, 周晓萍, 等. 红壤区受损湿地松林下6树种光合特性[J]. 江西农业大学学报, 2011, 33(2):318-321.
[24]何冬梅, 刘 庆, 林 波, 等. 人工针叶林林下11种植物叶片解剖特征对不同生境的适应性[J]. 生态学报, 2008,28(10):4739-4749.
[25]Montgomery D C. 实验设计与分析(汪仁官, 陈荣昭译)[M].北京:中国统计出版社, 1998.
[26]傅 杨,杨柳青, 吴红强, 等.高温胁迫下大苞景天的形态特征及生理响应[J]. 中南林业科技大学学报, 2015, 35(4):56-59.
[27]王 雁, 马武昌. 扶芳藤、紫藤等7种藤本植物光能利用特性及耐荫性比较研究[J]. 林业科学研究, 2004, 17(3):305-309.
[28]黄志玲, 姜 英, 郝海坤, 等. 不同光照强度对红锥光影响曲线的影响[J].中南林业科技大学学报, 2014, 34(8):30-33.
[29]金松恒, 李雪芹, 王俊刚. 高温胁迫对无柄小叶榕光合作用的影响[J]. 中国农学通报, 2009, 25(3): 83-87.
[30]王欢欢, 曹福亮, 刘新亮. 高温胁迫下不同叶色银杏嫁接苗光响应曲线的拟合[J]. 南京林业大学学报:自然科学版, 2015,39(2): 14-20.
[31]宋佩东. 5种光响应模型对小叶杨的适用性分析[J]. 辽宁林业科技, 2015(3):9-12.
[32]柴胜丰, 唐健民, 杨 雪, 等. 4种模型对黄枝油杉光合光响应曲线的拟合分析[J]. 广西科学院学报, 2015,31(4):286-291.
[33]邓云鹏, 雷静品, 潘 磊, 等. 不同种源栓皮栎光响应曲线的模型拟合及参数比较[J]. 生态学杂志, 2016, 35(2): 387-394.
[34]施 曼, 程龙霞, 祝遵凌. 欧洲鹅耳枥光响应曲线模型拟合与应用[J]. 福建林学院学报, 2014, 34(4): 349-355.
Effect of temperature on light response curve fi tting of Castanopsis tibetana
ZHANG Bin, LUO Xiao-lang
(College of Forestry, Central South University of Forestry and Technology, Changsha 410004, Hunan, China)
Measuring and fi tting the light response curve is one of the widely used methods when we cultivate the precious trees species.But as the difference among models and the genetic characters of tree species, the application of light response model is very important in the research of precious tree. The research were done in 2 years old seedlings of precious timber tree species ofCastanopsis tibetana,using LI-6400 photosynthesis system to measure the photosynthesic characteristics ofCastanopsis tibetana, compare 5 different light response models for the fi tting of the characteristic data of the combination at 25℃ and 45℃ , the results show: high temperature stress can enhance the ability of using weak light, and also signi fi cantly reduced the maximum net photosynthetic capacity; under normal temperature (25℃) the value of modi fi ed rectangular hyperbolic model (MRHM) is higher by 12.99% - 30.57% compare to other models and measured value, while higher by 12.44% - 26.91% under higher temperature (45℃ ); compare all the models, under 25℃ ,exponential model (EM), and modi fi ed exponential model (MEM) and non-rectangular hyperbolic model (NHM) have the best fi tting effect, but under the condition of 45℃, NHM and EM are the best models; Compere the real value and parameters of photosynthetic curve, NHM is the best model without the in fl uence of temperature.
light response curve; light response model;Castanopsis tibetanaHance; photosynthetic physiology
S718.51+2.2;Q945.11
A
1673-923X(2016)12-0055-06
10.14067/j.cnki.1673-923x.2016.12.010
http: //qks.csuft.edu.cn
2016-03-23
国家林业行业公益性项目(201204405)
张 斌,讲师,硕士;E-mail:zhangbin@csuft.edu.cn
张 斌,罗小浪. 不同温度对钩栗光响应曲线拟合的影响[J].中南林业科技大学学报,2016, 36(12): 55-60.
[本文编校:文凤鸣]
