小盗龙亚科(恐龙:兽脚类)与鸟类起源研究
2016-11-30胡东宇
裘 锐, 胡东宇,2,3
(1. 沈阳师范大学 古生物学院, 沈阳 110034;2. 国土资源部 东北亚古生物演化重点实验室, 沈阳 110034;3. 辽宁省古生物演化与古环境变迁重点实验室, 沈阳 110034)
小盗龙亚科(恐龙:兽脚类)与鸟类起源研究
裘 锐1, 胡东宇1,2,3
(1. 沈阳师范大学 古生物学院, 沈阳 110034;2. 国土资源部 东北亚古生物演化重点实验室, 沈阳 110034;3. 辽宁省古生物演化与古环境变迁重点实验室, 沈阳 110034)
小盗龙亚科兽脚类恐龙是我国辽宁西部早白垩世“热河生物群”带羽毛恐龙的代表,在恐龙系统演化中位于兽脚亚目驰龙科的基干位置。小盗龙亚科恐龙所表现出的一些骨骼和软组织特征,如肩臼窝开口于体侧、较长的前肢、具不对称正羽等,为鸟类起源、鸟类飞行起源以及近鸟类恐龙的演化研究提供了重要信息。回顾了近年来有关小盗龙亚科的一系列重要发现,总结了已发表属种的特征,归纳了该类群对鸟类起源以及鸟类飞行起源研究具有重要意义的骨骼和羽毛特征。也对小盗龙亚科与飞行有关的特征进行功能分析,分析结果进一步支持鸟类飞行的“树栖起源假说”。
热河生物群; 带羽毛恐龙; 骨骼特征; 鸟类飞行起源
0 引 言
小盗龙亚科(Microraptorinae)在恐龙系统演化中是位于兽脚亚目驰龙科(Dromaeosauridae)下的一个类群,该类群和驰龙科的其他类群具有很多同源特征:额骨上颞孔边缘呈“S”形,齿骨背腹边缘近平行,背椎椎体关节横突短棒状,尾椎前关节突和脉弧拉长,乌喙骨具发达的肩臼下凹,第5跖骨较长超过其他跖骨一半,第2趾特化等[1]。由于该类群也具有颈椎上突小,背椎椎体长大于高,且缺乏明显的侧凹,第2跖骨滑车关节较浅,第2趾第2节近端腹突不发达,而且骨体未明显上下收缩等相对原始的特征[2],因此在目前的系统演化分析中常将其置于驰龙科的基干位置(图1)。

图1 小盗龙亚科在近鸟类兽脚类恐龙中的系统发育位置简图
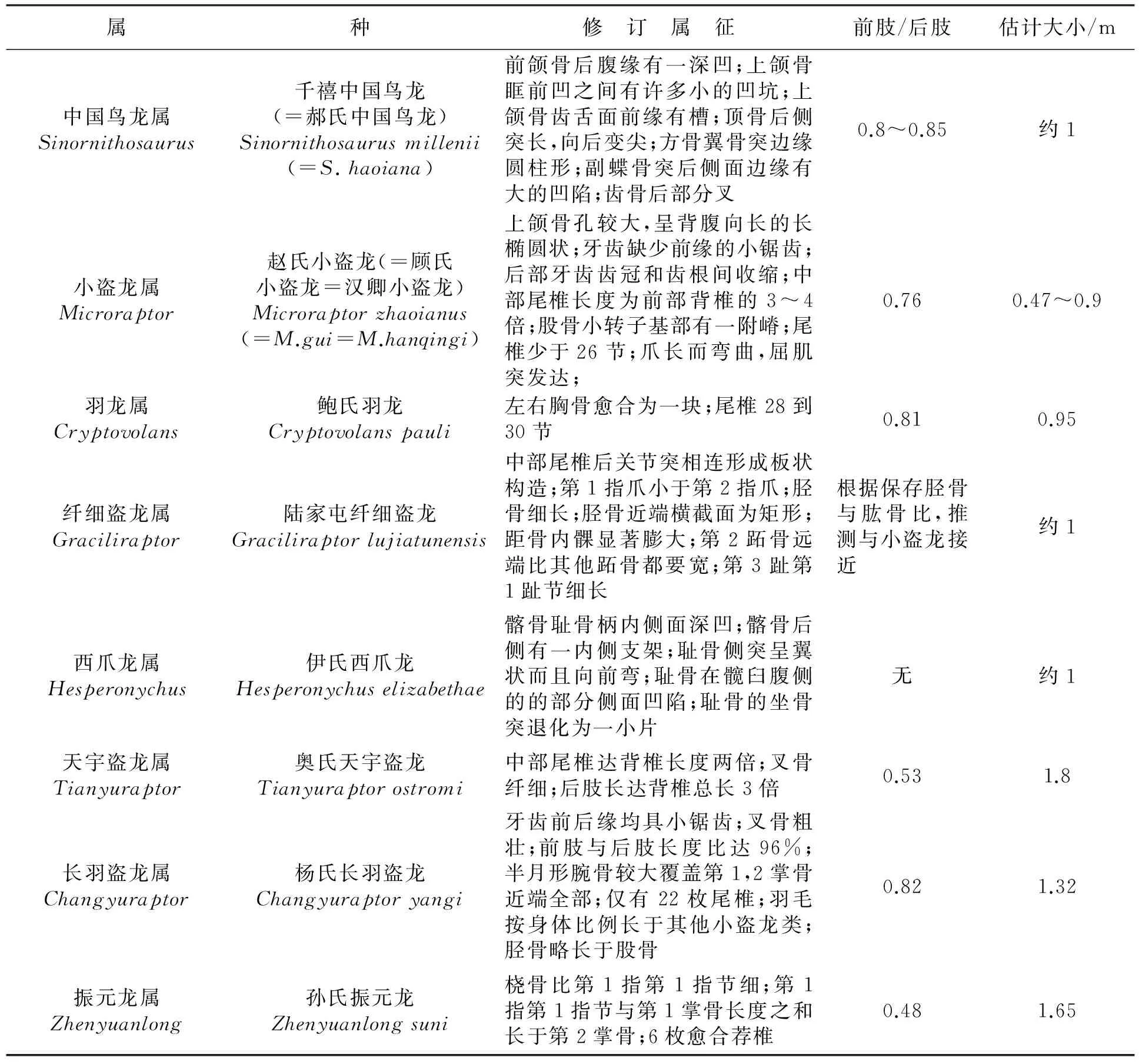
属种修 订 属 征前肢/后肢估计大小/m中国鸟龙属Sinornithosaurus千禧中国鸟龙(=郝氏中国鸟龙)Sinornithosaurusmillenii(=S.haoiana)前颌骨后腹缘有一深凹;上颌骨眶前凹之间有许多小的凹坑;上颌骨齿舌面前缘有槽;顶骨后侧突长,向后变尖;方骨翼骨突边缘圆柱形;副蝶骨突后侧面边缘有大的凹陷;齿骨后部分叉0.8~0.85约1小盗龙属Microraptor赵氏小盗龙(=顾氏小盗龙=汉卿小盗龙)Microraptorzhaoianus(=M.gui=M.hanqingi)上颌骨孔较大,呈背腹向长的长椭圆状;牙齿缺少前缘的小锯齿;后部牙齿齿冠和齿根间收缩;中部尾椎长度为前部背椎的3~4倍;股骨小转子基部有一附嵴;尾椎少于26节;爪长而弯曲,屈肌突发达;0.760.47~0.9羽龙属Cryptovolans鲍氏羽龙Cryptovolanspauli左右胸骨愈合为一块;尾椎28到30节0.810.95纤细盗龙属Graciliraptor陆家屯纤细盗龙Graciliraptorlujiatunensis中部尾椎后关节突相连形成板状构造;第1指爪小于第2指爪;胫骨细长;胫骨近端横截面为矩形;距骨内髁显著膨大;第2跖骨远端比其他跖骨都要宽;第3趾第1趾节细长根据保存胫骨与肱骨比,推测与小盗龙接近约1西爪龙属Hesperonychus伊氏西爪龙Hesperonychuselizabethae髂骨耻骨柄内侧面深凹;髂骨后侧有一内侧支架;耻骨侧突呈翼状而且向前弯;耻骨在髋臼腹侧的的部分侧面凹陷;耻骨的坐骨突退化为一小片无约1天宇盗龙属Tianyuraptor奥氏天宇盗龙Tianyuraptorostromi中部尾椎达背椎长度两倍;叉骨纤细;后肢长达背椎总长3倍0.531.8长羽盗龙属Changyuraptor杨氏长羽盗龙Changyuraptoryangi牙齿前后缘均具小锯齿;叉骨粗壮;前肢与后肢长度比达96%;半月形腕骨较大覆盖第1,2掌骨近端全部;仅有22枚尾椎;羽毛按身体比例长于其他小盗龙类;胫骨略长于股骨0.821.32振元龙属Zhenyuanlong孙氏振元龙Zhenyuanlongsuni桡骨比第1指第1指节细;第1指第1指节与第1掌骨长度之和长于第2掌骨;6枚愈合荐椎0.481.65
小盗龙亚科恐龙具有很多与鸟类相似的骨骼特征[2],并且在很多标本发现有与现生鸟类飞羽相似的羽毛印痕[3-8]。虽然该亚科恐龙并不是鸟类的直接祖先,但由于驰龙科所归属恐爪龙类(Deinonychosauria)与鸟类是具有最近共同祖先的姊妹群,而小盗龙亚科又是驰龙科乃至恐爪龙类中的一个原始类群,所以对小盗龙亚科的研究会为人们认识鸟类起源,以及鸟类飞行起源提供重要信息。
1 小盗龙亚科已知属种
千禧中国鸟龙(Sinornithosaurusmillenii)是小盗龙亚科最早被描述的成员,1999年发现于我国辽宁西部早白垩世“热河生物群”地层中[9]。至今,小盗龙亚科已有8属8种被报导,其中除西爪龙(Hesperonychus)发现于加拿大艾伯塔省晚白垩世恐龙公园组外[10],其他属种全部发现于我国辽西早白垩世义县组和九佛堂组(图2)。2009年Longrich等研究认为,已发现中国鸟龙、纤细盗龙(Graciliraptor)、小盗龙(Microraptor)、西爪龙在驰龙科中组成了一个单系类群,并以此建立小盗龙亚科[10]。后来陆续发现的天宇盗龙(Tianyuraptor)、长羽盗龙(Changyuraptor)、振元龙(Zhenyuanlong)也被归入该亚科[7-8]。
2 小盗龙亚科为“鸟类恐龙起源假说”提供了重要证据
鸟类起源问题长期以来一直存在争议。20世纪中叶以来,随着大量近鸟类恐龙化石陆续被发现,鸟类与兽脚类恐龙中恐爪龙类骨骼特征的相似性越来越被揭示[11- 12],因此“鸟类恐龙起源假说”也被越来越多的学者所接受。小盗龙亚科恐龙的发现则为“鸟类恐龙起源假说”提供了更为有力的证据。

A: 长羽盗龙;B:天宇盗龙; C: 振元龙; D: 小盗龙; E: 中国鸟龙。比例尺均为10 cm。
2.1 小盗龙亚科与鸟类具有相似的骨骼特征
小盗龙亚科恐龙与原始鸟类在骨骼特征上表现出很多的相似性[2,13](表2),这反映出二者间具有很近的亲缘关系,为鸟类与恐爪龙类具有最近的共同祖先的论点提供了化石证据。
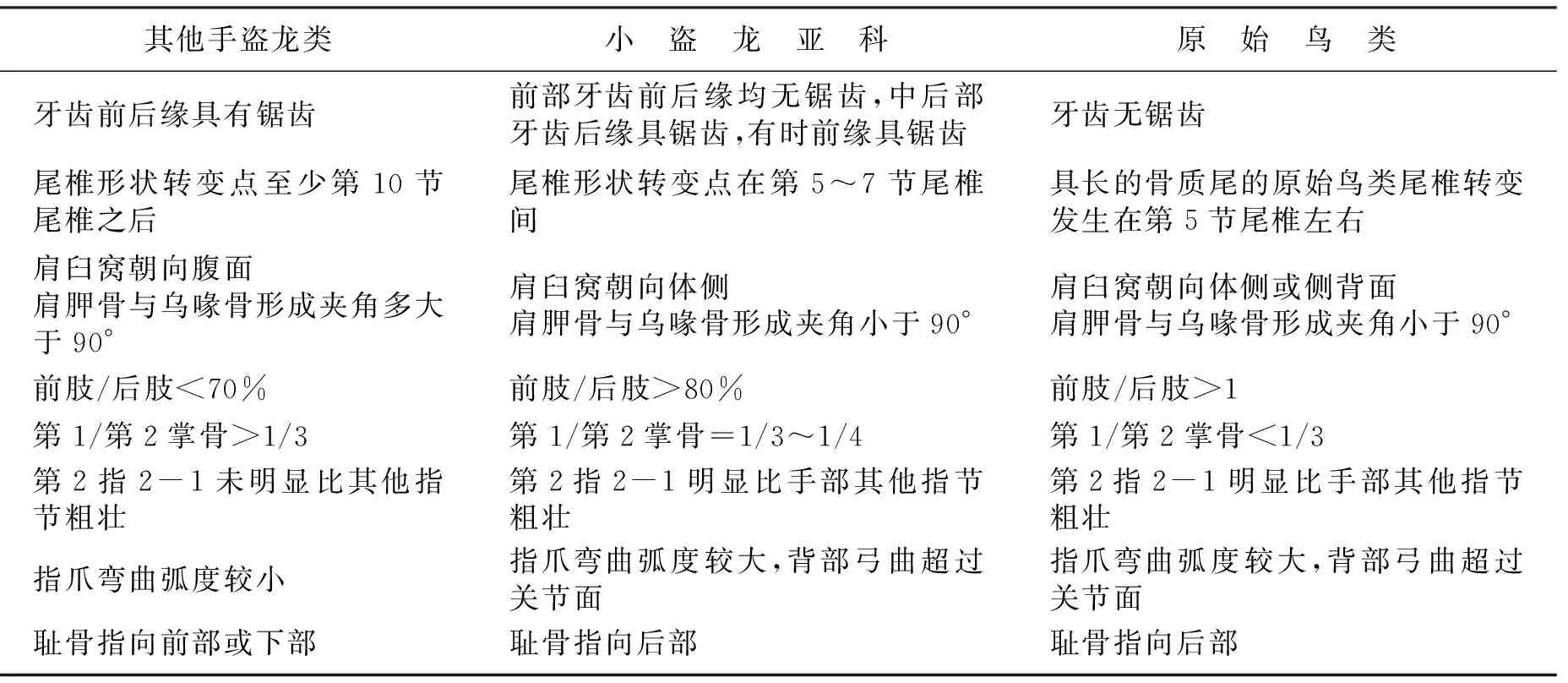
表2 其他手盗龙类、小盗龙亚科、原始鸟类的骨骼特征对比
2.2 小盗龙亚科恐龙与鸟类具有相似的羽毛特征
小盗龙亚科恐龙不仅具有与鸟类相似的骨骼特征,并且体表被有与鸟类相似的羽毛。目前在小盗龙亚科的8个属中,有5个属的标本保存有羽毛印痕[2-8],而且羽毛的形态结构已经有较高程度的分化,既有较为简单的不具分枝的毛状羽毛,也有与现生鸟类飞羽相似的,具有复杂分枝结构的片状羽毛(表3)。其他尚未发现有羽毛印痕保存的属种,很可能是由于某些原因其原来被有的羽毛未能在化石中保存下来所致。小盗龙亚科恐龙的羽毛与鸟类羽毛的相似性不仅体现在宏观形态上,也表现在微观结构上。在扫描电镜下已观察到,在小盗龙的羽毛印痕上存在有与现生鸟类羽毛中相似的亚细胞结构——“色素体”,在现生鸟类中,这些亚细胞结构决定了羽毛的颜色。因此,人们通过与现生鸟类色素体的对比,复原了小盗龙的羽毛颜色[14],也从亚细胞结构上进一步证实了恐龙羽毛与鸟类羽毛的同源性,为鸟类恐龙起源假说提供了更充分的证据。

表3 小盗龙亚科各属的羽毛特征
3 小盗龙亚科为鸟类飞行起源提供了重要证据
关于鸟类飞行起源,主要存在着“地栖起源”“树栖起源”和“斜坡起源”等3种假说。“地栖起源假说”最早由美国古生物学家Williston于1878年提出,之后许多学者给予了充实和丰富。该假说主要依据始祖鸟后肢胫骨与股骨长度比例较大,第1趾未完全反转等特征,认为鸟类的祖先是一类擅长奔跑的动物,飞行是在地面奔跑、跳跃中逐渐获得的[15]。该假说存在的问题是:在自然界中,不管是地史时期的,还是现在的地栖奔跑型两足动物,在后肢变得强壮的同时,其前肢都趋于退化,以减轻体重,减少空气阻力,提高奔跑时的稳定性。然而,鸟类要想飞向天空,必须演化出长的前肢,因此二者之间存在对立[16]。鸟类的祖先若通过在陆地上快速助跑起飞的话,长的翅膀势必带来大的阻力,要想达到足够的速度,又需要有更强有力的后肢,这无疑又增加了体重。“地栖起源假说”一直没能很好地解释上述问题,而且目前也没有具有说服力的化石证据。
“斜坡起源假说”由美国学者Dial等提出,他们观察到尚不具备飞行能力的幼鸟可借助拍打翅膀爬上近乎垂直的斜坡,以此推测鸟类祖先在斜坡上运动时逐渐具有了拍打前肢的能力,最终前肢演化成翅膀获得了飞行能力[15]。该假说仍然强调两足行走为飞行起源的前提条件,实际上是特定环境下的“地栖起源假说”,因此同样存在着“地栖起源假说”没能很好解释的问题。
“树栖起源假说”最初由美国古生物学家Marsh于1880年提出,该假说认为鸟类的祖先在演化过程中逐渐适应了树栖生活,羽毛特别是前肢上的羽毛发育,提高了在树间以及从树上到地面跳跃滑翔的能力,最终在滑翔中学会了拍打翅膀飞行[15]。与地栖起飞必须克服重力影响不同,树栖起飞不是克服而是利用了重力,因此“树栖起源假说”从起飞动力学上得到了很好的解释。小盗龙亚科恐龙的发现首次为该假说提供了可信的化石证据。
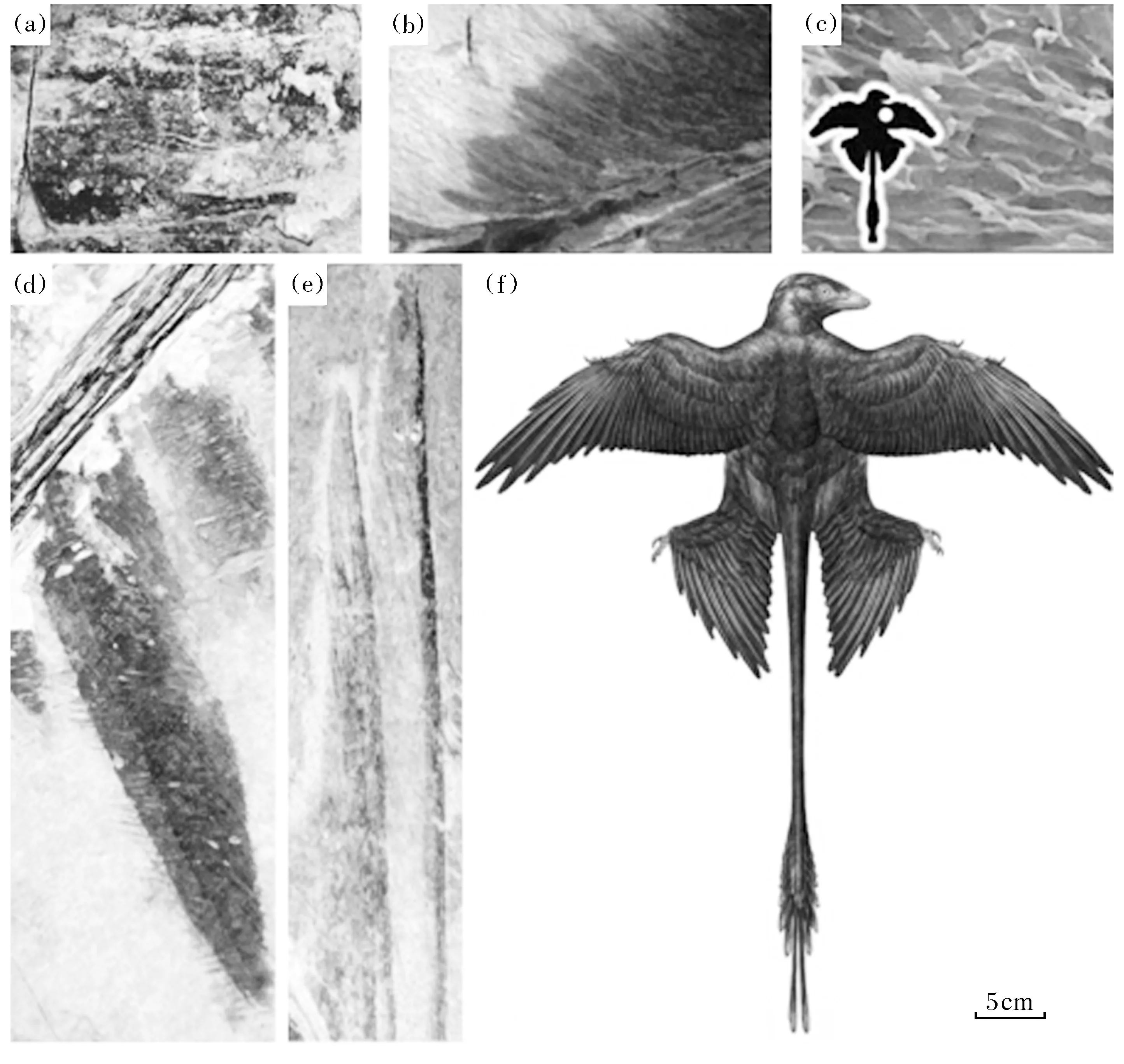
(a)—附着于中国鸟龙头部簇状绒羽(NGMC 91) [4];(b)—附着于羽龙后肢具分支结构正羽,羽毛左右对称(BPM1 3-13)[5];(c)—扫描电镜下观察到的小盗龙肩部羽毛黑色素[14];(d)、(e)—附着于小盗龙后肢具分支结构的正羽,羽毛左右不对称(BMNHC PH881/ IVPP V13352)[6,14];(f)—小盗龙羽毛及颜色复原图[14]。
已知的小盗龙亚科恐龙可简单地划分为长臂和短臂2种类型(表1),大多数属种表现为长臂类型,仅2个属种为短臂类型。长臂类型的体型相对较小,手部在其前肢中占很大的比例[2],手部和足部倒数第2指(趾)节长于倒数第3指(趾)节,远端指(趾)节多为大而弯曲的爪17-18],表现出对树栖攀援抓握生活的适应[19]。小盗龙亚科恐龙普遍具有朝向侧面的肩臼,使前肢可以完成侧向的抬升运动[2]。进一步的观察发现长臂类型的腰带和股骨发生一系列有利于股骨外展的变化:髋臼内部部分闭合,股骨头不能像其他恐龙那样完全插入髋臼,并且上髋臼冠有一定程度的退化,股骨外展的范围扩大[20],有学者估计小盗龙的股骨可以外展大约75°角[21];导致股骨外展的髂股肌起于髂骨背部而附着于股骨转子架[22],在其他恐龙中股骨转子架较为退化,而在该类型个体中却极发达形成侧嵴[23],说明髂股肌仍然很强壮。前后肢侧展范围扩大可使长臂类型小盗龙在攀爬树木时像现生攀爬树木的哺乳动物一样使重心尽可能贴近树干,减小重力力矩。
长臂类群不仅表现出对树栖生活的适应,并且是已知向飞行方面演化程度最高的非鸟类兽脚类恐龙。小盗龙属恐龙已经具有了一个结构上与现生鸟类相似的翼:前肢伸长,尺骨后弓[2,6];前肢羽毛可区分为着生于手部的初级飞羽和着生于尺骨的次级飞羽,最长初级飞羽达肱骨长度2.7倍,次级飞羽亦长于肱骨,并且飞羽已经演化成为具有空气动力学功能的羽轴,两侧羽片不对称[6];第1掌骨与肱骨间有翼膜形成[24],在现生鸟类中它可以防止前肢在迎风飞行时脱位或折断;第1指上着生有羽毛,在现生鸟类中该位置着生的羽毛——“小翼羽”在低速滑翔时有保持平衡作用[6]。具有空气动力学功能的不对称型飞羽不仅出现在前肢上构成“前翼”,也排列在后肢跖骨和胫骨上形成了“后翼”[6-7],因而形象地将该属种恐龙称之为“四翼恐龙”。显然支撑着一个大“后翼”的腿是很难在陆地上奔跑的,这也为其树栖生活习性提供了有力的证据。小盗龙属恐龙具有一个长而僵直的骨质尾,骨质尾的后部也着生有长的尾羽,形成宽阔的表面,甚至可视作“第五翼”[6-7]。
针对小盗龙的形态特征,人们对它可能的飞行方式和飞行能力也进行过分析,包括对其复原模型进行风洞实验。研究表明小盗龙具有很高的滑翔能力,很可能也有简单的扇动翅膀飞行的能力。“四翼”可提供足够的上升力使之能有效地滑翔[25],而后翼和尾部末端扇形尾羽对滑翔速度具有有效的控制作用[7]。新的发现表明“四翼”形态不仅见于小盗龙亚科,也不同程度存在于多种原始近鸟类恐龙与原始鸟类中[26-28],说明“四翼”是一种较普遍的特征,鸟类飞行演化初期很可能存在一个“四翼飞行”阶段。小盗龙亚科长臂类型的发现为鸟类飞行“树栖起源假说”提供了有力的证据。
小盗龙亚科短臂类型的体型相对较大,后肢相对长而粗壮,趾爪相对直而短,反映出对陆地奔跑生活的适应[8,29]。该类型与长臂类型之间的演化关系仍存在着争议,最近发现短臂类型的前肢也着生有长的飞羽,甚至是不对称型羽毛,因此认为短臂类型可能是长臂类型从树栖重新返回地栖生活而演化形成的次生特征[8]。
4 结 论
小盗龙亚科恐龙的发现促进了人类对鸟类起源和鸟类飞行起源的认识,为“鸟类恐龙起源假说”和“飞行树栖起源假说”提供了重要的证据。但对于上述假说的质疑依然存在[30],甚至对于小盗龙亚科恐龙的树栖习性以及2个形态类型的演化关系依然存在着争议和不确定性[29]。由于化石的埋藏环境和动物自身特点,小盗龙亚科化石多为二维保存,骨骼有不同程度的变形和破损,制约了人们对其骨骼特征的观察认识,特别是对骨骼间活动方式的认知还多停留在猜测水平上,难以像其他一些恐龙那样[31]通过实际测量数据来判断肢体骨骼的活动范围、骨骼肌肉附着状况,并模拟动物实际的运动方式以推断其生活习性。另外对小盗龙亚科恐龙的个体发育以及系统发育研究也很薄弱。解决这些问题,还需要通过新的技术手段对大量标本进行更细致的观察研究。对小盗龙亚科恐龙的深入研究将为探究鸟类起源和鸟类飞行起源等理论问题提供更多有价值的信息。
[ 1 ]NORELL M A, MAKOVICKY P J. The Dinosauria[M]. Berkeley: University of California Press, 2004.
[ 2 ]XU X. Deinonychosaurian fossils from the Jehol Group of western Liaoning and the coelurosaurian evolution[D].Beijing: Chinese Academy of Sciences, 2002.
[ 3 ]XU X, ZHOU Z H, PRUM R O. Branched integumental structures in Sinornithosaurus and the origin of feathers[J]. Nature, 2001,410:200-204.
[ 4 ]JI Q, NORELL M A, GAO K Q. The distribution of integumentary structures in a feathered dinosaur[J]. Nature, 2001,410:1084-1088.
[ 5 ]NORELL M A, JI Q, GAO K Q, et al. “Modern” feathers on a non-avian dinosaur[J]. Nature, 2002,416:36-37.
[ 6 ]XU X, ZHOU Z H, WANG X L, et al. Four-winged dinosaurs from China[J]. Nature, 2003,421:335-340.
[ 7 ]HAN G, CHIAPPE L M, JI S A,et al. A new raptorial dinosaur with exceptionally long feathering provides insights into dromaeosaurids flight performance[J]. Nature Communications, 2014,5:4382.
[ 8 ]LÜ J C, BRUSATTE S L. A large, short-armed, winged dromaeosaurid (Dinosauria: Theropoda) from the Early Cretaceous of China and its implications for feather evolution[J]. Scientific Reports, 2015,5:11775.
[ 9 ]XU X, WANG X L, WU X C. A dromaeosaurid dinosaur with a filamentous integument from the Yixian Formation of China[J]. Nature, 1999,401:262-266.
[10]LONGRICH N R, CURRIE P J. A microraptorine (dinosauria-dromaeosauridae) from the Late Cretaceous of North America[J]. PNAS, 2009,106(13):5002-5007.
[11]OSTROM J H. Archaeopteryx and the origin of birds[J]. Biol J Linn Soc, 1976,8:91-182.
[12]GAUTHIER J A. The origin of birds and evolution of flight[M]. San Francisco: California Academy of Sciences, 1986.
[13]SENTER P,KIRKLAND J I, DEBLIEUX D D, et al. New Dromaeosaurids (Dinosauria: Theropoda) from the lower cretaceous of Utah, and the evolution of the Dromaeosaurid tail[J]. PLoS One, 2012,7(5):e36790.
[14]LI Q G, GAO K Q, MENG Q J. Reconstruction of Microraptor and the evolution of iridescent plumage[J]. Science, 2012,335:1215-1219.
[15]李志恒,张玉光,周忠和. 鸟类飞行起源的研究[J]. 自然杂志, 2008,30(1):32-38.
[16]张福成,周忠和. 翱翔天空——动物飞行起源探究[M]. 上海:上海科学技术出版社, 2003.
[17]BURNHAM D A, FEDUCCIA A, MARTIN L D, et al. Tree climbing a fundamental avian adaptation[J]. Biol J Linn Soc, 2011,9(1):103-107.
[18]XU X, ZHOU Z H, WANG X L. The smallest known non-avian theropod dinosaur[J]. Nature, 2000,408:705-708.
[19]张玉光,刘迪,李志恒,等. 基于形态对比和统计分析的中生代鸟类栖息行为的识别和判断[J]. 地质论评, 2010,56(6):875-884.
[20]ALEXANDER D E, GONG E P, MARTIN L D, et al. Model tests of gliding with different hindwing configurations in the four-winged dromaeosaurid Microraptor gui[J]. PNAS, 2010,107(7):2972-2976.
[21]GONG E P, MARTIN L D, BURNHAM D A, et al. A new species of Microraptor from the Jehol Biota of northeastern China[J]. Palaeoworld, 2012,21(2):81-91.
[22]HUTCHINSON J R. The evolution of femoral osteology and soft tissues on the line to extant birds (Neornithes)[J]. Zoo J Linn Soc, 2001,131:169-197.
[23]HWANG S H, NORELL M A, JI Q, et al. New specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from northeastern China[J]. Am Mus Nat, 2002,3381:1-44.
[24]AGNOLIN F L, NOVAS F. Avian Ancestors: A Review of the Phylogenetic Relationships of the Theropods Unenlagiidae, Microraptoria, Anchiornis and Scansoriopterygidae[M]. Berlin: Springer-Verlag, 2013.
[25]DYKE G, KAT R, PALMER C, et al. Aerodynamic performance of the feathered dinosaur Microraptor and the evolution of feathered flight[J]. Nature Communications, 2013,4:2489.
[26]ZHANG F C, ZHOU Z H. Leg feathers in an Early Cretaceous bird[J]. Nature, 2005,431:925.
[27]Hu D Y, Hou L H, Zhang L J, et al. A pre-Archaeopteryx troodontid theropod from China with long feathers on the metatarsus[J]. Nature, 2009,461:640-643.
[28]ZHENG X T, ZHOU Z H, WANG X L, et al. Hind wings in Basal Birds and the evolution of leg feathers[J]. Science, 2013,339:1309-1312.
[29]ZHENG X T, XU X, YOU H L, et al. A short-armed dromaeosaurid from the Jehol Group of China with implications for early dromaeosaurid evolution[J]. Proc Roy Soc B, 2009,277:211-217.
[30]季燕南,王旭日,黑须球子,等. 论鸟类飞行起源的“树栖”与“地栖”假说之争[J]. 地质学刊, 2015,39(2):201-206.
[31]SENTER P. Comparison of forelimb function betweenDeinonychus and Bambiraptor (Theropoda: Dromaeosauridae)[J]. J Vert Paleont, 2006,26(4):897-906.
Received date: 2015-11-27.
Supported: Project supported by the Ministry of Education Key Laboratory under Grant (LABKF1406), and Education Department of Liaoning Province (L2015377, L2014516).
Biography: SONG Shiwei(1983-), male, was born in Dalian city of Liaoning province, lecturer of Shenyang Institute of Engineering, doctor.
文章编号: 1673-5862(2016)02-0140-04
材料科学
Abstract: The Ga-face GaN-based device exist a great polarization field, which cause carriers overflow and other problems to degrade the performance of GaN-based devices. However, N-face GaN can resolve these problems with the inversion of the polarization field. The N-face GaN epilayer was prepared on the C-face SiC substrate by MOCVD system, and the basic characters of the N-face GaN were investigated. N-face GaN has a rough surface, and exists a lot of edge dislocations and mixed dislocations. The yellow band is not observed at room temperature PL spectra despite a lot of Si-doped. After a phosphoric acid solution etching, a large number of Ga vacancies are produced on the surface of epilayer, and the corresponding yellow band appeared, which indicated the yellow band origins from Ga vacancy. After etching, GaN surface covered 12 pyramidal structures, which relaxes the tensile stress. In addition, as corrosion progresses, FWHM of low temperature PL spectra narrowed. Key words: N-face GaN; photoluminescence; gallium vacancy
Key words: N-face GaN; photoluminescence; gallium vacancy
doi: 10.3969/ j.issn.1673-5862.2016.02.003
The subfamily Microraptorinae (Dinosauria: Theropoda) and research in origin of birds
QIURui1,HUDongyu1,2,3
(1. College of Paleontology, Shenyang Normal University, Shenyang 110034, China; 2. Key Laboratory for Evolution of Past Life in Northeast Asia, Ministry of Land and Resources, Shenyang 110034, China; 3. Key Laboratory of Evolution of Past Life & Paleoenvironmental Changes in Liaoning, Shenyang 110034, China)
The subfamily Microraptorinae is a representative group of the feathered dinosaurs from the Early Cretceous “Jehol Biota” of western Liaoning, China. As a basal dromaeosaurid, the osteologic and soft tissue characters of this group, such as laterally faced glenoid fossa, elongated forelimb and asymmetrical pennaceous feathers provide important evidence for the research of the origins of birds and bird flight, and the research of the evolution of deinonychusauria. This article reviews the recent discoveries of Microraptoria, concludes the diagnosis of the reported groups, and summarizes their important osteologic and feather structures similar to birds. An analysis of flight-related functional morphology supports “arboreal hypothesis” of the origin of bird flight.
Jehol Biota; feathered dinosaurs; osteologic characters; the origin of bird flight
N-face GaN epilayer grown on C-face SiC substrate by MOCVD
SONGShiwei1,ZHANGDong1,2,ZHAOYan1,WANGCunxu1,KEYunjie1,LIYucai1,WANGJian1,WANGGang1,DINGYanbo1,WANGHan1,LIULiying1,GUORui1
(1. New Energy Source Institute, Shenyang Institute of Engineering, Shenyang 110136, China;2. School of Physics and Optoelectronic Technology, Dalian University of Technology, Dalian 116024, China)
TN 304.55Document code: A
2015-11-27。
国家自然科学基金资助项目(41172026); 辽宁省科技厅自然科学基金资助项目(201102199); 辽宁省高校优秀科技人才支持计划(LR2012038)。
裘 锐(1989-),男,天津人,沈阳师范大学硕士研究生; 通信作者: 胡东宇(1963-),男,辽宁沈阳人,沈阳师范大学教授,博士。
1673-5862(2016)02-0133-07
Q915.864
A
10.3969/ j.issn.1673-5862.2016.02.002
